



Exploitation of geophytes (perennial plants that store edible starch in underground storage organs, or “USOs”) has been well documented in the ethnographic literature in western North America. During their 1804–1806 expedition along the Missouri and Columbia Rivers and across the Rockies, Lewis and colleagues (Reference Lewis, Clark and Biddle1814) noted roots and tubers from several different plant taxa—such as cattails (Typha), rush (Carex), wapato (Sagittaria), and thistle (Cirsium)—that were commonly collected and consumed by the Northwest and Plains Indian tribes as well as the Northern Shoshone in southern Idaho. Geophytes commonly consumed by Native American tribes of the Great Basin (e.g., Northern Paiute, Southern Paiute, Ute, Shoshone, Goshute) include the taproots or rootstocks of biscuitroot (Lomatium), yampah (Perideridia), bitterroot (Lewisia), springparsley (Cymopterus); bulbs of fritillary (Fritillaria), wild onion (Allium), sego lily (Calochortus); and corms of Brodiaea-type geophytes (Dichelostemma; Chamberlin Reference Chamberlin1909, Reference Chamberlin1911; Couture et al. Reference Couture, Ricks and Housley1986; Kelly Reference Kelly1932, Reference Kelly1934; Lawton et al. Reference Lawton, Wilke, DeDecker and Mason1976; Steward Reference Steward1933). Several tribes in the American Southwest (e.g., Apache, Navajo, Hopi, Pueblo, Zuni) also collected geophytes, including rootstocks of springparsley, tubers of wild potato (Solanum jamesii), horsenettle (Solanum fendleri), and bulbs of sego lily and wild onion (Castetter Reference Castetter1935; Castetter and Opler Reference Castetter and Opler1936; Fewkes Reference Fewkes1896; Swank Reference Swank1932; Vestal Reference Vestal1940, Reference Vestal1952).
Geophytes are known to be a highly valued food item, but they also have socioecological implications for human societies across the globe (Fulkerson and Tushingham Reference Fulkerson and Tushingham2021; O'Connell et al. Reference O'Connell, Hawkes and Blurton Jones1999). Archaeologists working in southern Wyoming hypothesize that these predictable and nutritious plant resources encouraged a stable, long-term pattern of land use that ultimately resulted in sustaining prehistoric human populations throughout the Holocene (Larson Reference Larson1997; Smith Reference Smith2003; Smith and McNees Reference Smith and McNees2011). Although return rates for geophytes in western North America vary widely, some (e.g., Lomatium) have a ranking similar to avian and small mammal resources, and their encounter rates are less variable than those of large, mobile prey (Couture et al. Reference Couture, Ricks and Housley1986; Smith and McNees Reference Smith and McNees2005; Zeanah Reference Zeanah2004; Figure 1). Unfortunately, there is difficulty in finding archaeological evidence of geophyte use because the fleshy underground storage organs do not preserve well compared to other plant parts that are hard and dense and/or charred (Lepofsky and Peacock Reference Lepofsky, Peacock and Prentis2004). Most archaeologists, therefore, rely on indirect evidence such as pits, slab-lined hearths, flake tool morphology, digging sticks, or digging stick weights to infer consumption (Bradley et al. Reference Bradley, Smith and Wriston2020; Dering Reference Dering1999; Eerkens and Rosenthal Reference Eerkens and Rosenthal2002; Lepofsky and Peacock Reference Lepofsky, Peacock and Prentis2004; McGuire and Stevens Reference McGuire, Stevens, Hildebrandt, McGuire, King, Ruby and Young2016; Pool Reference Pool2001; Sutton Reference Sutton2014; Thoms et al. Reference Thoms, Short, Kamiya and Laurence2018). Although digging sticks are suggestive, they often do not preserve well, and their size and shape do not indicate which plant species were being utilized. Likewise, if flake tools are used to infer the manufacture of digging sticks, then this becomes indirect evidence of indirect evidence.

Figure 1. Post-encounter return rates (kcal/hour) for dietary resources common across western North America. Return rate values are compiled from the published literature (e.g., Broughton et al. Reference Broughton, Cannon, Bayham and Byers2011; Couture et al. Reference Couture, Ricks and Housley1986; Diehl and Waters Reference Diehl, Waters, Kennett and Winterhalder2006; Gremillion Reference Gremillion2004; Kelly Reference Kelly1995; Rhode and Rhode Reference Rhode and Rhode2015; Simms Reference Simms1987; Smith and McNees Reference Smith and McNees2005; Smith et al. Reference Smith, William and Johansen2001; Ugan and Rosenthal Reference Ugan and Rosenthal2016).
Burned remains of geophytes provide direct evidence of use and have been found in archaeological contexts (Dering Reference Dering1999; Gill Reference Gill2016; Gill et al. Reference Gill, Braje, Smith and Erlandson2021; Lyons and Ritchie Reference Lyons and Ritchie2017; Thoms Reference Thoms1989). Starch granule analysis also provides direct evidence of geophyte use and may allow identification—sometimes down to species level—of plants processed on or cooked in implements, such as baskets, ground stone, or pottery (e.g., Herzog and Lawlor Reference Herzog and Lawlor2016; Rankin Reference Rankin2016; Rhode and Rankin Reference Rhode, Rankin and Thomas2020; Louderback and Pavlik Reference Louderback and Pavlik2017). Even though ethnographic evidence suggests that geophytes were an important food source to Native peoples living in southern Wyoming (Steward Reference Steward1933), there is virtually no support in the archaeological record. However, there appears to have been processing of Lomatium and Lewisia at High Rise Village in high elevations of the Wind River Range, Wyoming (Rankin Reference Rankin2016).
A common geophyte species found near and within archaeological sites in southern Wyoming is Cymopterus bulbosus A. Nelson (bulbous springparsley). Its starchy taproot (Figure 2) could have been an important food source for prehistoric humans living in the region. For example, traditional processing methods suggest that C. bulbosus was eaten raw after bark was peeled from the taproot, or roasted in ashes, cooked in milk, or dried and ground for overwintering (Castetter Reference Castetter1935; Vestal Reference Vestal1952). Furthermore, experimental data show that the caloric return rates of C. bulbosus were enough to support seasonal exploitation by foragers (Smith and McNees Reference Smith and McNees2005). Yet, there has been no direct evidence for the use of this geophyte from the archaeological record.

Figure 2. Cymopterus bulbosus with starchy taproots. Plants were collected in June 2017 from patches adjacent to sites 48UT375 and 48SW8842 in the Wyoming Basin (photograph courtesy of Erick Robinson). (Color online)
In this study, starch granule analysis on ground stone tool collections from older sites located in lowland settings was conducted to provide further evidence for geophyte exploitation in southern Wyoming. Focusing specifically on C. bulbosus, we systematically measured granules from modern reference specimens to aid in the identification of archaeological granules. Starch granules were extracted from 10 manos and metates found in two stratified, multicomponent archaeological sites—48UT375 and 48SW8842—in the Wyoming Basin to contribute to a more thorough understanding of resource consumption patterns throughout the Holocene in this region.
Study Sites
Site 48UT375
Site 48UT375 is a multicomponent site situated on a low interfluvial ridge near the ephemeral drainage of Austin Wash, approximately 13 km northeast of Lyman in Uinta County, Wyoming (Reust et al. Reference Reust, Richard, Eckerle, Taddie, Smith and Newton2002; Smith et al. Reference Smith, Reust and Richard2003; Figure 3). Aeolian deposits up to 118 cm deep yielded four separate components: Late Paleoindian, Paleoindian / Early Archaic (Great Divide Phase), Middle Archaic (Pine Spring Phase), and Late Archaic / Late Prehistoric (Reust et al. Reference Reust, Richard, Eckerle, Taddie, Smith and Newton2002). The focus of this study will be on Components 1 (Late Paleoindian, ~9400–9600 cal BP) and 3 (Middle Archaic, ~4400–4900 cal BP) because they are the two components that yielded ground stone tools. Geophyte consumption was suggested at site 48UT375 due to the presence of ground stone tools, the abundance of fire-cracked rock, and the dense patches of geophytes surrounding the site, although no direct evidence was recorded at the time (Smith et al. Reference Smith, Reust and Richard2003).

Figure 3. Location of sites 48UT375 and 48SW8842 in Green Basin, Wyoming. Cymopterus bulbosus populations were located in the immediate vicinity (<200 m) of the sites. Bottom-middle photo shows field crew collecting C. bulbosus at Population 1, near site 48UT375 (photograph courtesy of Bruce Pavlik). (Color online)
Site 48SW8842
Site 48SW8842 is a deeply stratified site situated in sand shadow deposits in the lee of a sandstone ridge overlooking a playa flat that is a tributary of North Barrel Springs Draw, approximately 24 km south of Wamsutter in Sweetwater County, Wyoming (Pool Reference Pool2001; Figure 3). Five stratified cultural levels were excavated, defined as “analytical units”: AU1 (Late Prehistoric Uinta Phase), AU2 (Late Archaic Pine Spring Phase), AU3 (Early Archaic Opal Phase), AU4 (Early Archaic Great Divide Phase), and AU5 (Paleoindian and transitional Paleoindian / Early Archaic Great Divide Phase). This study will focus on ground stone tools recovered form AU1, AU2, AU3, and AU4, ranging in age from approximately 1400 to 8300 cal BP. It has been suggested that several of the slab-lined pits and hearths excavated from 48SW8842 were used for the preparation and baking of geophytes (Pool Reference Pool2001).
Material and Methods
Ground Stone Assemblage
The ground stone collections from sites 48UT375 and 48SW8842 are accessioned at the University of Wyoming Archaeological Repository. Ten ground stone tools that span the Holocene in age (1400–9600 cal BP) were examined for starch granules. Ground stone artifacts from 48UT375 (three manos) were all manufactured from quartzite, whereas at 48SW8842 (one mano, six metates), were manufactured from sandstone. These artifacts were chosen because they occur in multiple levels spanning the Early Archaic to Late Prehistoric times, including a mano from 48UT375 that dates to around 9400–9600 cal BP. Full descriptions of the ground stone tools are in Reust and colleagues (Reference Reust, Richard, Eckerle, Taddie, Smith and Newton2002) and Pool (Reference Pool2001), and images of the 10 tools examined in this study are in Supplemental Figure 1.
Starch Extraction from Reference Materials
Reference material for this study includes a total of six C. bulbosus plants collected from two populations (n = 3 individuals from each population) in southwestern Wyoming (Figure 3). Population 1 is located approximately 170 m west and 150 m north of site 48UT375, and Population 2 is situated about 170 m west and north of site 48SW8842. Cymopterus bulbosus specimens that were collected in the field were brought to the Natural History Museum of Utah (NHMU) Archaeobotany Lab for sample preparation and processing. Cortex and stele of the taproots were isolated and ground with a small amount of 50/50 glycerol and DH2O solution in a sterile tissue homogenizer and mounted onto glass slides.
Starch Extraction from Ground Stone Artifacts
A portion of the worked surface from each ground stone artifact was sonicated for three minutes in a sterile weigh boat with a small amount of deionized (DH20) water. The samples were then sieved through a 125 μm mesh Endecott sieve to remove material greater than 125 μm. The samples (<125 μm) were then transferred to a 50 mL tube and centrifuged for 5 minutes at 3,000 RPM, and the supernatant was discarded. The pellets were resuspended and transferred to 15 mL tubes and centrifuged for 5 minutes at 3,000 RPM, and the supernatant was decanted. Heavy liquid was used to isolate starch granules from the samples; 5 mL of lithium heteropolytungstate (LST, specific gravity 2.00) was added to each sample and resuspended with a vortex mixer. The samples were set aside for 20 minutes to allow starch granules and other organics to float to the top. The top layer of organics was then removed with a pipette and placed in new 15 mL tube. Samples were rinsed twice with 10 mL of DH2O and centrifuged for 3 minutes at 3,000 RPM in order to remove any residual heavy liquid. A final rinse was performed with 7 mL of acetone and centrifuged for 5 minutes at 3,000 RPM. The supernatant was decanted, and the pellets were left to dry overnight. The pellets were mixed with a small amount of 50/50 glycerol and DH2O solution and mounted on glass slides.
Microscopy
Each slide was scanned using a transmitted brightfield microscope fitted with polarizing filters and Nomarski optics (Zeiss Axioskop 2, Zeiss International, Göttingen, Germany). A digital camera (Zeiss HRc) with imaging and measurement software (AxioVision) were used to capture images of and measure starch granules. For each reference sample, randomly generated X and Y coordinates on the microscope stage were used to measure and photograph approximately 100 granules from each individual plant (n = ~300 from each population). All starch granules present in these photographs were measured and examined for morphological features. Photograph locations were distant enough from one another to ensure that starch granules were not photographed twice.
To examine surface features and three-dimensional shapes of starch granules, scanning electron microscopy (SEM) was also used. A small portion of each processed sample was mounted directly onto aluminum stubs using double-sided carbon tape. An FEI Quanta 600F SEM (FEI Company, Hillsboro, Oregon, USA) with a high-resolution field-emission source was used to image starch granules.
Granule Size and Morphology
The size of each starch granule was measured as the maximum length through the hilum. Granules were also examined for the presence of morphological features including lamellae, hilum position, angular margins, fissures at the hilum, pressure facets, and depressions along the margins (International Code for Starch Nomenclature [ICSN] 2011; Reichert Reference Reichert1913). Starch granule size tends to be non-normally distributed and, therefore, relying on mean granule size is not appropriate for identification purposes (Louderback et al. Reference Louderback, Herzog and Pavlik2017). To assess the normality of granule size distributions, the Shapiro-Wilk test was employed and reported as p-values using the R statistical environment (R Core Team 2019). P-values less than 0.05 indicate that the distributions deviate significantly from normal.
Applying a statistical analysis to starch granule identification promotes reproducibility and an overall increase in confidence (Gao et al. Reference Gao, Moussavi and Myers2021; Louderback et al. Reference Louderback, Herzog and Pavlik2017). It is becoming standard practice to measure the size, shape, and morphological features of reference starch granules from multiple populations so that variation and statistical significance can be assessed (see Brown and Louderback Reference Brown and Louderback2020; Liu et al. Reference Liu, Ma and Cui2014; Wilks et al. Reference Wilks, Louderback and Boomgarden2021).
Results
Reference Material
Granule Size
Starch granule lengths were not normally distributed in all reference specimens except for individual B (p = 0.4). Therefore, all measurements have been log transformed for calculation of descriptive statistics (Table 1). Starch granule measurements from individual plants were then pooled for overall size range and morphological analyses. Smaller granules generally exhibit fewer diagnostic features (Louderback et al. Reference Louderback, Herzog and Pavlik2017), so the top 20% fraction was used for taxonomic identification. Although granule length was slightly different between the two populations, it is not statistically significant (t(4) = 0.611, p = 0.57).
Table 1. Descriptive Statistics and Shapiro-Wilk Normality Test of Starch Granule Lengths of Individual Plants (A, B, C, X, Y, Z) and Populations (1 and 2).

Note: All measurements have been log transformed except for Population 1B, which was normally distributed.
Granule Morphology
Morphological features including hilum position, lamellae, granule shape, fissures, pressure facets, and depressions were recorded for each granule (Figure 4). Although these features have long been known and previously described (ICSN 2011; Reichert Reference Reichert1913), some had to be refined for the present study. For example, pressure facets are large, obvious indentations that develop in compound granules common to Poaceae, Apiaceae, and other plant families (Figures 4b and 4e). Depressions (“pits”), however, are small, subtle indentations that may or may not be the result of compound granule formation (ICSN 2011; Figures 4c and 4f). When viewed in transmitted light (two dimensions), depressions on spherical granules (e.g., Cymopterus) can only be seen when positioned along the outer margin. But on the surface of flattened, lenticular granules (e.g., Triticeae grasses, such as Leymus, Hordeum) they are readily observed (Brown and Louderback Reference Brown and Louderback2020; Perry and Quigg Reference Perry and Quigg2011).
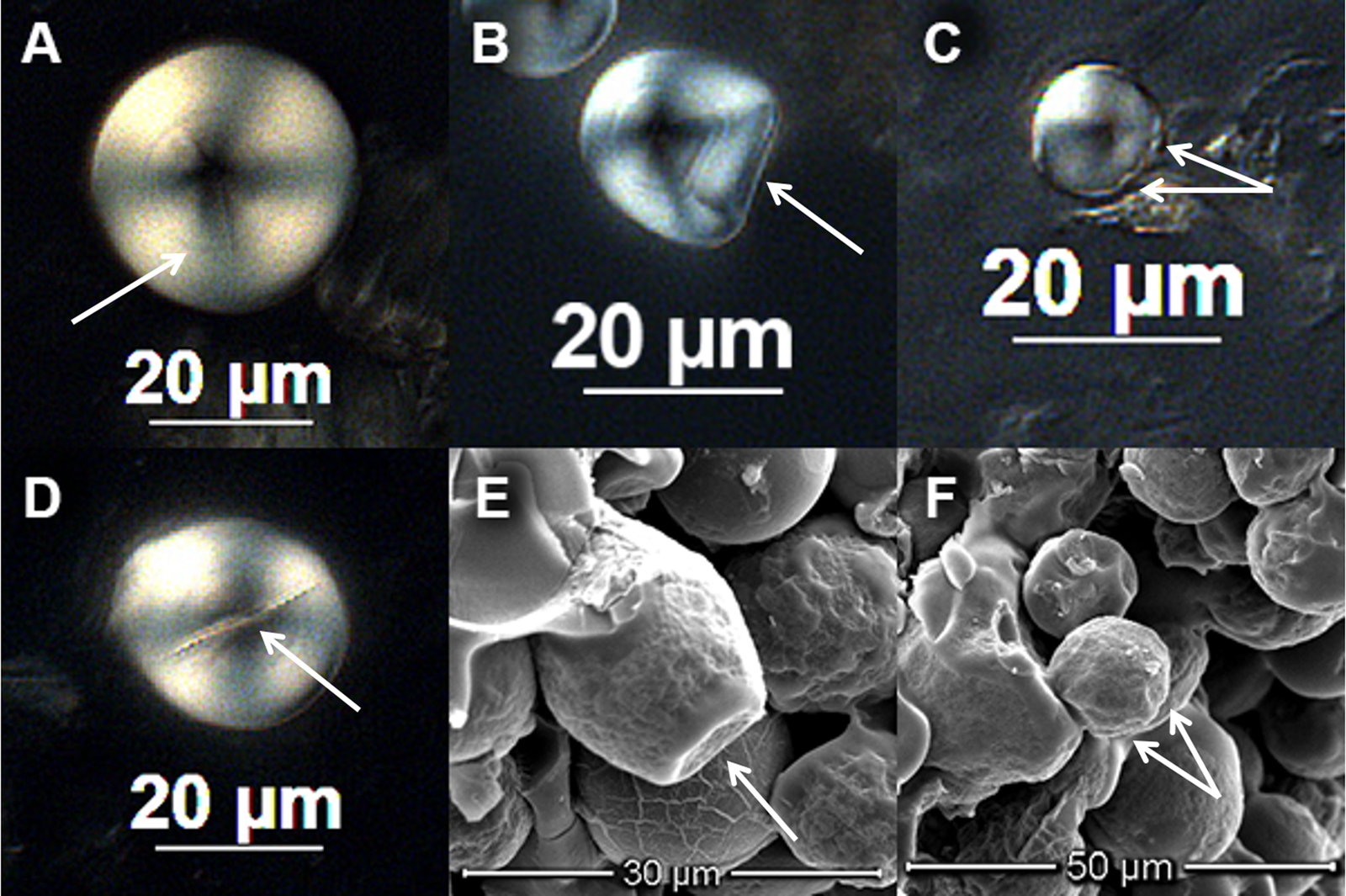
Figure 4. Morphological features of C. bulbosus. Images from C. bulbosus reference granules taken under transmitted light (differential interference contrast; DIC) and scanning electron microscope (SEM): (A) lamellae (Population 1B, DIC), (B) pressure facet (Population 2Z, DIC), (C) depressions (Population 1B, DIC), (D) fissures at the hilum (Population 1B, DIC), (E) pressure facet (SEM), (F) depressions (SEM). SEM images are from a C. bulbosus specimen from the Garrett Herbarium (NHMU). (Color online)
Fissures at the hilum (Figure 4d) are also commonly observed in many plant families and often used to identify archaeological maize granules (e.g., transverse, stellate, radial fissures; Holst et al. Reference Holst, Moreno and Piperno2007; Musaubach et al. Reference Musaubach, Plos and Pilar Babot2013; Wilks et al. Reference Wilks, Louderback and Boomgarden2021). They are also common in Apiaceae, but not necessarily diagnostic at the level of genus. Fissures and pressure facets have been readily observed on Apiaceae starch (e.g., Lomatium cous), but depressions along the margin of the granules seem unique to Cymopterus (bulbosus, in this study), thereby distinguishing it from species of Lomatium and other Apiaceae so far examined (Herzog Reference Herzog2014; Herzog and Lawlor Reference Herzog and Lawlor2016; Rankin Reference Rankin2016).
Population 1
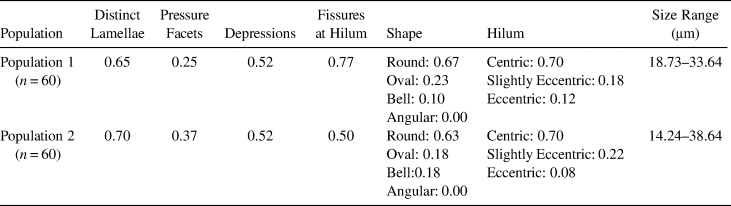
Features common for the modern reference samples collected from Population 1 include distinct lamellae (frequency [f] = 0.65), pressure facets (f = 0.25), depressions (f = 0.52), fissures occurring at the hilum (f = 0.77). The most common shape for the granules is round (f = 0.67), with few that are oval (f = 0.23) and bell shaped (f = 0.10). No angular shapes were recorded. Centric hila are the most prevalent (f = 0.70), with few that are slightly eccentric (f = 0.18) and eccentric (f = 0.12). The size range for the upper 20% of granules is 18.73–33.64 μm (Figure 4, Table 2).
Table 2. Population, Frequencies of Morphological Features, and Size Range of the Upper 20% of Reference Starch Granules from the Two C. bulbosus Populations.
Population 2
Population 2 feature frequencies were similar to Population 1. All diagnostic features were the same, with some variation in frequency. Population 2 granules exhibited distinct lamellae (f = 0.70), pressure facets (f = 0.37), depressions (f = 0.53), and fissures at the hilum (f = 0.50). Round shapes are most common (f = 0.67), with a smaller percentage of oval (f = 0.18) and bell shapes (f = 0.18). No angular shapes were documented. Centric hila are observed the most (f = 0.70), with fewer slightly eccentric (f = 0.22) and eccentric (f = 0.08). The size range for the upper 20% of granules is 14.24–38.64 μm (Figure 4, Table 2).
Ground Stone Artifacts
The tools from 48UT375 yielded 169 granules, and tools from 48SW8842 yielded 292 granules, for a total of 461 granules (Table 3). Nine out of the 10 tools produced granules assigned to Cymopterus. Using the morphological characteristics (i.e., lamellae, pressure facets, fissures, and depressions) defined by the reference granules, three archaeological granules were definitively assigned to Cymopterus, and 123 (for a total of 126) were likely or possibly Cymopterus or other Apiaceae based on possession of three or two characteristics (Figure 5, Supplemental Figure 2, Supplemental Table 1). Because we focused specifically on Cymopterus, the remaining 335 archaeological granules have yet to be identified. The three granules definitively assigned to Cymopterus (Figures 5c and 5d) were extracted from the same tool (SW8842-829; Figure 6), whereas the others came from the rest of the assemblage. Granule size for the 126 granules ranged from 8.48 μm to 38.85 μm. Reduced confidence in identification was often due to the lack of depressions diagnostic of Cymopterus.

Figure 5. Archaeological granules extracted from SW8842-1934: (A) and (B) granules likely Cymopterus or other Apiaceae based on three diagnostic characteristics; (D) and (D) granules definitively assigned to Cymopterus based on four diagnostic characteristics. (Color online)

Figure 6. Sandstone mano (SW8842-1934) that yielded starch granules assigned to Cymopterus. (Color online)
Table 3. Summary of Starch Granules Extracted from Ground Stone Tools Excavated at Sites 48UT375 and 48SW8842.
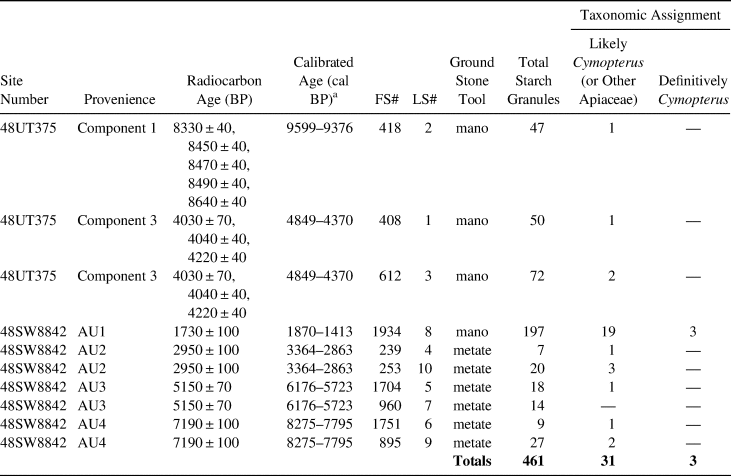
Note: Definitive assignment of three granules to Cymopterus based on the possession of four diagnostic characteristics. Another 123 granules were likely or possibly Cymopterus or other Apiaceae based on possession of fewer characteristics.
a Ages were calibrated using IntCal (Reimer et al. Reference Reimer, Bard, Bayliss, Warren Beck, Blackwell, Ramsey and Buck2013). A Bayesian model was used to obtain 95% ranges.
Discussion
Our approach to identifying archaeological starch granules from ground stone artifacts (Louderback and Pavlik Reference Louderback and Pavlik2017; Louderback et al. Reference Louderback, Herzog and Pavlik2017) began with a systematic study of C. bulbosus granules extracted from extant plant populations within the vicinity of sites 48UT375 and 48SW8842 in the Wyoming Basin. This allowed us to develop a set of statistically defined characteristics so that archaeological granules from many species could be sorted for purposes of identification. Using reference materials from two populations of C. bulbosus, those characteristics include hilum position, granule shape, distinct lamellae, fissures at the hilum, pressure facets, and depressions along the margin. All of these morphological features are commonly observed on Apiaceae granules, but only depressions along the margin were found to be a reliable feature of Cymopterus. Likewise, granule size was not an appropriate criterion for taxonomic identification because the upper 20% size range overlaps with several other plant taxa (e.g., Lomatium, Leymus, Quercus; Brown and Louderback Reference Brown and Louderback2020; Perry and Quigg Reference Perry and Quigg2011). Although it is highly probable that the granules observed on the ground stone tools are C. bulbosus, we took a more conservative approach by assigning taxonomy to archaeological granules at the genus (Cymopterus) or family (Apiaceae) levels.
A total of 461 granules were recovered from 10 ground stone tools, but less than 30% were assigned to Cymopterus or other Apiaceae. Nevertheless, these starch residues provide direct evidence that Cymopterus was collected, processed, and consumed and therefore support hypotheses about geophyte exploitation in the Wyoming Basin (Smith et al. Reference Smith, Reust and Richard2003).
The availability of predictable food resources, such as Cymopterus, may have been among the ecological drivers for an observed increase in the number of housepits in Wyoming during the middle Holocene (Smith Reference Smith2003; Smith and McNees Reference Smith and McNees2011). This proliferation of housepit construction corresponds with a period of exponential population growth (8000–5500 cal BP) and increased spatial clustering of these foraging populations across the landscape (Robinson et al. Reference Robinson, Jabran Zahid, Codding, Haas and Kelly2019; Zahid et al. Reference Zahid, Robinson and Kelly2016). Seasonally predictable geophyte resources were likely a contributing factor in the persistent reoccupation of sites (Smith Reference Smith2003; Smith and McNees Reference Smith and McNees2011). The present study concludes that these foragers were collecting and processing geophytes, in particular Cymopterus, from dense patches adjacent to inhabited sites during a period of environmental change. Underground dormancy imparts a biological resistance to environmental change such that productive populations of this nutritious taproot would have been consistently available, even during the warm and dry middle Holocene (Eckerle and Taddie Reference Eckerle, Taddie, Reust, Richard, Eckerle, Taddie, Smith and Newton2002; Smith and McNees Reference Smith and McNees2005) offering high return rates and reliable yields.
Acknowledgments
Funding for this project was provided by the June Frison Memorial Fund of the George C. Frison Institute of Archaeology and Anthropology at the University of Wyoming. We thank the University of Wyoming Archaeological Repository (UWAR) for loaning the ground stone tool collections to the Natural History Museum of Utah (NHMU), where the starch analysis was conducted. Thanks to Michelle Knoll for photographing the ground stone tools. Many thanks to our field crew who helped collect several Cymopterus bulbosus plants: Carlton Gover, Ross Hillman, Jordan Jensen, and Bruce Pavlik. Thanks to Broadbent Land and Resources, GGR Management, and Sims Sheep Company for access to 48UT375; and to Diane Sapounakis for granting access to 48SW8842. We thank the Rawlins BLM field office for its support of our work at 48SW8842. The Garrett Herbarium at NHMU provided additional reference material for Cymopterus bulbosus. We are also grateful to Bruce Pavlik and members of the University of Utah Archaeological Center (UUAC) for providing feedback on earlier versions of this article. Thank you to three anonymous reviewers for thoughful comments and to Laura Perez for translating the abstract into Spanish.
Data Availability Statement
All data (starch granule measurements, descriptions, and images) are available on request. Please contact [email protected].
Supplemental Material
For supplemental material accompanying this article, visit https://doi.org/10.1017/aaq.2021.115.
Supplemental Figure 1. Ground stone tools from 48UT375 analyzed for starch: A) UT375-408; B) UT375-418; C) UT375-612. Ground stone tools from 48SW8842 analyzed for starch: D) SW8842-239; E) SW8842-273; F) SW8842-895; G) SW8842-960; H) SW8842-1704; I) SW8842-1751; J) SW8842-1934.
Supplemental Figure 2. Images of archaeological granules assigned as likely or definitively belonging to Cymopterus. Granules 1–31 exhibit 3 diagnostic features (likely Cymopterus). Granules 32–34 exhibit 4 diagnostic features (definitively Cymopterus). Granules 1–4 are from site 48UT375 (granule 1 from FS418, 2 from FS408, and 3–4 from FS612). Granules 5–31 are from site 48SW8842 (granules 5–23 from FS 1934, 24 from FS239, 25–27 from FS253, 28 from FS1704, 29 from FS1751, 30–31 from FS895). All granules showing 4 diagnostic features (32–34) are from site 48SW8842 and FS1934
Supplemental Table 1. Inventory of starch granules from ground stone tools at sites 48UT375 and 48SW8842. Definitive taxonomic assignment of granules to Cymopterus is based on the possession of four diagnostic features. Granules that possessed three (likely Cymopterus) or two (possibly Cymopterus) features are also assigned












 Open access
Open access