


Introduction
Wright (Reference Wright, Wright, Simons and Gursky2003) noted that most species of tarsier, small nocturnal primates of the genus Tarsius, were categorized as Data Deficient on the IUCN Red List, and that ‘the first step in tarsier conservation is to change their Data Deficient status'. Wright went on to identify four high priority taxa, one of which was Tarsius sangirensis. Shekelle & Leksono (Reference Shekelle and Leksono2004) proposed a conservation strategy for the Sulawesi biogeographical region using tarsiers as flagship species. They identified 11 populations of tarsiers in the region that warranted further taxonomic investigation, and developed a biogeographical hypothesis for the region that predicted the possible existence of numerous other species. Together with the five species they recognized from the region, this meant that Sulawesi and surrounding island groups were subdivided into 16 or more biogeographical subregions of tarsier endemism. This distribution was hypothesized to have resulted from tectonic activity that brought a proto-Sulawesi archipelago together during the Miocene and Pliocene, and which had been further modified by range fragmentation during the Pleistocene. Each of these subregions could hypothetically contain an endemic tarsier taxon, many of which were suspected to be under threat of extinction. Thus, they proposed a plan to identify and name all cryptic tarsier species in these biogeographical subregions, determine their conservation status, and use these taxa as flagship species to promote conservation.
Thus, establishing the conservation status of tarsier populations is important both for tarsier conservation and for promoting biodiversity conservation throughout the biogeographical region that includes Sulawesi and the offshore island groups that share a similar biota. Here we use the geographical information system (GIS) database of the Sulawesi Ecoregional Conservation Assessment (Summers et al., Reference Summers, Gunawan, Salim, Sriyanto, Tasirin and Iskandar2005) prepared by The Nature Conservancy, in conjunction with field surveys, to determine the conservation status of the two tarsier populations thought to be most highly threatened: T. sangirensis from Sangihe Island, one of the four priority taxa identified by Wright (Reference Wright, Wright, Simons and Gursky2003), and a newly described species, Tarsius tumpara (Shekelle et al., Reference Shekelle, Groves, Merker and Supriatna2008), which had formerly been regarded as an insular population of T. sangirensis. As T. sangirensis is regarded as a priority species for conservation, it follows that a smaller, insular population formerly regarded as T. sangirensis is also a priority species. The new species, from Siau Island, was included on the 2006–2008 list of the world's top 25 most endangered primates (Mittermeier et al., Reference Mittermeier, Ratsimbazafy, Rylands, Williamson, Oates and Mbora2007).
Both Sangihe and Siau Islands are in the Sangihe Islands archipelago, a volcanic arc island chain in the Sulawesi Sea between Sulawesi and the Philippine island of Mindanao (Fig. 1). Sangihe and Siau Islands are c. 60 km apart and are separated by ocean depths > 1,000 m, far greater than the 180 m depth often used to estimate exposed land during Pleistocene glacial maxima (Fig. 1). Because of the way that island arcs form, it is most likely that each island was formed and colonized independently by plants and animals. Because of this, and the distance between the islands, it is highly unlikely that any recurrent gene flow is occurring or has ever occurred between the islands' tarsier populations. In the original description, T. sangirensis was described (Meyer, Reference Meyer1897) as an insular form from the islands of Sangihe (alternatively spelled Sangi and Sangir, and alternatively with the name Great or Greater affixed to the beginning, e.g. Greater Sangi Island) and Siau (alternatively spelled Siao). Genetic data indicate that T. sangirensis is a monophyletic clade and the sister taxon of tarsiers from northern and central Sulawesi, with a genetic distance consistent with isolation for a period of at least a few million years (Shekelle et al., Reference Shekelle2008). Given the great ocean depths between islands in the Sangihe chain and their geological history as an island arc, Brandon-Jones et al. (Reference Brandon-Jones, Eudey, Geissmann, Groves, Melnick and Morales2004) suggested that the tarsiers on Siau could be taxonomically separable from T. sangirensis, and suggested surveying for the presence of tarsiers on other islands in the chain. In March 2005 a team led by MS located tarsiers on Siau Island.
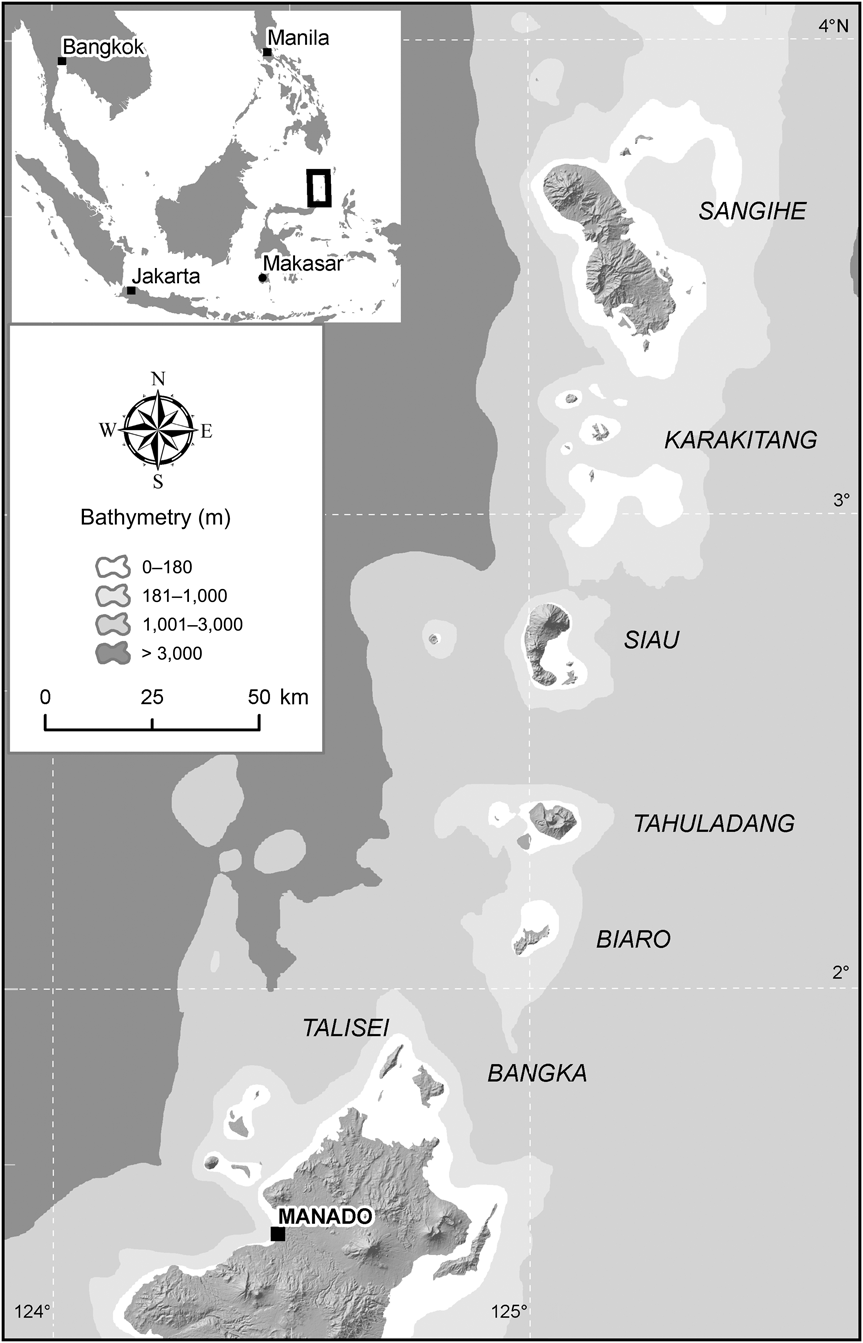
Fig. 1 The locations of Sangihe and Siau Islands within the Sangihe Island archipelago, with bathymetric contours. The rectangle on the inset indicates the location of the main figure in Indonesia.
Until recently T. sangirensis was categorized as Data Deficient on the IUCN Red List (IUCN, 2007). Supriatna et al. (Reference Supriatna, Manansang, Tumbelaka, Andayani, Indrawan and Darmawan2001), however, recommended a conservation status of Endangered (based on criteria B1a,b,c), with an estimated potential population size of 3,000 animals, based upon comparisons with other tarsiers for which density estimates exist, an extent of occurrence of 101–5,000 km2, and an area of occupancy of 11–500 km2. In this study we assess the conservation status of T. sangirensis and T. tumpara using GIS data, field surveys, and density estimates for the closest relatives of these taxa for which we have data. We draw attention to the conservation problems that tarsiers face on the Sangihe Islands, and recommend a Red List status for both tarsier species.
Methods
We treated Sangihe and Siau Islands as biogeographical subregions, each of which is a cluster of several islands separated from each other by shallow ocean (< 180 m). We estimated the maximum extent of occurrence for T. sangirensis and T. tumpara to be equal to the land area of the Sangihe and Siau Island subregions, respectively.
GIS data from the Sulawesi Ecoregional Conservation Assessment prepared by The Nature Conservancy, with Landsat images accurate to 30 m, were used to estimate the extent and quality of tarsier habitat. Habitats were classified as primary forest, secondary forest (including undisturbed and disturbed mangrove), brush (includes agriculture with brush, and brush with swamp), unsuitable (includes agriculture, no brush, swamp, open and village) and unknown (obscured by clouds).
Population densities have never been directly estimated for either T. sangirensis or T. tumpara but they have been estimated for some related species from Sulawesi. MacKinnon & MacKinnon (Reference MacKinnon and MacKinnon1980) estimated tarsier densities of Tarsius tarsier (= spectrum) to be 70 km-2 at Tangkoko Nature Reserve (North Sulawesi), mostly at sites along the coast. Gursky (Reference Gursky1997) estimated tarsier densities within the same reserve, further inland, to be 156 km-2. Tangkoko consists of mixed primary and regenerating forest, with the least disturbed habitats at higher elevations, and regenerating forests along the coast. Substantial encroachment of cultivated areas at the Reserve's edge is causing an overall reduction in tarsier habitat. Thus, the large differences between the two estimates, almost 20 years apart, could have resulted from compacting of populations as a result of habitat loss over time, methodological differences or different habitat types. In a study of Tarsius dentatus (= dianae) in habitats of varying human disturbance, Merker (Reference Merker2003) found that population density estimates in and around Lore Lindu National Park (Central Sulawesi) varied from 270 km-2 in pristine habitat to only 45 km-2 in the most heavily disturbed habitats, with 190 km-2 and 130 km-2 in areas of slight and moderate disturbance, respectively. Thus, for estimating potential population sizes of T. sangirensis and T. tumpara in the two biogeographical subregions we used the following typical tarsier densities: primary forest, 156–270 km-2 (Gursky, Reference Gursky1997; Merker, Reference Merker2003), secondary forest, 70–130 km-2 (MacKinnon & MacKinnon, Reference MacKinnon and MacKinnon1980; Merker, Reference Merker2003), and brush 45 km-2 (Merker, Reference Merker2003). For unknown habitats we produced a range of potential population estimates based on the assumption that unknown could be any habitat type except primary forest, i.e. 0–130 km-2. Using these estimates in conjunction with GIS data allows us to provide conservation assessments of the two tarsier species based upon falsifiable hypotheses for extent of occurrence, area of occupancy, habitat, habitat quality, tarsier population size estimates, human population density and geology.
Surveys for tarsiers in this study were part of a larger phylogeographical study of tarsiers with sampling at c. 100 km intervals but with sampling ad libitum within a site. The objectives of that study were to collect genetic and morphological data via trap-and-release, and acoustic data by recording the tarsiers’ dawn duet calls. Potential trapping localities were identified during daylight hours by smelling for the presence of tarsier scent marks, which are a good indicator of the presence of tarsiers. These were supplemented by interviews with local people. Dawn surveys were conducted in areas where scent marks and/or local knowledge indicated tarsier presence. Presence was confirmed either by trapping, visual sighting or the unmistakable tarsier vocalizations. Surveys of tarsier populations on Sangihe Island were conducted over 20–31 December 1995 by MS, an Indonesian student assistant, a Department of Forestry ranger, and various local assistants, and again over 11–18 July 1997 by MS, a Department of Forestry ranger, and various local assistants. Siau Island was surveyed by MS, two Department of Forestry rangers, and one para-biologist field assistant over 27 March to 1 April 2005.
Results
The Sangihe Island biogeographical subregion has an area of 585 km2, 93.5% (547 km2) of which comprises the main island of Sangihe, and the Siau biogeographical subregion has an area of 125 km2, of which Siau Island comprises 92.8% (116 km2; Fig. 2, Table 1). Estimates of habitat areas, and areas of occupancy of the two tarsier species, are given in Table 2 and illustrated in Fig. 2.
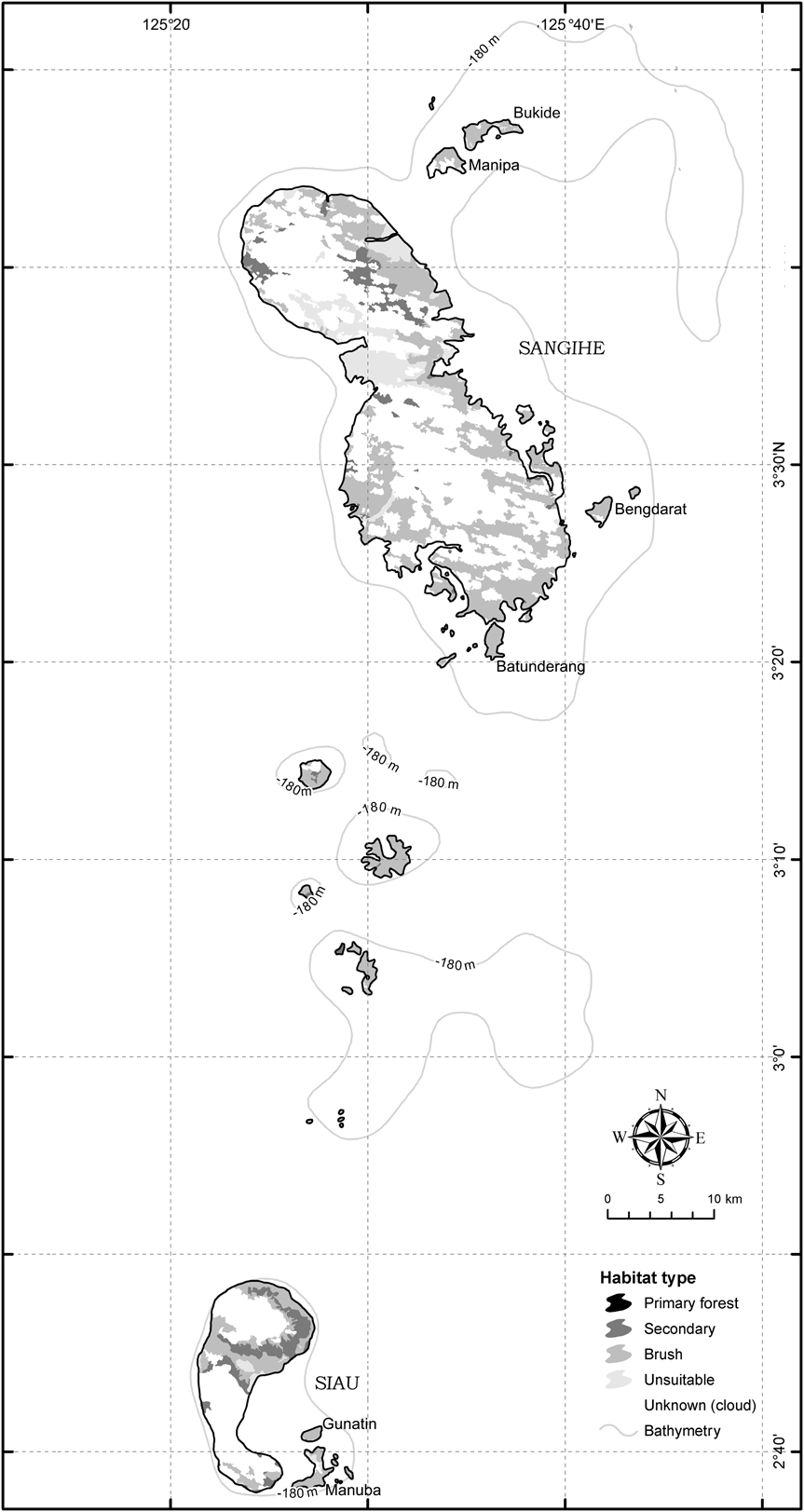
Fig. 2 GIS-based models (see text for further details) of remaining tarsier habitat on Siau and Sangihe Islands (Fig. 1).
Table 1 Areas of the main and secondary islands in the Sangihe and Siau biogeographical subregions (Figs 1–2), with human population and density, and principal geology.
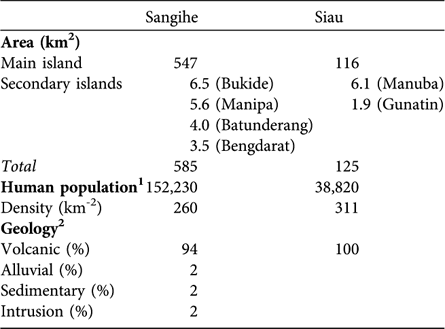
1 Sensus Penduduk Indonesia (2000)
2 Pusat Geologi Bandung (1978–1981)
Table 2 Areas of habitat types (see text for details) on Sangihe and Siau Islands (Figs 1–2), with estimates of potential tarsier population sizes derived by using typical tarsier densities for these habitats (see text for details).
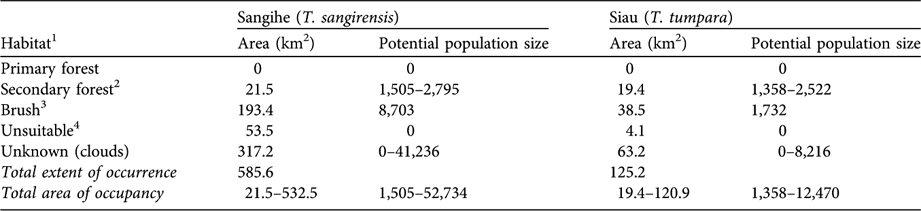
1 Determined using a Landsat Image (date of acquisition 2000), EROS Data Center, NASA
2 Includes mangrove
3 Includes agriculture with brush, and brush with swamp
4 Includes agriculture with no brush, swamp, open and village
Using the typical tarsier density estimates for habitat types in other locations, the potential population size of T. sangirensis was estimated to be 1,505–52,734. The large range is due to the high proportion of habitat classified as unknown and to lack of knowledge regarding the suitability or otherwise of brush habitat for tarsier populations. Threats facing T. sangirensis include the large active volcano, Mt Awu, a human population density of 260 people km-2 and the lack of suitable primary habitat (nor are there any wildlife conservation areas within the Sangihe biogeographical subregion). In the field surveys 12 family groups were identified. The habitat types of these locations were agroforestry, village, coconut plantation, sagu palm, secondary forest (on steep slope), scrub (on stream bank) and mangrove (Shekelle, Reference Shekelle2003). These sites, while in marginal habitat themselves, were scattered among even more degraded habitat that did not appear to support any tarsier populations at all.
Using the typical tarsier density estimates for habitat types in other locations, the potential population size of T. tumpara was estimated to be 1,358–12,470. Threats facing T. tumpara include the highly active and dangerous Mt Karengentang, a human population density of 311 people km-2 and the absence of any remaining primary forest or even notable tracts of secondary forest (see also Riley, Reference Riley2002) and, as with Sangihe, there are no wildlife conservation areas within the subregion. A single confirmed tarsier group, comprising two individuals, an adult female and a subadult male, was located during the surveys in a patch of mixed secondary forest/agroforestry habitat. On two occasions tarsiers were heard but the sites were not sufficiently distant to rule out the possibility they were the located group. In one other instance tarsier scent marks were found on a steep rocky cliff above a road but no other sign of tarsiers was detected. Except for the one confirmed tarsier group we did not detect tarsiers in marginal habitats, such as agroforestry, where tarsiers were found on Sangihe. Interviews with local people indicated that, formerly, tarsiers were commonly hunted and eaten, as many as 5–10 at a time, as part of a common activity on Sunday afternoons in which small animals are roasted on a spit, a snack food called tola-tola. Furthermore, these same people indicated that tarsiers are now absent or extremely rare in areas where they had been common as recently as 10 years ago.
Discussion
Commenting on Wright's (Reference Wright, Wright, Simons and Gursky2003) statement that it should be a priority to survey all tarsier species, Shekelle (2005) observed ‘the reality is that the number of tarsier species may well outnumber the pool of active tarsier field biologists, making it … an impractical goal until more researchers are trained and funded.' Thus, the choice was between an unreasonably long period of time until each tarsier species could be assigned a Red List threat status based on direct surveys, or developing an indirect approach that could combine existing tarsier data with remote sensing and other GIS tools. Our goal, therefore, was to prepare tarsier Red List assessments in the form of falsifiable hypotheses using existing data and GIS, supplemented where possible with direct field surveys.
The GIS data allow us to estimate that the maximum extent of occurrence for each of these populations is well below the 5,000 km2 cut-off for consideration for Endangered (criterion B1; IUCN, 2001). The extent of occurrence of T. tumpara is near, but slightly above, the 100 km2 cut-off for consideration for Critically Endangered (criterion B1). If one were to remove the part of the range of T. tumpara that forms the cone of the active volcano, however, the extent of occurrence would be < 100 km2.
The threat from volcanism for both of these populations is great. The principal volcano on Sangihe Island is Mt Awu, one of Indonesia's deadliest volcanoes, the numerous eruptions of which have caused > 8,000 human fatalities since 1711 (Global Volcanism Program, 2006a). Mt Karangetang on Siau Island is even more active, with > 40 recorded eruptions since 1674, the most recent of which was 19 August 2007, with some avalanches reportedly reaching the coast (Global Volcanism Program, 2006b). High human population density threatens both tarsier populations, and with human population density come the associated problems of continued habitat loss and declining habitat quality, as well as the threat of predation by feral cats and dogs. Frequent hunting of T. tumpara for bushmeat is reported from Siau. In both cases the populations are severely fragmented, with projected continued declines in area of occupancy, quality of habitat, number of subpopulations, and the number of mature individuals.
Our provisional conclusions are that both T. sangirensis and T. tumpara should be categorized as Endangered based on criteria B1a,bii,iii,iv,v (IUCN, 2001); i.e. extent of occurrence of 100–5000 km2, populations severely fragmented, and with a continuing projected decline in (ii) area of occupancy, (iii) area, extent and quality of habitat, (iv) number of locations or subpopulations, and (v) number of mature individuals. For several of the threat factors the problem is more acute for the Siau Island tarsier than for the Sangihe Island tarsier: namely, the extent of occurrence, area of occupancy and potential population size are all lower, the percent of volcanic terrain is higher, the volcano is more active, the human population density is 20% higher, and there is the threat of bushmeat hunting. The reports from local people on Siau Island that tarsiers are now rare or completely absent from areas where they were once plentiful could be interpreted as evidence that they should be categorized as Critically Endangered (based on criteria A1a,c,d; IUCN, 2001). Thus, over the past three tarsier generations there has been a suspected reduction in population size of > 90% based upon the direct observations by local people living in proximity to tarsiers, along with declines in the area of occupancy and quality of habitat, as well as actual levels of exploitation. The methods described herein were adopted by the IUCN Species Survival Commission Primate Specialist Group and applied to all tarsier species outside the Philippines at the Asian Primate Red List Workshop of 8–12 September 2006 in Phnom Penh, Cambodia. Our Red List categorization recommendation for T. sangirensis was added to the IUCN Red List on 6 October 2008. Because the description of T. tumpara was not published until December 2008 (Shekelle et al., Reference Shekelle, Groves, Merker and Supriatna2008) our assessment for that taxon has yet to be considered.
Although tarsiers are detected in a variety of habitats we base our Red List assessments on our estimates of potential population in primary forest (which are zero for both species) and secondary forest only. Firstly, Merker's (Reference Merker2003) evidence indicates that agricultural habitats alone cannot sustain tarsier populations and that tarsier populations in brush should be viewed as potential recolonizers. Secondly, neither our field surveys nor those of Riley (Reference Riley2002) offer any particular hope that large tracts of primary forest, or even late succession secondary forest, will be found in the unknown pixels on the satellite photographs. Thirdly, there are no wildlife conservation areas in the region and thus no tarsiers in protected areas. Whitten (Reference Whitten2006) reported that a watershed on Sangihe Island, composed of a 940 ha patch of mixed primary and old secondary forest is providing some habitat for three species of Critically Endangered birds but this appears to be the largest and best tract of habitat for wildlife on either Sangihe or Siau Islands. Finally, even if future evidence finds that brush is suitable habitat for tarsier populations, and that discoveries of suitable tarsier habitat in the unknown pixels greatly increase the area of occupancy, it is unlikely that this would change our assessments of the species' conservation status.
Relevant to the priority placed upon in situ tarsier conservation is that all ex situ conservation efforts have failed. Of the 146 known importations of tarsiers to North America and Europe (Fitch-Snyder, Reference Fitch-Snyder, Wright, Simons and Gursky2003), only one was alive at the time of writing (and that one has since perished, Shekelle & Nietsch, 2008). Many of these importations were for the specific purpose of establishing ex situ breeding programmes. At present, and for the foreseeable future, tarsier conservation is therefore entirely reliant on in situ conservation, and the necessity for this becomes more critical as taxonomic research subdivides species into cryptic taxa, with smaller population sizes, more restricted ranges, and commensurately greater conservation risks.
The outlook for conservation on the Sangihe Island chain is bleak. This fact was illustrated memorably on the cover of the first issue of Conservation Biology, which had a picture of the Cerulean paradise-flycatcher Eutrichomyias rowleyi from Sangihe Island and a commentary that questioned whether the species had gone extinct (Whitten et al., Reference Whitten, Nash and Bishop1987). Twenty years later, when it was reported that the flycatcher was not extinct but merely Critically Endangered (Whitten Reference Whitten2006), this good news served to emphasize the seriousness of the problem in the Sangihe Island chain (Shekelle et al., 2007). Further surveys have found tarsier populations on much smaller, yet still geologically isolated, islands in the chain (MS, unpubl. data). Mitigating the threats faced by these species and populations in a country noted for political and economic instability is a huge challenge (Lowe, Reference Lowe2006; Sodhi et al., Reference Sodhi, Acciaioli, Erb and Tan2007). Those unfamiliar with the local context may be surprised by what is perceived to work and what not to work, such as Whitten's (Reference Whitten2006) suggestion that the 940 ha patch of mixed primary and old secondary forest on Sangihe Island may offer better protection because it is not a legally recognized protection area of the federal government. Deep understanding of conservation in the region is crucial.
Rylands et al. (Reference Rylands, Williamson, Hoffmann and Mittermeier2008) emphasized the importance of accumulating and publishing information such as we present herein, and then moving forward. Thus a global tarsier action plan conference was conducted in Manado, North Sulawesi, Indonesia, over 2–6 November 2008. Results from this conference are still in preparation but some conclusions are clear. Firstly, taxonomic work is ongoing and there is the expectation of further reclassification, which will result in more new taxa. Secondly, indirect methods for tarsier Red List assessments, such as those we use here for T. sangirensis and T. tumpara, and direct methods, such as used by MacKinnon & MacKinnon (Reference MacKinnon and MacKinnon1980), Gursky (Reference Gursky1997) and Merker (Reference Merker2003), are complementary, and it is important to continue using both methods. Thirdly, there are encouraging reports from small-scale captive breeding programmes in habitat countries that protocols for ex situ conservation are improving, and more investment is needed in this area. Fourthly, there are currently two tarsier conservation programmes that should be emulated elsewhere: one is the tarsier sanctuary on the Philippine island of Bohol for Tarsius syrichta fraterculus run by the Philippine Tarsier Foundation; the other is tarsier-tracking activities near Tangkoko Nature Reserve, North Sulawesi, Indonesia, which help to support small-scale, community-based ecotourism and in turn improve the effectiveness of the Reserve. Sanctuaries and tarsier tracking can be developed side-by-side and, together, they can help to promote more effective in situ and ex situ conservation. Effective conservation in the Sangihe Island chain could be aided by using endemic, threatened tarsiers to promote awareness, justify conservation programmes and attract ecotourism, much as envisioned by Shekelle & Leksono (Reference Shekelle and Leksono2004). The next step is to develop a tarsier sanctuary in North Sulawesi.
Acknowledgements
This material is based on work supported by the National Science Foundation under Grant No. INT 0107277 to MS, and grants from the Margot Marsh Biodiversity Foundation and the Gibbon Foundation to MS. Sponsorship for MS in Indonesia was provided by Noviar Andayani of the Center for Biodiversity and Conservation Studies, University of Indonesia, and by the Indonesian Institute for Science. Permits for conducting research in conservation areas and for trapping tarsiers were provided by the Indonesian Department of Forestry. Anthony Rylands read a draft of this manuscript and offered helpful comments.
Biographical sketches
Myron Shekelle is a member of the IUCN SSC Primate Specialist Group and has recently helped complete conservation assessments for all species of tarsiers. His research as a primatologist, evolutionary ecologist and conservation biologist focuses on systematic sampling of wild tarsier populations using genetic, acoustic and morphological data to address questions of evolution, taxonomy and biogeography, and then relating those results to regional conservation. He recently founded the science-based conservation organization tarsier.org. Agus Salim was the Ecoregional GIS and Data Manager for The Nature Conservancy Indonesia Program during 2001–2005. His major projects include the East Kalimantan Ecoregional Planning and Sulawesi Ecoregional Conservation Assessment. He recently finished coordinating The Nature Conservancy's forest monitoring project in East Kalimantan, and now works for PT Hatfield Indonesia as Senior Geospatial Scientist with land-cover mapping projects in Kalimantan, Papua and Sumatra, and responsibility for designing the GIS enterprise system.




