



Introduction
The Late Cretaceous is an important interval of time in the development of the North American fish fauna, with many of the families that are characteristic of the modern fauna already present. However, the Late Cretaceous is also poorly understood, with many taxa represented by fragmentary skeletal remains (e.g., Brinkman et al., Reference Brinkman, Newbrey and Neuman2014, Reference Brinkman, Divay, DeMar and Wilson Mantilla2021), and relatively few groups of fishes are known from whole-body fossils from the Late Cretaceous (e.g., Grande, Reference Grande1986; Murray et al., Reference Murray, Newbrey, Neuman and Brinkman2016, Reference Murray, Brinkman, Newbrey and Neuman2020a). Recently, a mass mortality of fishes has been discovered at the Tanis Site in the North Dakota portion of the Hell Creek Formation (DePalma et al., Reference DePalma, Smit, Burnham, Kuiper and Manning2019). This mass mortality, containing many well-preserved, three-dimensional skeletons of fishes, has been interpreted to be the result of a seiche wave (DePalma et al., Reference DePalma, Smit, Burnham, Kuiper and Manning2019) that was produced by the impact of the Chicxulub asteroid off the Yucatan coast, marking the Cretaceous–Paleocene transition and leading to one of Earth's great mass extinctions (Jablonski, Reference Jablonski1994). So far, the fishes described from the Tanis site (Fig. 1) are sturgeons (Acipenseridae) and paddlefishes (Polyodontidae) (Hilton and Grande, Reference Hilton and Grande2022; this paper). The detailed preparation of the polyodontid specimens described herein, and the sturgeons from this locality that were previously described, required several years of effort by fossil preparators. During this time, new information on the structure and biology of the acipenseriform fishes from the Tanis Site, including the undescribed polyodontids (During et al., Reference During, Smit, Voeten, Berruyer, Tafforeau, Sanchez, Stein, Verdegaal-Warmerdam and van der Lubbe2022), was also discovered using propagation phase-contrast synchrotron radiation micro-computed tomography (PPC-SRμCT) and digital reconstruction. By bringing together the data and interpretation of classically prepared specimens and digital reconstructions, we are now able to describe the polyodontids in more detail than by either source alone. This paper is also intended to serve as a complement to that which describes two species of Acipenseridae that were preserved in the same blocks as these polyodontids (Hilton and Grande, Reference Hilton and Grande2022).

Figure 1. Tanis Site, Late Cretaceous (Maastrichtian) Hell Creek Formation, North Dakota, USA. (1) Stratigraphic column with details for the Tanis site, showing the seiche deposit transition between the Hell Creek Formation and the overlying Fort Union Formation (Paleocene). (2) Map showing the geographic location (inset) and extent of the Tanis Site. (3) Photograph showing the seiche deposit of the Hell Creek Formation.
Acipenseriformes (sensu Bemis et al., Reference Bemis, Findeis and Grande1997) includes the extinct families †Chondrosteidae (see Hilton and Forey, Reference Hilton and Forey2009) and †Peipiaosteidae (see Grande and Bemis, Reference Grande, Bemis, Stiassny, Parenti and Johnson1996; Jin, Reference Jin, Chen and Jin1999; Hilton et al., Reference Hilton, Grande and Fan2021) and two extant families: Acipenseridae (sturgeons) and Polyodontidae (paddlefishes). Acipenseridae has approximately 25 extant species in four broadly recognized genera: Acipenser Linnaeus, Reference Linnaeus1758, Huso Linnaeus, Reference Linnaeus1758, Scaphirhynchus Heckel, Reference Heckel1836, and Pseudoscaphirhynchus Nikolskii, Reference Nikolskii1900. It also includes several fossil species of Acipenser (see Hilton and Grande, Reference Hilton and Grande2006, Reference Hilton, Grande and Schultze2021, Reference Hilton and Grande2022; Grande and Hilton, Reference Grande and Hilton2006) plus several monotypic genera known exclusively as fossils (†Protoscaphirhynchus Wilimovsky, Reference Wilimovsky1956; †Priscosturion Grande and Hilton, Reference Grande and Hilton2009; †Anchiacipenser Sato et al., Reference Sato, Murray, Vernygora and Currie2018; †Engdahlichthys Murray et al., Reference Murray, Brinkman, Demar and Wilson2020b [Murray et al., Reference Murray, Brinkman, Demar and Wilson2020b]).
Within Acipenseriformes, Polyodontidae are regarded as a highly derived group of fishes known only from China and North America. The family first appears in the fossil record 130 million years ago in the Early Cretaceous of China (†Protopsephurus liui Lu, Reference Lu1994; see Grande et al., Reference Grande, Jin, Yabumoto and Bemis2002). The modern species Psephurus gladius (Martens, Reference Martens1862), found in China, was declared extinct in 2020 (Zhang et al., Reference Zhang, Jarić, Roberts, He, Du, Wu, Wang and Wei2020). In North America, paddlefishes are known by six species. These include the two Late Cretaceous species described in this paper, the Late Cretaceous †Paleopsephurus wilsoni MacAlpin, Reference MacAlpin1941, the early Paleocene †Polyodon tuberculata Grande and Bemis, Reference Grande and Bemis1991, the early Eocene †Crossopholis magnicaudatus Cope, Reference Cope1883, and the extant Polyodon spathula (Walbaum, Reference Walbaum1792).
The common name “paddlefish” derives from the fact that these fishes have a highly elongated snout or “paddle.” This structure develops through pronounced allometric growth of the rostrum, eventually producing an adult head length that is up to half of the fish's total length (Grande and Bemis, Reference Grande and Bemis1991). The paddle is supported by six series of elongate dorsal and ventral median rostral bones flanked laterally by many loosely interdigitating stellate bones (Grande and Bemis, Reference Grande and Bemis1991). In living polyodontids, the spaces between the stellate bones contain electrosensory ampullary organs that allow the fish to navigate in turbid riverine waters and detect prey items (Northcutt, Reference Northcutt, Bullock and Heiligenberg1986; Bemis and Grande, Reference Bemis and Grande1992). Psephurus was piscivorous, as were at least some fossil taxa such as †Crossopholis (e.g., Grande and Bemis, Reference Grande and Bemis1991, fig. 47) and probably the two species described in this paper given their short gill rakers. By contrast, Polyodon spp. are filter feeders and have elongate gill rakers (unique among polyodontids) that serve to sieve zooplankton from the water column. With only two Recent species and at least six fossil species, much of what is known about the evolution of Polyodontidae lies in the morphological diversity found in the fossil taxa. The purpose of this paper is to describe two new polyodontid taxa from the Tanis Site of the Hell Creek Formation, setting the stage for a review of the systematics and evolution of the family Polyodontidae.
Materials and methods
Preparation, photography, and illustration
Most specimens described herein were mechanically prepared under stereo microscopes with the use of sharpened pin vises. They were then photographed using a Canon EOS 5D digital camera. Some specimens were coated with ammonium chloride sublimate before photography to enhance contrast. Line drawings were rendered by hand by E.J.H. on the basis of these images.
Scanning and digital reconstruction
Specimen FAU.DGS.ND.161.4559.T was scanned at beamline BM05 of the European Synchrotron Radiation Facility (ESRF) in Grenoble, France (Tafforeau et al., Reference Tafforeau, Boistel, Boller, Bravin and Brunet2006; During et al., Reference During, Smit, Voeten, Berruyer, Tafforeau, Sanchez, Stein, Verdegaal-Warmerdam and van der Lubbe2022). The complete specimen, consisting of a skull (without the rostrum) and pectoral girdle, was scanned at a voxel size of 43.5 μm. The white beam of BM05, with an average energy of 132 keV, was used, filtered with Mo and Cu, at 0.4 mm and 9 mm, respectively. The detector consisted of a 2 mm thick LuAG:Ce scintillator optically coupled to a PCO edge 4.2 CLHS sCMOS camera. To obtain sufficient phase contrast, propagation distance was set at 5 m. A total of 205 scans were performed at a vertical field of view of 2.8 mm by taking 5,000 projections at 7 ms intervals, resulting in a double scan of the complete specimen. An increase in lateral field of view was achieved by scanning in half-acquisition mode. The volume was reconstructed using a single-distance phase retrieval algorithm combined with filtered-back projection as implemented in the ESRF software PyHST2. Vertical concatenation, 16-bit conversion, and ring artifact corrections were performed using MATLAB scripts developed at the ESRF. Subsequently, the gill region was scanned at a voxel size of 13.67 μm (filters: 0.4 mm of Mo and 6 mm of Cu; scintillator: LuAG:Ce, 500 μm thick; detected energy: 166 keV; propagation distance: 2.5 m). The gill region was also scanned in half-acquisition mode, consisting of 4,998 projections at 5 ms intervals. After reconstruction, the scans were laterally concatenated. Digital three-dimensional extraction and rendering was performed in VGStudio MAX 3.2 and 3.5 (Volume Graphics). Digital reconstructions were made by MADD. Synchrotron scanning data are available at https://doi.org/10.5281/zenodo.5776294 and the http://paleo.esrf.eu database.
Counts, measurements, and style
Counts and measurements follow Grande and Bemis (Reference Grande and Bemis1991). In cases in which a measurement could not be made due to incomplete preservation, the measurement was recorded as estimated (est.). Daggers (†) precede the names of taxa known exclusively as fossils. Terminology for the skull roofing bones and the bones of the rostrum follow Grande and Bemis (Reference Grande and Bemis1991).
Repositories and institutional abbreviations
Materials that serve as the basis for description of the new species are referred to in the relevant sections that follow. Comparative material is listed in Grande and Bemis (Reference Grande and Bemis1991, Reference Grande, Bemis, Stiassny, Parenti and Johnson1996), Grande et al. (Reference Grande, Jin, Yabumoto and Bemis2002), Hilton and Forey (Reference Hilton and Forey2009), Hilton et al. (Reference Hilton, Grande and Bemis2011, Reference Hilton, Grande and Fan2021), and Hilton and Grande (Reference Hilton and Grande2022).
DMNH EVP, Denver Museum of Nature and Science, Denver, Colorado, USA. FMNH, Field Museum, Chicago, Illinois, USA. FAU.DGS.ND, Florida Atlantic University, Boca Raton, Florida, USA.
Systematic paleontology
Class Osteichthyes Huxley, Reference Huxley1880
Subclass Actinopterygii Cope, Reference Cope1887
Infraclass Chondrostei Müller, Reference Müller1844
Order Acipenseriformes Berg, Reference Berg1940
Family Polyodontidae Bonaparte, Reference Bonaparte1835
Genus †Parapsephurus new genus
Type species
†Parapsephurus willybemisi n. gen. n. sp.
Diagnosis
As for species; monotypic.
Etymology
A combination of Para, from the Greek word pará, meaning “around” or “near,” and psephurus, a recently extinct genus of Polyodontidae with similarly short gill rakers.
Remarks
Among Polyodontidae, the presence of short gill rakers, presumably indicative of a piscivorous diet, is plesiomorphic and found in all polyodontids except Polyodon, as well as other members of the order Acipenseriformes.
†Parapsephurus willybemisi new species
Figures 2–15

Figure 2. †Parapsephurus willybemisi n. gen. n. sp., whole body specimens. (1) Photograph of FMNH PF17545 (holotype); image reversed so anterior facing left. (2) Line drawing of FMNH PF17545 (holotype). (3) Photograph of FMNH PF17626; anterior facing left. (4) Line drawing of FMNH PF17626. Scale bars = 5 cm.

Figure 3. †Parapsephurus willybemisi n. gen. n. sp., skull in lateral view, FMNH PF17545 (holotype). (1) Photograph. (2) Line drawing. Right side, image reversed so that anterior facing left. Scale bars = 5 cm. cb = ceratobranchial; cl = cleithrum; d = dentary; dpl = dermopalatine; dpt = dermopterotic (= intertemporo-supratemporal of Grande and Bemis, Reference Grande and Bemis1991; see Grande et al., Reference Grande, Jin, Yabumoto and Bemis2002); exc = extrascapular bones; fr = frontal; gr = gill rakers; h = hyomandibula; ih = interhyal; mfl = middle fenestra longitudinalis; mpec = ectopterygoid process of the dermopalatine; pa = parietal; pas = parasphenoid; pcl = postcleithrum; pfl = posterior fenestra longitudinalis; pfr = pectoral-fin rays; ppt = palatopterygoid; pt = posttemporal; qj = quadratojugal; scl = supracleithrum.
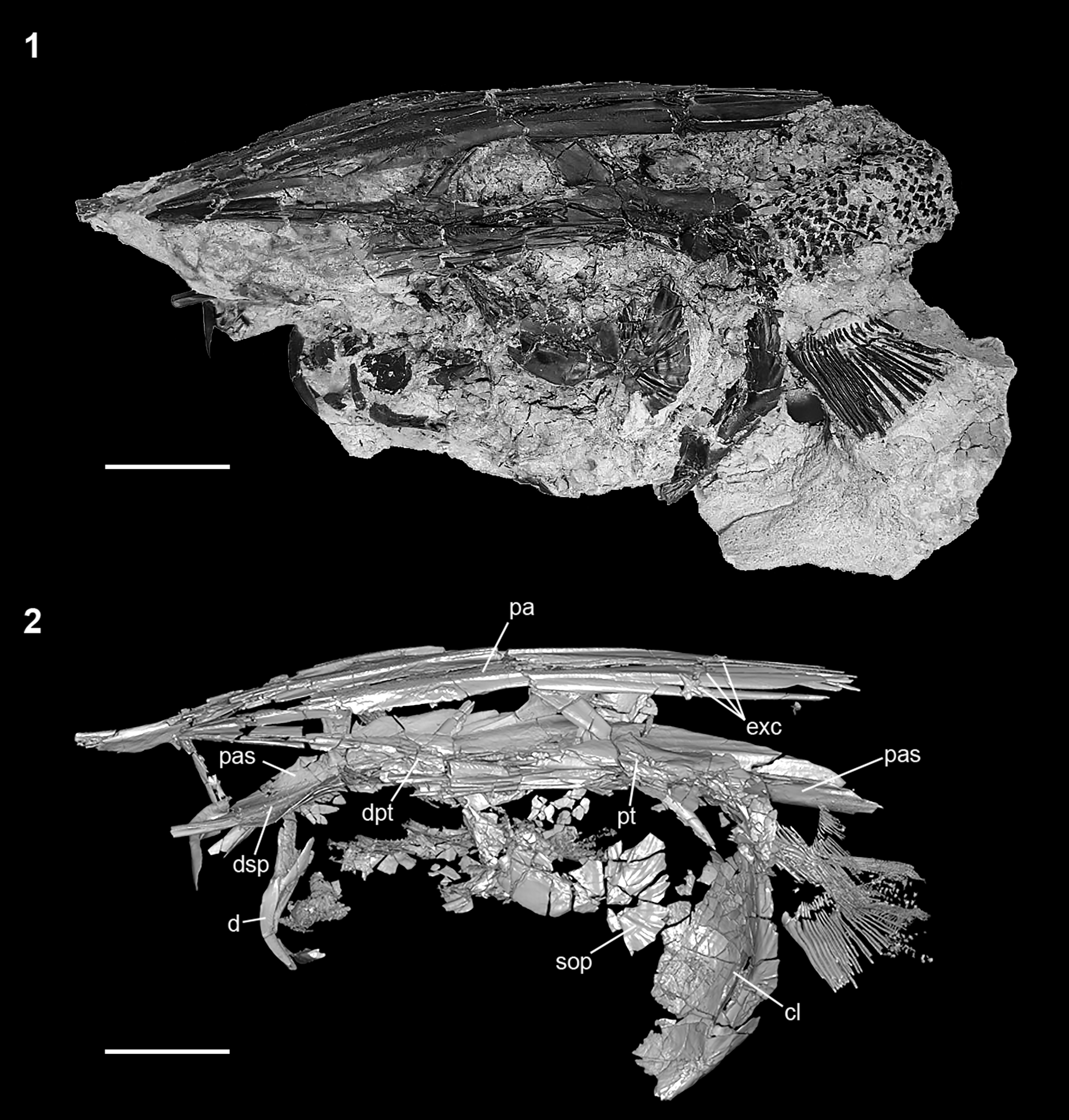
Figure 4. †Parapsephurus willybemisi n. gen. n. sp., Three-dimensional rendering of specimen FAU.DGS.ND.161.4559.T skull in lateral view. Right side, image reversed so that anterior facing left. (1) Photograph of specimen. (2) Digital reconstruction from tomography data. Scale bars = 2 cm. cl = cleithrum; d = dentary; dpt = dermopterotic (= intertemporo-supratemporal of Grande and Bemis, Reference Grande and Bemis1991; see Grande et al., Reference Grande, Jin, Yabumoto and Bemis2002); dsp = dermosphenotic; exc = extrascapular bones; pa = parietal; pas = parasphenoid; pt = posttemporal; sop = subopercle.
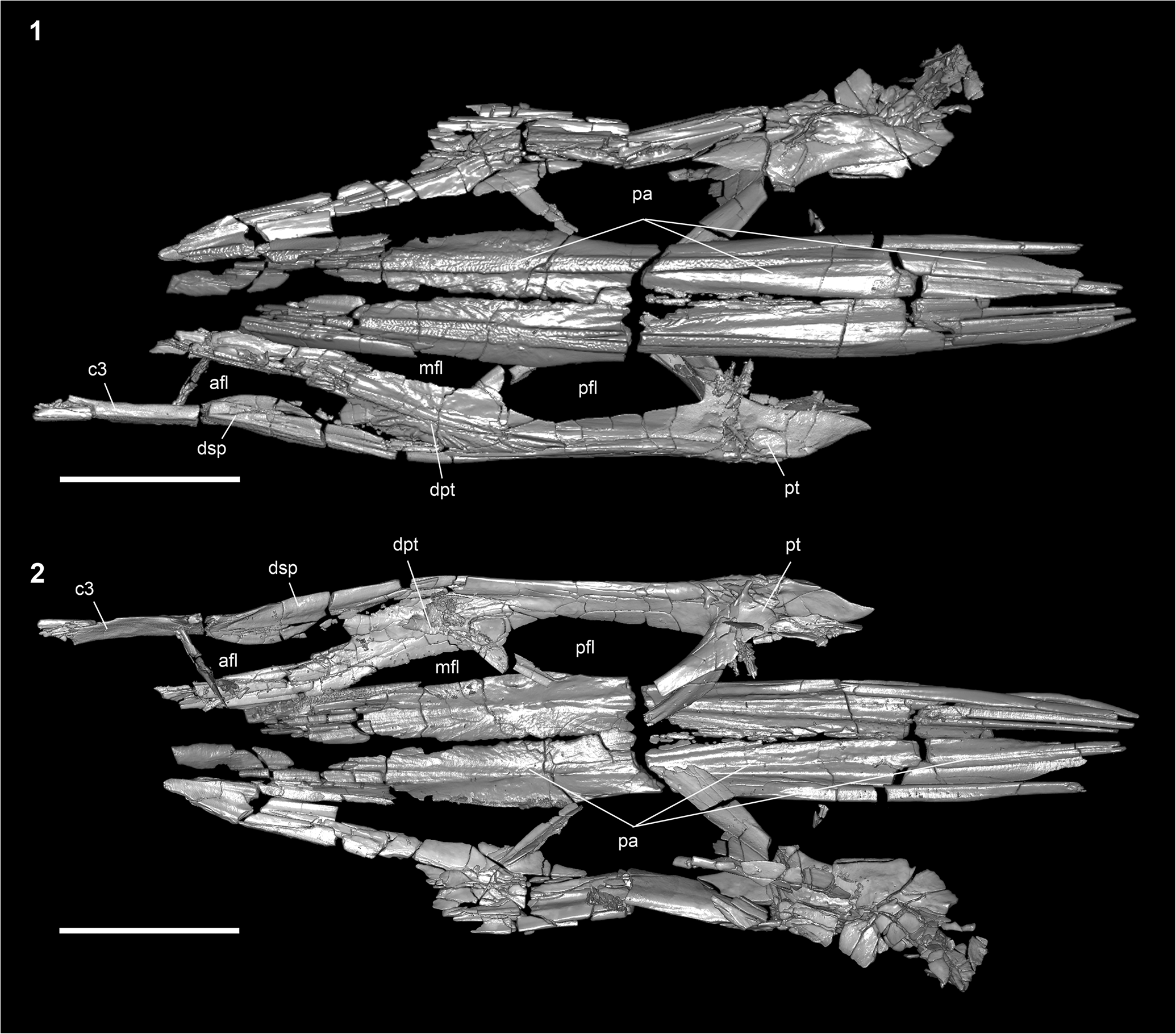
Figure 5. †Parapsephurus willybemisi n. gen. n. sp., digital reconstruction from tomography data of the skull roof; FAU.DGS.ND.161.4559.T. (1) Dorsal view. (2) Ventral view. Anterior facing left. Scale bar = 2 cm. afl = anterior fenestra longitudinalis; c3 = posterior bone of the c series; dpt = dermopterotic (= intertemporo-supratemporal of Grande and Bemis, Reference Grande and Bemis1991; see Grande et al., Reference Grande, Jin, Yabumoto and Bemis2002); dsp = dermosphenotic; mfl = middle fenestra longitudinalis; pa = parietal; pfl = posterior fenestra longitudinalis; pt = posttemporal.
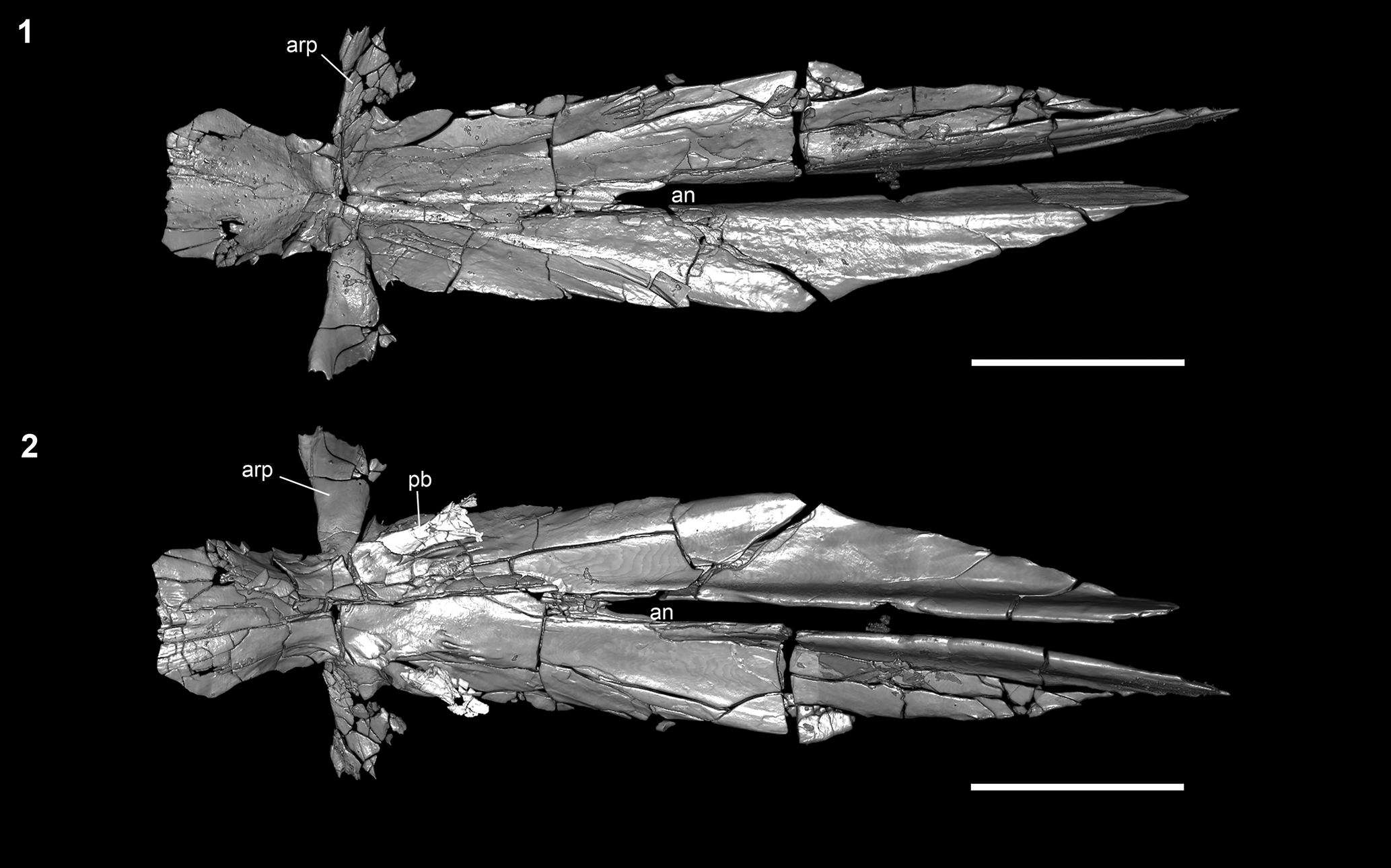
Figure 6. †Parapsephurus willybemisi n. gen. n. sp., digital reconstruction from tomography data of parasphenoid; FAU.DGS.ND.161.4559.T. (1) Dorsal view. (2) Ventral view. Anterior facing left. Scale bar = 2 cm. an = aortic notch; arp = ascending ramus of the parasphenoid; pb = pharyngobranchial.

Figure 7. †Parapsephurus willybemisi n. gen. n. sp., skull in ventral view, FMNH PF17626. (1) Photograph. (2) Line drawing. Scale bars = 2 cm. br = branchiostegal; c3 = posterior bone of the c series; cb = certatobranchial; cha = anterior ceratohyal; chp = posterior ceratohyal; cl = cleithrum; clv = clavicle; d = dentary; dpl = dermopalatine; hh = hypohyal; ih = interhyal; mpec = ectopterygoid process of the dermopalatine; p = median ventral rostral bone of the p series; pas = parasphenoid; pfr = pectoral-fin rays; urb = unidentified rostral bone; v = vomer; (l) = left; (r) = right.
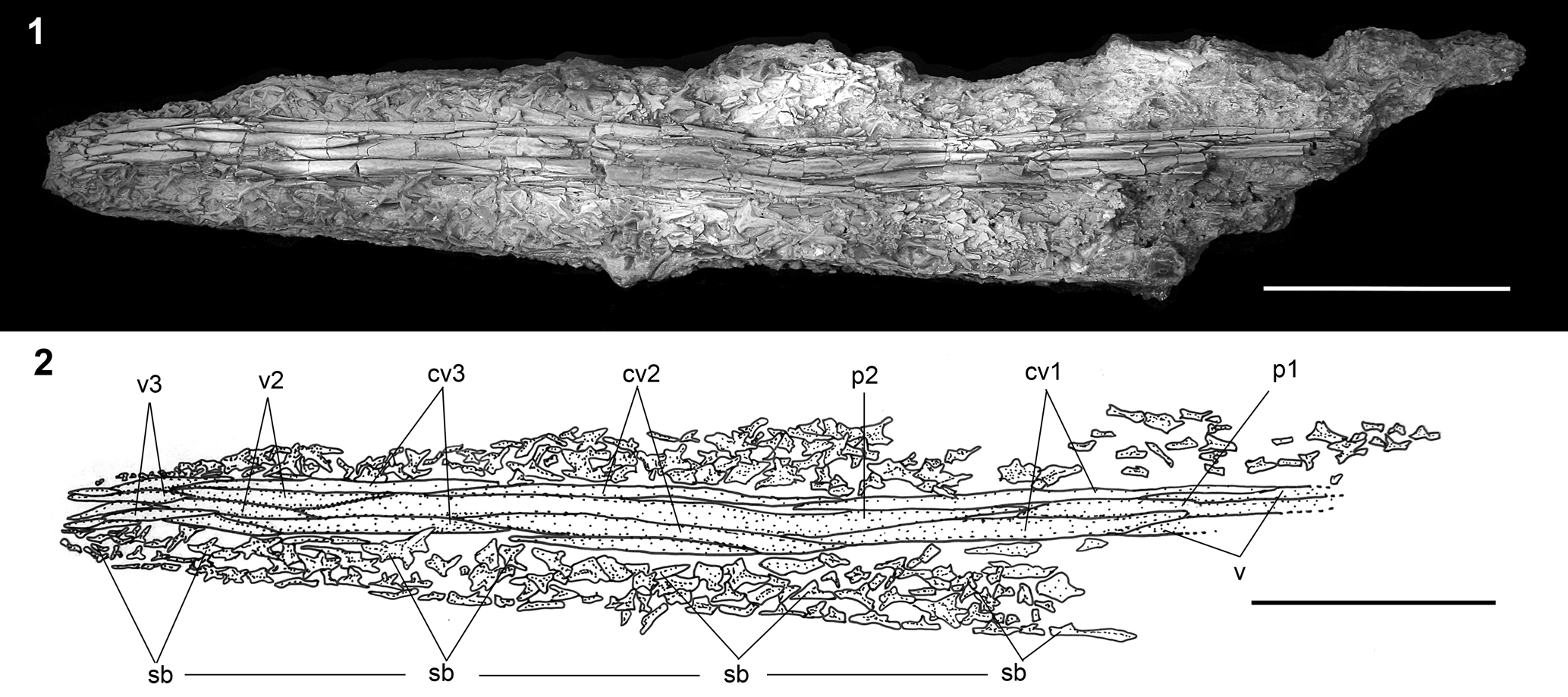
Figure 8. †Parapsephurus willybemisi n. gen. n. sp., rostrum in ventral view, FMNH PF17545 (holotype). (1) Photograph. (2) Line drawing. Anterior facing left. Scale bars = 5 cm. cv1–cv3 = paired outer lateral ventrolateral bones of the rostrum; p1, p2 = unpaired ventral rostral splints belonging to the median series; sb = stellate bones; v = vomer; v2, v3 = paired ventral rostral splints.
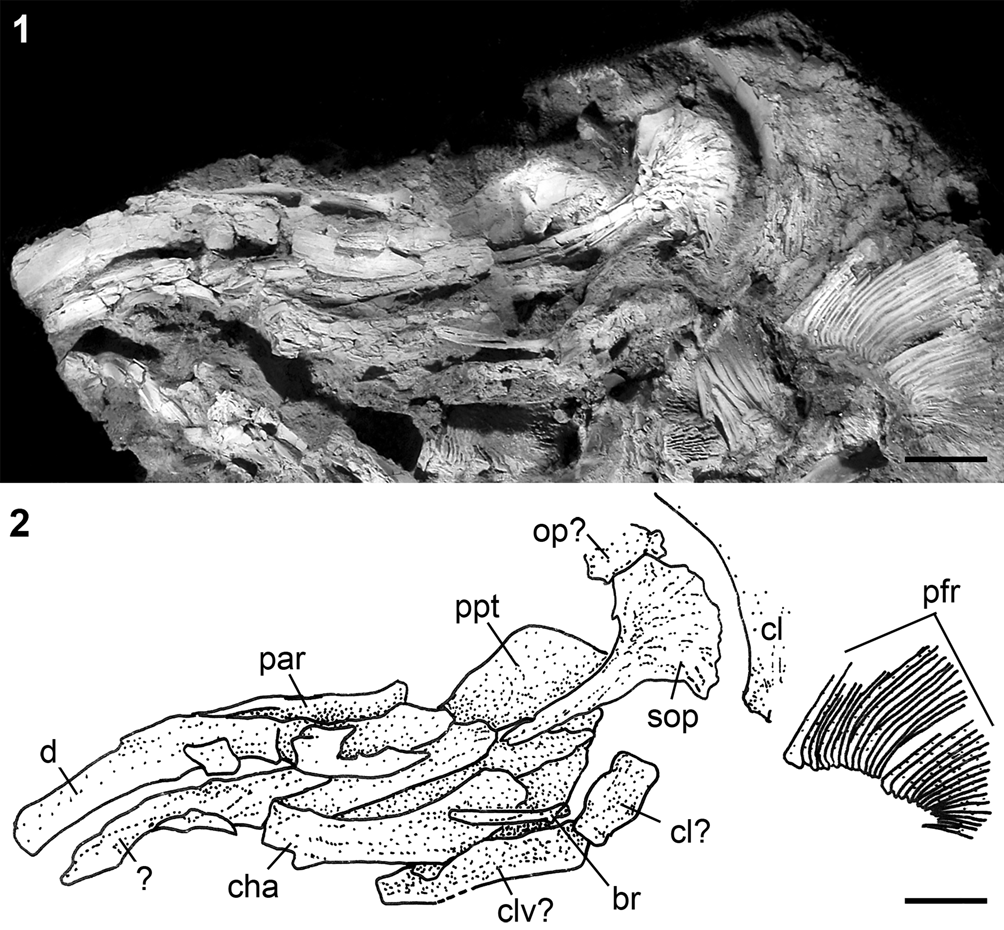
Figure 9. †Parapsephurus willybemisi n. gen. n. sp., bones of the jaws, opercular series, and other skull elements, FMNH PF17627. (1) Photograph. (2) Line drawing. Anterior facing left. Scale bars = 2 cm. br = branchiostegal; cha = anterior ceratohyal; cl = cleithrum; clv = clavicle; d = dentary; op = opercle; par = prearticular; pfr = pectoral-fin rays; ppt = palatopterygoid; sop = subopercle.

Figure 10. †Parapsephurus willybemisi n. gen. n. sp., jaws in lateral view, FMNH PF17545 (holotype). (1) Photograph. (2) Line drawing. Right side, image reversed so that anterior facing left. Scale bars = 2 cm. d = dentary; dpl = dermopalatine; mpec = ectopterygoid process of the dermopalatine; ppt = palatopterygoid; qj = quadratojugal.

Figure 11. †Parapsephurus willybemisi n. gen. n. sp., digital reconstruction from tomography data of isolated jaw bones of FAU.DGS.ND.161.4559.T. (1–4) Dermopalatine: (1) dorsal view; (2) lateral view; (3) ventral view; (4) medial view. (5–8) Lower jaw: (5) dorsal view; (6) lateral view; (7) ventral view; (8) medial view. Anterior facing left in all except (4, 8). Scale bars = 5 mm. d = dentary; dpl = dermopalatine; m = mentomeckelian; mpec = ectopterygoid process of the dermopalatine; par = prearticular; (l) = left; (r) = right.

Figure 12. †Parapsephurus willybemisi n. gen. n. sp. (1, 2) Digital reconstruction from tomography data of isolated ventral portions of the gill arches of specimen FAU.DGS.ND.161.4559.T: (1) dorsal view; (2) ventral view. (3) Exposed gill arch with gill rakers (arrows) in lateral view, FMNH PF17545 (holotype). Anterior facing left in all (in 3, image reversed). (1, 2) Scale bars = 5 mm; (3) scale bar = 2 cm. cb = ceratobranchial (arches unidentified); gr = gill rakers; hb = hypobranchial.

Figure 13. †Parapsephurus willybemisi n. gen. n. sp., median fins in lateral view, FMNH PF17545 (holotype). (1) Photograph. (2) Line drawing. Right side, image reversed so that anterior is facing left. Scale bars = 5 cm. afr = anal-fin rays; drf = dorsal-fin rays; mr = middle radials; pr = proximal radials.

Figure 14. †Parapsephurus willybemisi n. gen. n. sp., caudal fin in lateral view, FMNH PF17545 (holotype). (1) Photograph. (2) Line drawing. Right side, image reversed so that anterior is facing left. Scale bars = 5 cm. cfr = caudal-fin rays; dcf = dorsal caudal fulcra; rcs = rhomboid caudal scales.

Figure 15. †Parapsephurus willybemisi n. gen. n. sp., digital reconstruction, from tomography data of body scales, of specimen FAU.DGS.ND.161.4559.T. Scale bar = 2 mm.
Holotype
FMNH PF17545 (Fig. 2), whole-body fossil preserved in three dimensions, including skull (with an associated rostrum), pectoral girdle and fins, median fins, and caudal fin (estimated 7 cm missing from posterior tip); estimated 160.5 cm total length; prepared from the right side.
Paratypes
FMNH PF17626, whole-body fossil preserved and prepared in ventral view, including portions of the rostrum, skull, pectoral girdle and fins, body, and pelvic fins; missing caudal peduncle and fin; FMNH PF17627, partially articulated skull and left pectoral girdle and fin, preserved in ventral view; FAU.DGS.ND.161.4559.T, posterior portions of the skull (lacking a rostrum) and right pectoral girdle and fin, exposed in right view.
Diagnosis
A species of the unranked polyodontid clade Polyodonti sensu Grande and Bemis (Reference Grande, Bemis, Stiassny, Parenti and Johnson1996) (i.e., the group that includes all fossil and living polyodontids except for †Protopsephurus, which lacks a well-developed series of stellate bones) that is distinguished from all other known species through a combination of the following characters: elongation of the parietals posteriorly (unlike the condition in †Paleopsephurus, in which the posterior margin is “non-emarginate … similar to acipenserids and other primitive actinopterygians but unlike other polyodontids,” Grande and Bemis, Reference Grande and Bemis1991, p. 85); having the middle fenestra longitudinalis bordered medially by the parietal and frontal and laterally by the dermopterotic (†Protopsephurus has only the frontal included in this margin, and in all other polyodontids this fenestra is bordered by the dermopterotic and dermosphenotic); slender and numerous dorsal caudal fulcra (unlike the condition in Psephurus, in which the dorsal caudal fulcra are robust and number 11 or fewer); an elongate hyomandibula that is not hourglass shaped (unlike the condition in †Crossopholis, in which the hyomandibula is hourglass shaped); and gill rakers that are short, stout, less numerous, and widely spaced (unlike the condition in Polyodon, in which the gill rakers are elongate, thin, more numerous, and very closely spaced).
Occurrence
Tanis Site (De Palma et al., Reference DePalma, Smit, Burnham, Kuiper and Manning2019) of the Hell Creek Formation, Bowman County, North Dakota, USA; Late Cretaceous (Maastrichtian). The rocks containing the holotype and FMNH paratypes were collected by Steve Nicklas and Robert Coleman in 2010.
Description
The holotype and most complete specimen of †Parapsephurus willybemisi n. gen. n. sp. best preserves the body outline of the fish. The rostrum is detached but measures 35 cm, which is 22% of the estimated total length. The insertion of the dorsal fin is slightly in advance of that of the anal fin. The caudal peduncle, although not well preserved, does not appear very long. The margin of the caudal fin is unclear beyond being heterocercal and forked, although its outline is suggestive of unequal lobes of the fin, with the dorsal lobe being longer than the ventral lobe. The unequal lobe morphology is like the condition in the Chinese Psephurus and unlike the condition in adult individuals of the North American Polyodon and †Crossopholis, in which the dorsal and ventral lobes are more or less equal in length. The equal length lobe state in adult Polyodon and †Crossopholis is likely derived within Polyodontidae, judging from ontogenetic studies showing juvenile Polyodon to have the unequal lobes (Bemis and Grande, Reference Bemis, Grande, Arratia and Schultze1999).
Dermal bones of the skull
The preserved portions of the skull roof of †Parapsephurus willybemisi n. gen. n. sp. include the parietals, frontals, dermopterotic, and dermosphenotics; anteriorly, these bones contact the median rostral bones, which are described in the following. In addition, there are small, irregularly shaped canal bones that are positioned posteriorly on the skull roof dorsal to the parietals that likely represent extrascapular bones, although nothing can be described of their arrangement (Figs. 3–5). There is slight ornamentation of the skull roofing bones, with low radiating ridges from the center of ossification and along the central axis of the parietals (Fig. 5). The parietals are well ossified, extend far posteriorly, and have a fimbriate posterior margin as in most other polyodontids. The left and right parietals contact each other (or nearly so) along much of the midline (the anterior end of these is unclear and may have defined a pineal foramen, as found in Psephurus). Anteriorly, the parietals contact the frontals, which are incompletely preserved in all of our specimens.
Laterally, the skull roof of †Parapsephurus willybemisi n. gen. n. sp. has two dermal bones: posteriorly the dermopterotic and anteriorly the dermosphenotic; in addition, the posttemporal is incorporated into the skull roof and is described in this section (Figs. 3–5). The posttemporal is most robust posterolaterally. Medially, there is a stout projection that contacts the ventral surface of the parietal at about the midpoint of the center of ossification of that bone (Fig. 5). The anterolateral arm of the posttemporal contacts the dermopterotic, forming the posterior margin of the posterior fenestra longitudinalis. The dermopterotic is robust and bears low, roughened ornamentation radiating out from its center of ossification. This bone contacts the parietal posteriorly through a slender posteromedial projection, defining the posterior margin of the middle fenestra longitudinalis. Anteriorly, the dermopterotic contacts the lateral margin of the frontal, with which it encloses the middle fenestra longitudinalis, although its exact relation to the frontal is not clearly preserved. This is different from what is found in most other polyodontids, in which the middle fenestra longitudinalis is defined medially by the dermopterotic and laterally by the dermosphenotic (e.g., see Grande and Bemis, Reference Grande and Bemis1991, figs. 32A, 50B, 67A for conditions in Psephurus, †Crossopholis, and †Paleopsephurus, respectively) and is similar to the condition in †Protopsephurus, in which the frontal forms the median margin of the middle fenestra longitudinalis (Grande et al., Reference Grande, Jin, Yabumoto and Bemis2002, fig. 7B); the condition in †P. willybemisi n. gen. n. sp. is unique, however, in that both the frontal and parietal form the median margin of the middle fenestra longitudinalis. Posteriorly, the dermopterotic contacts the posttemporal, and posterolaterally it contacts the dermosphenotic, with which it defines the posterior margin of the anterior fenestra longitudinalis. The precise shapes and extents of the anterior fenestra longitudinalis, the dermosphenotic, and the frontal are unknown.
The parasphenoid of †Parapsephurus willybemisi n. gen. n. sp. bears elongated posterior rami that extend beyond the pectoral girdle. These rami define an elongate aortic notch (Fig. 6) and are posteriorly pointed, unlike the more truncated or fimbriate ends in Psephurus and Polyodon, respectively. As in most polyodontids, the ascending processes are directed laterally to slightly anteriorly, extending from the main body of the parasphenoid at nearly right angles. This is in contrast to the condition in †Paleopsephurus, in which the ascending processes are posteriorly directed. A pair of elongate pharyngobranchials articulate with the parasphenoid at the base of the ascending processes (Fig. 6). The anterior tip of the parasphenoid contacts the median ventral bones of the rostrum and appears to continue anteriorly as a median anterior process, although the shape and extent of this process are not clearly preserved.
Dermal bones of the rostrum
A few rostral bones of †Parapsephurus willybemisi n. gen. n. sp. are present in FMNH PF17626 (Fig. 7), but the rostrum is most completely known from the holotype (FMNH PF17545; Fig. 8). The rostrum of the holotype is exposed in ventral view. This portion of the specimen was collected separately from the large block that contained the rest of the specimen; therefore, a portion of the bones connecting it to the rest of the skull is missing. In FMNH PF17626, the paired vomers are in contact with the anterior tip of the parasphenoid and are relatively long, smooth elements (Fig. 7). The vomers are not well preserved in the holotype due to the gap between the rostrum and the posterior portion of the skull, but those elements interpreted as the anterior tips of the vomers show them to be paired (Fig. 8). The more anterior bones of the ventral rostral skeleton comprise bones of the p, cv, and v series, which extend the length of the rostrum and, collectively, have lateral margins that are parallel. The p1–2 and cv1–2 bones are the longest elements of the rostrum, together extending to almost 70% of the preserved rostral length. The anterior cv3 and v2–3 bones are substantially shorter than the more posterior elements; there are additional splint-like bones near the anterior tip of the rostrum that are of unclear series.
There is a dense array of stellate bones present along the entire length of the preserved rostrum (Fig. 8). The stellate bones are well developed and largest medially, where as a series, they contact the median rostral bones. Laterally, the stellate bones are smaller, more irregularly shaped elements.
Opercular bones
The opercular bones of †Parapsephurus willybemisi n. gen. n. sp. include the subopercle (which is the largest and best-preserved element) and a single branchiostegal; a fragment of dermal bone positioned dorsally to the subopercle in FMNH PF17627 may represent a portion of an opercle, as found in basal polyodontids (e.g., Grande et al., Reference Grande, Jin, Yabumoto and Bemis2002) and non-acipenserid Acipenseriformes (Hilton and Grande, Reference Hilton, Grande and Schultze2021), but this requires additional material to confirm. The subopercle is preserved most completely in FMNH PF17627 (Fig. 9) and partially in FAU.DGS.ND.161.4559.T (Fig. 4). The subopercle is a largely flat, superficially ornamented bone with a broadly rounded posterior portion and a long, narrow anterior portion that extends from the midpoint of the main body of the subopercle (Fig. 9). The posterior margin of the subopercle of †Parapsephurus willybemisi n. gen. n. sp. does not bear the splint-like projections found in most other polyodontids. A single slender, elongate bone that is interpreted as a branchiostegal is preserved in FMNH PF17626 (Fig. 7) and FMNH PF17627 (Fig. 9). This bone is similar in shape to that found in Psephurus (Grande and Bemis, Reference Grande and Bemis1991, figs. 34, 36, 37) and †Paleopsephurus (Grande and Bemis, Reference Grande and Bemis1991, fig. 68) and unlike other polyodontids, which have a narrow, needle-like extension similar to the subopercle.
Jaws, suspensorium, and hyoid arch
The upper jaw of †Parapsephurus willybemisi n. gen. n. sp. comprises paired dermopalatines, palatopterygoids, and quadratojugals. The dermopalatine is a relatively narrow element that narrows anteriorly and forms the anterolateral margin of the upper jaw (for consistency with our other recent descriptions of the osteology of Acipenseriformes [Grande and Hilton, Reference Grande and Hilton2006; Hilton et al., Reference Hilton, Grande and Bemis2011, Reference Hilton, Dillman, Zhang, Zhang and Zhuang2016, Reference Hilton and Grande2023], we use the term “dermopalatine” here, with the acknowledgment that this element may represent the maxilla of other actinopterygians [Datovo and Rizzato, Reference Datovo and Rizzato2018]). At its posterior end, a well-developed ectopterygoid process extends posteromedially (Figs. 2, 10, 11.1–11.3) and appears to contact the palatopterygoid in the holotype. The palatopterygoid is a large, thin sheet of bone. There is no trace of an autopalatine. The posterior margin of the upper jaw is defined by the quadratojugal, which bridges the dermopalatine laterally and the palatopterygoid medially. The quadratojugal has elongate dorsal and ventral extensions that flare to broadly contact the other elements of the jaws. This is similar in form to the quadratojugal found in Psephurus (Grande and Bemis, Reference Grande and Bemis1991, fig. 35) and †Paleopsephurus (Grande and Bemis, Reference Grande and Bemis1991, fig. 69).
The longest bone of the lower jaw in †Parapsephurus willybemisi n. gen. n. sp. is the dentary, which is preserved in the holotype (FMNH PF 17545), as well as in FMNH PF17676, PF17627, and FAU.DGS.ND.161.4559.T. The dentary tapers anteriorly to a rounded, narrow tip that is bent medially at about one-fourth its length to meet its antimere at its symphysis. The dentary is deepest posteriorly but tapers anteriorly and ends posteriorly as a point. The mentomeckelian is best seen in FAU.DGS.ND.161.4559.T and is a conical element that sits in a median sulcus at the anterior tip of the dentary (Fig. 11.8). The prearticular is large and is anteriorly closely applied to the medial surface of the dentary. Posteriorly, the prearticular is separated from the dentary at a point of the jaw that presumably was filled with the articular portion of Meckel's cartilage in life (Fig. 11.5–11.7).
The hyomandibula, preserved only in the holotype of †Parapsephurus willybemisi n. gen. n. sp., is exceptionally long and is wider posteriorly than anteriorly (Fig. 3). The anterior head is narrow and is preserved in a position just posterior to the ascending process of the parasphenoid. The hyomandibula extends from this point at about a 45° angle posteroventrally. It is shaped much like the combined ossified and cartilaginous portions of the hyomandibula of Psephurus (Grande and Bemis, Reference Grande and Bemis1991, fig 34). The interhyal is large, robust, and hour-glass shaped (Figs. 3, 7). The anterior ceratohyal is also large, and there is a smaller irregularly shaped element tentatively identified as the posterior ceratohyal; this element has been displaced and separated from the anterior ceratohyal, but it is associated with the proximal tip of the interhyal (Fig. 7). The left hypohyal is present and is associated with the anterior tip of the anterior ceratohyal in FMNH PF17676; it is small, roughly triangular, and has a rounded anterior end (Fig. 7).
Gill arches
In general, the gill arches of †Parapsephurus willybemisi n. gen. n. sp. are poorly preserved. Although portions of the certatobranchial series (Figs. 3, 7, 12) and pharyngobranchial series (Fig. 5) are preserved, not much detail can be determined. The gill rakers are preserved and exposed in the holotype (FMNH PF17545) as well as in FAU.DGS.ND.161.4559.T (Figs. 3, 12). The gill rakers are relatively short and widely spaced, unlike the gill rakers of Polyodon but similar to all other polyodontids for which these structures are known (see Grande and Bemis, Reference Grande and Bemis1991, fig. 26 for comparisons of gill-raker morphology among polyodontids and other acipenseriforms for which these structures are known). The gill rakers appear to be superficially ossified elements that surrounded a soft-tissue core (possibly cartilage) as there is a space in the center of each element. Anteriorly and posteriorly, each gill raker is flared out proximally where they contact the underlying branchial elements (Fig. 12.3).
Vertebral column
Nothing is known of the vertebral elements of †Parapsephurus willybemisi n. gen. n. sp.
Dorsal and anal fins and supports
The dorsal and anal fins and their supports are preserved only on the holotype of †Parapsephurus willybemisi n. gen. n. sp. (Fig. 13). There are approximately 25 dorsal-fin rays, although posteriorly these become difficult to count. There are nine proximal radials and about four or five middle radials preserved supporting the dorsal fin, but these series are incomplete.
There are at least 32 anal-fin rays in †Parapsephurus willybemisi n. gen. n. sp. supported by 14 middle radials (Fig. 13); there are five proximal radials supporting the anal fin, but this series, as for the dorsal fin, is incomplete.
Caudal fin and supports
The caudal fin is preserved only on the holotype of †Parapsephurus willybemisi n. gen. n. sp., which shows a typically heterocercal tail with a dorsal lobe that is longer than the ventral lobe (Fig. 14). Although present, the caudal-fin rays are not sufficiently preserved to provide counts. There are at least 20 dorsal caudal fulcra preserved along the dorsal margin of the caudal fin. These elements are more slender than those in most other polyodontids. The rhombic caudal scales are present but are largely displaced. However, these do appear smaller and slenderer than those of comparably sized specimens of other polyodontids.
Pectoral and pelvic girdles and fins
The preserved portions of the pectoral girdle of †Parapsephurus willybemisi n. gen. n. sp. comprise the posttemporal, supracleithrum, postcleithrum, cleithrum, and fragments of the clavicle. The supracleithrum is preserved on the holotype but displaced from its presumed articulation with the posttemporal. The supracleithrum is an elongate bone that is slightly broader toward its anterior end than its posterior end before tapering to a sharp anterior point (Fig. 3). A smooth crescent-shaped bone that we interpret as a postcleithrum is present between the posterior end of the supracleithrum and cleithrum but is clearly not in its natural position. The cleithrum is well preserved in the holotype and resembles that of Psephurus in its overall form. There is no distinct notch along its posterior margin (e.g., as in †Crossopholis), but it does have a shallow concavity that presumably indicates where the pectoral fin inserts (this concavity is absent in Polyodon). The dorsal tip of the cleithrum appears notched (Fig. 3). In FAU.DGS.ND.161.4559.T, the cleithrum broadens medially to form the dorsal portion of the opercular wall, as found in other Acipenseriformes (Fig. 4). The clavicle is best preserved in FMNH PF17672 (Fig. 7), in which it is seen in ventral view. The left and right clavicles essentially meet along the midline and make broad contact with the cleithra laterally (Fig. 7), as in Polyodon (Grande and Bemis, Reference Grande and Bemis1991, fig. 20). We did not find an interclavicle, but this is possibly due to lack of preservation. The pectoral fin of the holotype has at least 23 fin rays preserved, and FMNH PF17627 has at least 27 fin rays preserved, but it appears to be missing some medially. The leading fin ray is larger than the more posterior ones but does not appear to be spine-like, as in †Protopsephurus (Grande et al., Reference Grande, Jin, Yabumoto and Bemis2002).
No traces of the pelvic girdle or fin are preserved on the holotype. In FMNH PF17626, there are at least 22 right pelvic-fin rays preserved, but this fin appears incomplete; there are at least five left pelvic-fin radials preserved, but again, this series appears incomplete.
Body scales
The body scales of †Parapsephurus willybemisi n. gen. n. sp. are small and scattered along the length of the holotype (FMNH PF17545) and are well preserved just posterior to the pectoral girdle in FAU.DGS.ND.161.4559.T (Fig. 4). These are the denticular trunk scales of Grande and Bemis (Reference Grande and Bemis1991) and consist of a robust base with one to three spines projecting from the base (Fig. 15). We found no round-based scales associated with the opercular wall formed by the cleithrum and clavicle, as found in other acipenseriforms, although this may be due to lack of preservation.
Etymology
willybemisi, named in honor of our colleague Dr. William (Willy) E. Bemis for his many contributions to the systematic biology of fossil and living polyodontid fishes.
Remarks
Aspects of the neurocranium of †Parapsephurus willybemisi n. gen. n. sp. are preserved in FAU.DGS.ND.161.4559.T and are the focus of an ongoing study (During et al., unpublished data, 2022).
Genus †Pugiopsephurus new genus
Type species
†Pugiopsephurus inundatus n. gen. n. sp., by monotypy.
Diagnosis
As for species; monotypic.
Etymology
Combination of Pugio, Latin, meaning “dagger” in reference to the shape of the rostrum, and psephurus, a Recent genus of Polyodontidae, in reference to the recently extinct polyodontid genus Psephurus.
Remarks
The holotype and only known specimen of †Pugiopsephurus inundatus n. gen. n. sp. is preserved on a block of stone that also includes a nearly complete specimen of †Acipenser praeparatorum Hilton and Grande, Reference Hilton and Grande2022 (DMNH EVP.138511) and three isolated polyodontid rostra described in the following as Polyodontidae gen. indeterminate sp. indeterminate.
†Pugiopsephurus inundatus new species
Figures 16–18

Figure 16. † Pugiopsephurus inundatus n. gen. n. sp. DMNH EPV.138508 (holotype and only known specimen). Anterior facing left. Scale bar = 5 cm.

Figure 17. †Pugiopsephurus inundatus n. gen. n. sp. Skull and pectoral girdle, DMNH EPV.138508 (holotype and only known specimen). (1) Photograph. (2) Line drawing. Anterior facing left. Scale bars = 5 cm. afl = anterior fenestra longitudinalis; c3 = paired outer lateral dorsolateral bones of the rostrum; cv1–3 = paired outer lateral ventrolateral bones of the rostrum; dpl = dermopalatine; dpt = dermopterotic (= intertemporo–supratemporal of Grande and Bemis, Reference Grande and Bemis1991; see Grande et al., Reference Grande, Jin, Yabumoto and Bemis2002); dsp = dermosphenotic; fr = frontal; mfl = middle fenestra longitudinalis; p1–3 = unpaired ventral rostral splints belonging to the median series; pas = parasphenoid; pfr = pectoral-fin rays; qj = quadratojugal; v = vomer; v2 = paired ventral rostral splints; (l) = left; (r) = right.

Figure 18. †Pugiopsephurus inundatus n. gen. n. sp. Body scales, DMNH EPV.138508 (holotype and only known specimen). Anterior facing left. Scale bar = 10 mm.
Holotype
DMNH EPV.138508 (only known specimen; Fig. 16). Portions of the skull, rostrum, pectoral girdle, and fin and anterior portion of body exposed in ventral view. Preserved portion of the skull measures 320 mm.
Diagnosis
A member of Polyodontidae that is diagnosed by a combination of characters: dermopalatine with medial expansion (unlike all polyodontids except †Protopsephurus) and lacking an ectopterygoid process (unlike all polyodontids except Polyodon). The only known specimen appears to lack stellate bones; if this is supported by the discovery of additional specimens, this condition would be unique among polyodontids.
Occurrence
Tanis Site (De Palma et al., Reference DePalma, Smit, Burnham, Kuiper and Manning2019) of the Hell Creek Formation, Bowman County, North Dakota, USA; Late Cretaceous (Maastrichtian).
Description
In this section, we provide a description of the anatomy of †Pugiopsephurus inundatus n. gen. n. sp. as far as is possible from the single known specimen. Only portions of the skull and anterior body are preserved, and nothing is known of the skull roof, the opercular series, much of the jaws, suspensorium, hyoid arches, gill arches, vertebral column, pelvic girdle and fins, dorsal and anal fins and their supports, or the caudal fin and skeleton.
Dermal bones of the skull
The bones of the skull roof of †Pugiopsephurus inundatus n. gen. n. sp. that are best preserved include the dermosphenotic and the dermopterotic (Fig. 17). The left dermopterotic is more complete than its antimere and may also show the anterior portion of the posterior fenestra longitudinalis, although this is not clear. The dermosphenotic and dermopterotic contact one another through a slender bridge that separates the anterior and middle fenestrae longitudinalis. This bridge is more slender than that of †Parapsephurus willybemisi n. gen. n. sp. The dermosphenotic and c3 bone laterally and the dermopterotic medially define the large anterior fenestra longitudinalis. Portions of the left frontal are preserved and exposed in ventral view, but few details are available.
Portions of the parasphenoid are preserved, including the tips of the posterior rami, which reach to the level of the pectoral fins. Anteriorly, the parasphenoid is exposed and contacts the paired vomers. The parasphenoid tapers anteriorly, but a median anterior process is not distinctly preserved.
Dermal bones of the rostrum
Portions of the dorsal and ventral rostral bones are preserved, including the c3 bones, which are in contact with the anterior tip of the dermosphenotics. Other lateral rostral bones are dorsally and ventrally in contact with the paired vomers, the cv1–3 bones, and the p2–4 bones. There is no indication of stellate bones. The c3 bone does not have a posterior notch, as that of other polyodontids (e.g., see Grande and Bemis, Reference Grande and Bemis1991), but rather contacts the dermosphenotic through a lap suture. Anterior and lateral to the c3 bone, there is a series of bones that are trough-like and may be interpreted as canal bones, carrying the infraorbital canal anteriorly. The paired vomers are elongate and extend over one-third the length of the rostrum. The median p1 bone separates the left and right vomers along much of their length; p2–4 bones are progressively shorter. The cv1–3 bones are of similar lengths and are positioned lateral to the p series.
Jaws, suspensorium, and hyoid arch
The only elements of the jaws, suspensorium, and hyoid arch that are preserved in the single specimen of †Pugiopsephurus inundatus n. gen. n. sp. are the left and right dermopalatines, the right quadratojugal, and fragments of the right dentary and other unidentified bones of the suspensoria (Fig. 17). The right quadratojugal is incompletely preserved, but it appears to have the anterior flaring found in Psephurus and some other polyodontids, although the extent and shape of these is unknown. Two bones are identified as the left and right dermopalatines because they are positioned closely to the ventral surface of the parasphenoid (i.e., there are no intervening elements, so it is unlikely these are the dentaries). These bones are exposed in ventral view and appear to be more or less complete. Both elements are bent near their midpoints at about a 30° angle, although their anterior margins are smoothly curved and appear to have a distinct medial expansion, similar to though not as extreme as in the dermopalatines of †Protopsephurus and most Acipenseridae. There is no trace of an ectopterygoid process.
Pectoral and pelvic girdles and fins
Portions of the pectoral girdle are represented by fragments of smooth dermal bone scattered posterior to the skull; no details can be provided. There are 31 pectoral-fin rays present on the right fin, which appears complete; 25 fin rays are preserved in the left pectoral fin, although this fin-ray count appears incomplete medially.
Body scales
Denticulate scales are present (Fig. 18) and are most abundant on the sides of the anterior portion of the body and sparser along the midline; they are largest immediately behind the pectoral girdle and decrease in size posteriorly (the posterior portion of the body is not preserved, but the density of scales is less posteriorly on the preserved portion, suggesting an anterior-to-posterior gradation). The scales are roughly triangular in shape, with the widest edge facing anteriorly.
Etymology
inundatus, Latin, adjective, meaning flooded, in reference to the seiche wave that has been implicated in the mass burial of fishes at the Tanis site.
Remarks
The absence of stellate bones in the unique specimen is possibly an artifact of lack of preservation or ontogenetic variation and needs to be confirmed in additional specimens.
Polyodontidae genus indeterminate species indeterminate
Figure 19
Remarks
Within the available polyodontid material from the Tanis Site are additional specimens of Polyodontidae, although they cannot be diagnosed to any particular taxon beyond the family level. These include three isolated rostra of varying lengths (Fig. 19). The smallest of the three (DMNH EPV.138510; preserved length 320 mm) is from an individual of comparable size to the holotype of †Pugiopsephurus inundatus n. gen. n. sp. and consists of the anterior portion of the skull and rostrum, exposed in dorsolateral view (Fig. 19.1). In addition to the bones of the rostrum, which are ornamented with very fine rows of denticle-like ornamentation arranged along the long axis of the bone, portions of the frontal, dermopterotic, and dermosphenotic are preserved, defining the anterior extent of the anterior fenestra longitudinalis. The suture between the dermopterotic and the dermosphenotic is narrow and formed by the extensions of both bones. A portion of the lower jaw is also preserved, but no details can be observed. DMNH EPV.138509 (Fig. 19.2) is from a slightly larger individual than DMNH EPV.138510 and is also exposed in dorsolateral view (preserved length 405 mm). Dermal bones that likely represent portions of the frontal, dermopterotic, and dermosphenotic are preserved but too fragmentary to be informative. The rostral bones of this specimen have similar ornamentation to DMNH EPV.138510, consisting of longitudinal rows of denticle-like ridges. Both specimens appear to lack stellate bones. This absence does not appear to be a factor of lack of preservation or preparation as equally fine bones (e.g., canal bones) are preserved. Rather, this absence is likely either taxonomic or the result of ontogenetic variation.

Figure 19. Isolated rostra of Polyodontidae indeterminate (specimen above and close-up of tip of rostrum in inset). (1) DMNH EPV.138510; preserved length 320 mm. (2) DMNH EPV.138509; preserved length 405 mm. (3) DMNH EPV.138512; preserved length 515 mm; note stellate bones. Anterior facing left. Scale bars = 5 cm.
The largest of the three indeterminate polyodontids examined (DMNH EPV.138512) is a rostrum exposed in ventral view; the preserved length of this specimen is 515 mm. The bones of the rostrum are, for the most part, not well preserved, such that identification of individual bone according to their series is not possible. However, an array of stellate bones is preserved along the length of the rostrum and is particularly dense at the anterior tip of the rostrum (Fig. 19.3).
Discussion
The two species of paddlefishes described in this paper add to the morphological and known taxonomic diversity of Polyodontidae, a family first represented in the known fossil record by the 130-million-year-old †Protopsephurus liui from China (Grande et al., Reference Grande, Jin, Yabumoto and Bemis2002). †Protopsephurus is a relatively basal “sturgeon-like” polyodontid, yet it displays several characteristics that firmly place it within Polyodontidae, including the presence of anterior, middle, and posterior fenestrae longitudinalis and the presence of stellate bones, albeit these are weakly developed. †Parapsephurus willybemisi n. gen. n. sp. shares with †Protopsephurus many plesiomorphic characters (e.g., short, widely spaced gill rakers) but has a well-developed series of stellate bones, similar to more-derived polyodontid taxa. The unique and incomplete specimen of †Pugiopsephurus inundatus n. gen. n. sp. also includes a number of plesiomorphic characters found in †Protopsephurus, such as absence of stellate bones, as well as more-derived polyodontid characteristics, such as the large anterior and middle fenestrae longitudinalis. The phylogenetic information of these characters awaits a full analysis of the relationship among fossil and living Polyodontidae, which is beyond the scope of this paper. The named representation of the family in the Late Cretaceous of North America now stands at three monotypic genera (†Pugiopsephurus n. gen.; †Parapsephurus n. gen.; †Paleopsephurus; see Grande and Bemis, Reference Grande and Bemis1991; Hilton and Grande, Reference Hilton, Grande and Schultze2021), all from the Hell Creek Formation. Fossil polyodontid diversity in North America extends into the Paleogene, including the species †Polyodon tuberculata from the early Paleocene Tullock Formation (Montana) and the monotypic genus †Crossopholis from the early Eocene Green River Formation (Wyoming). It will be important to reassess the phylogenetic relationships of fossil and living polyodontids to better understand the evolutionary history of not only the taxa, but also the evolution of the morphological characteristics associated with this family. For example, the absence of stellate bones in †Pugiopsephurus inundatus n. gen. n. sp. needs to be confirmed with additional specimens. If stellate bones are truly absent, this absence represents either a plesiomorphic condition or a reversal. We note that the other rostral elements of the genus are more elongate and its rostrum is elongated and similar to all other polyodontids except †Protopsephurus, which has a much shorter, acipenserid-like rostrum and weakly developed stellate bones.
The presence of three paddlefish species within the Late Cretaceous Hell Creek Formation of North America, and one from the Early Cretaceous of China, hints at substantial diversity of polyodontids already by the Cretaceous. Their biogeographic distribution, confined to freshwaters or freshwater deposits of East Asia and North America, indicate a trans-Pacific, rather than a trans-Atlantic, pattern of historical relationship dating back to the Early Cretaceous, when the two continental regions were in close contact (Grande, Reference Grande1985; Grande and Bemis, Reference Grande and Bemis1991). There are probably more polyodontid fossil taxa waiting to be discovered in China. In addition, there are several polyodontid indeterminate records from the Late Cretaceous Hell Creek deposits, as well as the proximate formations of the Judith River Group (Grande and Bemis, Reference Grande and Bemis1991), that may be better interpreted with the discovery of better-preserved material.
Acknowledgments
We thank C. Van Beek and K. Passaglia for preparation of the FMNH specimens of the two species described herein and J. Weinstein for photographic assistance. T. Larson and K. Mackenzie graciously provided access to the DMNH specimen examined; N. Nev-Yagle prepared the DMNH specimen. We thank A. Murray and an anonymous reviewer for their comments that improved the manuscript. This study was supported by the US National Science Foundation (DEB-1754483) and grants from the Negaunee Foundation. E.J.H. was also supported by a Bass Fellowship from the Field Museum that supported an extended visit to the Field Museum. M.A.D.D. and P.E.A. were supported by the Swedish Research Council (2020-03685).
Declaration of competing interests
The authors declare none.




















 Open access
Open access