


Mortality and morbidity of neonates continue to be major problems in humans and animals, and their most common cause is diarrhoea. Optimal neonatal health depends on many factors related to management and nutrition, but improvement of the immune system is required to prevent diarrhoea. IgA is the most abundant Ig isotype in mucosal secretions and provides protection against microbial antigens on mucosal surfaces( Reference Fagarasan and Honjo 1 – Reference Ertesvåg, Naderi and Blomhoff 3 ), and IgA antibodies produced by IgA antibody-secreting cells (ASC) in the intestines and mammary glands are mainly secreted as dimers( Reference Fagarasan and Honjo 1 , Reference Wheeler, Hodgkinson and Prosser 4 ). Supplemental vitamin A and β-carotene enhance the immune response of neonates( Reference Bendich and Shapiro 5 – Reference Kume and Toharmat 9 ), and supplementation of vitamin A decreases the incidence of diarrhoea and mortality in malnourished children( Reference Mora and von Andrian 10 , Reference Stephensen, Moldoveanu and Gangopadhyay 11 ). In previous studies( Reference Nishiyama, Sugimoto and Ikeda 12 , Reference Nishiyama, Yasumatsuya and Kasai 13 ), it has been observed that β-carotene supplementation in maternal mice during pregnancy and lactation increases the numbers of IgA ASC in the mammary glands and ileum of lactating mice and enhances IgA transfer from maternal milk to neonatal mice. However, very few IgA ASC are detected in the jejunum and ileum of neonatal mice at 14 d of age and most IgA in neonatal mice may be derived from milk IgA( Reference Nishiyama, Sugimoto and Ikeda 12 ). Decreased intestinal secretions of IgA in mice can be found at weaning, and the concentrations of IgA rise rapidly after weaning and reach a maximum level at 10 weeks of age( Reference Van del Heijden, Bianchi and Stok 14 ). Thus, the enhancement of mucosal IgA induction is important for maintaining a normal immune system in neonates.
Gut-associated lymphoid tissue is the largest immunological tissue in the body. Peyer's patches are the main site for the generation of IgA+ B cells, and plasmablasts differentiating from IgA+ B cells preferentially home in on the gut lamina propria through the thoracic duct and blood by the expression of homing ligands and receptors( Reference Fagarasan and Honjo 1 – Reference Ertesvåg, Naderi and Blomhoff 3 ). Chemokines are transmembrane proteins that play important roles in innate and acquired immunity( Reference Morteau, Gerard and Lu 15 – Reference Mora, Iwata and Eksteen 17 ), and chemokine ligand CCL25 is selectively expressed in the small intestine and its receptor CCR9 is expressed by almost all T cells in the small intestine and a fraction of IgA ASC( Reference Hieshima, Kawasaki and Hanamoto 18 ). The interplay of CCL25 and CCR9 is likely to have a significant role in the recruitment of developing thymocytes( Reference Wurbel, Phillipe and Nguyen 19 ), and the selective expression of CCL25 in the small bowel underlies the homing of CCR9+ intestinal memory T cells to the small bowel( Reference Papadakis, Prehn and Nelson 20 ).
The vitamin A metabolite all-trans retinoic acid (RA) plays important roles in gut immunity and RA is necessary for the imprinting of gut-homing specificity on T cells and the induction of gut-homing receptors on B cells and IgA ASC( Reference Iwata, Hirakiyama and Eshima 16 , Reference Mora, Iwata and Eksteen 17 ). Several effects of carotenoids are thought to be mediated by their metabolism to vitamin A and subsequent mediation of retinoic acid receptor (RAR) and retinoid X receptor (RXR) response pathways( Reference Rühl 7 ). RA is a highly potent activator of RAR and RXR and influences the transcription of various retinoid response genes by the activation of these receptors( Reference Rühl 7 , Reference Haq, Pfahl and Chytil 21 , Reference Takase, Suruga and Goda 22 ). RA regulates the apoptosis of T lymphocytes through interplay between RAR and RXR, but RARα and RARγ induce opposite effects during thymic selection( Reference Szondy, Reichert and Fesüs 23 ). The expressions of RAR and RXR isotypes vary greatly in different tissues, but human lymphocytes express RXRα, RARα and RARγ( Reference Ertesvåg, Naderi and Blomhoff 3 , Reference Lømo, Smeland and Ulven 24 ). Supplemental β-carotene increases the mRNA expression of IgA C-region and the numbers of IgA ASC in the ileum of lactating mice, and these effects may be mainly due to the RA-mediated immune response( Reference Nishiyama, Sugimoto and Ikeda 12 ). However, the exact mechanism of the enhancement of mucosal IgA induction in the intestines of neonates by β-carotene is still unclear, although β-carotene supplementation has been expected to enhance RA-mediated immune response in neonates( Reference Nishiyama, Sugimoto and Ikeda 12 , Reference Nishiyama, Yasumatsuya and Kasai 13 ).
In the present study, we investigated the effect of supplemental β-carotene on the concentrations of IgA, the numbers of IgA ASC and the mRNA expressions of IgA C-region, CCL25, RXRα, RARα and RARγ in the jejunum and ileum of weanling mice. The present study demonstrates that β-carotene supplementation in weanling mice is effective to enhance mucosal IgA induction in the jejunum or ileum, because it increases the concentrations of IgA, the numbers of IgA ASC and the mRNA expression of IgA C-region in the jejunum and also the numbers of IgA ASC in the ileum. These effects may be mainly due to the RA-mediated immune response, owing to the increased mRNA expressions of CCL25, RXRα, RARα and RARγ in the jejunum and of RXRα and RARγ in the ileum.
Materials and methods
Animals and diets
Male weanling ICR mice aged 21 d (n 44) were purchased from Clea Japan (Tokyo, Japan). They were housed in individual polycarbonate cages and maintained in an air-conditioned room (24 ± 2°C) under controlled lighting conditions (14 h light–10 h dark cycle). They received humane care in accordance with the ‘Regulation on Animal Experimentation at Kyoto University’ (Animal Research Committee, Kyoto University, revised 2007).
Weanling mice were randomly allocated to the control or β-carotene group at 21 d of age. Mice in the control group were fed rodent feed (Oriental Yeast) for 7 (n 8), 14 (n 8) or 21 (n 6) d, and those in the β-carotene group were fed 50 mg/kg β-carotene-supplemented rodent feed for 7 (n 8), 14 (n 8) or 21 (n 6) d. The rodent feed contained a vitamin mix, and vitamin A concentration was 385 μg retinol activity equivalent/100 g, but β-carotene was not supplemented to the control group. For the β-carotene group, β-carotene was mixed with the rodent feed at 50 mg/kg. All mice were given free access to water and feed. Body weights and feed intake of mice were measured at 07.00 hours every day.
Sample collection
Blood samples from eight mice in the control and β-carotene groups after 7 and 14 d of treatment and six mice in the control and β-carotene groups after 21 d of treatment were obtained by cardiac puncture under anaesthesia with Avertin (2,2,2-tribromoethanol; Sigma-Aldrich Chemical), and then jejunum, ileum and rectal faeces were removed after euthanasia by cervical dislocation. The samples of jejunum and ileum were immediately frozen in dry ice-cooled isopentane (2-methylbutane; Wako Pure Chemicals) for immunohistochemical analysis or frozen in liquid N2 and stored at − 80°C for IgA immunoassay and semi-quantitative RT-PCR. Blood samples were left to stand at room temperature for 1 h and then centrifuged at 3000 rpm for 15 min. The samples of serum and rectal faeces were stored at − 20°C until IgA analysis.
IgA immunoassay and immunohistochemical analysis
IgA immunoassay of serum, jejunum, ileum and faeces and immunohistochemical analysis of jejunum and ileum were carried out as described previously( Reference Nishiyama, Sugimoto and Ikeda 12 ). IgA concentrations were measured using the Mouse IgA ELISA Quantitation Kit (Bethyl Laboratories) and the ELISA Starter Accessory Package (Bethyl Laboratories) according to the manufacturer's instructions. The sections obtained from the immunohistochemical analysis were examined under an epifluorescence microscope (BX50; Olympus), and the resulting images were analysed using Image J software (National Institutes of Health). IgA-positive cells in the jejunum and ileum were counted in the lamina propria of villi in eight randomised villi from each mouse and reported as IgA ASC/unit area of the lamina propria of villi (unit = 10 000 μm2).
Semi-quantitative RT-PCR
The mRNA expressions of IgA C-region, CCL25, RXRα, RARα and RARγ in the jejunum and ileum were examined by semi-quantitative RT-PCR as described previously( Reference Nishiyama, Sugimoto and Ikeda 12 ). The relative abundance of a specific mRNA was normalised by the abundance of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA. The primer pairs and PCR conditions used for IgA C-region and GAPDH were the same as those used in the previous study( Reference Nishiyama, Sugimoto and Ikeda 12 ). The primer pairs for CCL25 were as follows: forward: 5′-CCTTCAGGTATCTGGAGAGGAGATC-3′ and reverse: 5′-CAAGATTCTTATCGCCCTCTTCA-3′. The PCR was carried out as follows: after 95°C for 7 min to denature DNA, PCR was conducted for thirty-three cycles at 94°C for 1 min, 55°C for 1 min and 72°C for 1 min, and then for one cycle at 72°C for 7 min. The primer pairs for RXRα were as follows: forward: 5′-GAGCAGCACTGAGGATATCAAGC-3′ and reverse: 5′-GGTCAGGTCTTTGCGTACTGTCC-3′. The PCR was carried out as follows: after 95°C for 7 min to denature DNA, PCR was conducted for thirty-five cycles at 94°C for 1 min, 57°C for 1 min and 72°C for 1 min, and then for one cycle at 72°C for 7 min. The primer pairs for RARα were as follows: forward: 5′-AGCACCAGCTTCCAGTCAGT-3′ and reverse: 5′-AGTGGTAGCCGGATGATTTG-3′. The PCR was carried out as follows: after 95°C for 7 min to denature DNA, PCR was conducted for thirty-seven cycles at 94°C for 1 min, 53°C for 1 min and 72°C for 1 min, and then for one cycle at 72°C for 7 min. The primer pairs for RARγ were as follows: forward: 5′-GGGCAAGTACACCACGAACT-3′ and reverse: 5′- ATCCGCAGCATTAGGATGTC-3′. The PCR was carried out as follows: after 95°C for 7 min to denature DNA, PCR was conducted for thirty-seven cycles at 94°C for 1 min, 53°C for 1 min and 72°C for 1 min, and then for one cycle at 72°C for 7 min.
Statistical analysis
Data are expressed as mean values with their standard errors. Data of body weights and feed intake were analysed by least-squares ANOVA using the general linear model procedure of Statistical Analysis Systems (SAS Institute)( 25 ). The model was as follows:
 $$\begin{eqnarray} Y _{ ijk } = \mu + T _{ i } + M _{( i ) j } + D _{ k } + TD _{ ik } + e _{ ijk }, \end{eqnarray}$$
$$\begin{eqnarray} Y _{ ijk } = \mu + T _{ i } + M _{( i ) j } + D _{ k } + TD _{ ik } + e _{ ijk }, \end{eqnarray}$$
where μ is the overall mean, T i is the effect of treatment, M (i)j is the random variable of mice nested in treatment, D k is the effect of sampling day, TD ik is the interactions and e ijk is the residuals. The general linear model procedure of Statistical Analysis Systems( 25 ) was used to analyse the effects of treatment or time on some variables in mice. Significance was declared at P< 0·05.
Results
IgA concentrations in serum and tissues
Body-weight gains and feed intake of mice during 21 d of treatment as well as those of mice during 7 and 14 d of treatment (data not shown) were similar between the groups (Fig. 1). IgA concentrations in the jejunum of the β-carotene group were significantly higher than those of the control group after 14 (P< 0·01) and 21 (P< 0·05) d of treatment, but IgA concentrations in the serum, ileum and faeces of mice after 7, 14 and 21 d of treatment were not affected by the treatment (Fig. 2). Compared with the IgA concentrations of mice after 7 d of treatment, those in the serum (P< 0·001) and faeces (P< 0·01) increased after 14 and 21 d of treatment. IgA concentrations in the jejunum and ileum increased (P< 0·001) with age.

Fig. 1 Body weights (BW) and feed intake of the control (■; n 6) and β-carotene (□; n 6) groups during 21 d of treatment.

Fig. 2 IgA concentrations (μg/g) in the (a) serum, (b) faeces, (c) jejunum and (d) ileum of the control group (■) after 7 (n 8), 14 (n 8) and 21 (n 6) d of treatment and of the β-carotene group (□) after 7 (n 8), 14 (n 8) and 21 (n 6) d of treatment. Values are means, with their standard errors represented by vertical bars. Mean value was significantly different from that of the control group: * P< 0·05, ** P< 0·01.
IgA antibody-secreting cells in tissues
The numbers of IgA ASC in the jejunum of the β-carotene group were significantly higher than those of the control group after 7 (P< 0·05), 14 (P< 0·001) and 21 (P< 0·001) d of treatment (Fig. 3), and those in the ileum of the β-carotene group were significantly higher than those of the control group after 14 (P< 0·001) and 21 (P< 0·05) d of treatment. The numbers of IgA ASC in the jejunum and ileum of mice increased (P< 0·001) with age.
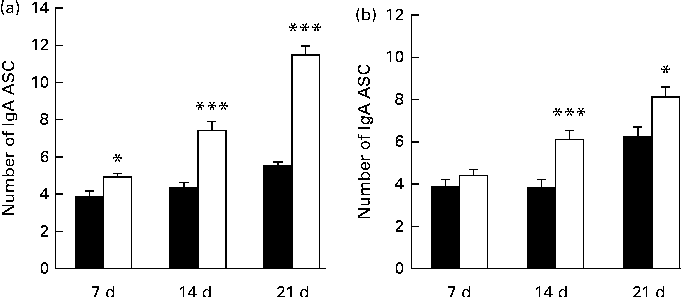
Fig. 3 Numbers of IgA antibody-secreting cells (ASC) in the (a) jejunum and (b) ileum of the control group (■) after 7 (n 8), 14 (n 8) and 21 (n 6) d of treatment and of the β-carotene group (□) after 7 (n 8), 14 (n 8) and 21 (n 6) d of treatment. Values are means, with their standard errors represented by vertical bars. The numbers of IgA ASC in the jejunum and ileum were counted in the lamina propria of villi in eight randomised villi from each mouse. Mean value was significantly different from that of the control group: * P< 0·05, *** P< 0·001.
Expression of mRNA in tissues
The mRNA expression of IgA C-region in the jejunum of the β-carotene group was significantly higher (P< 0·01) than that of the control group after 14 and 21 d of treatment, and the mRNA expression of CCL25 in the jejunum of the β-carotene group after 14 (P< 0·01) and 21 (P< 0·05) d of treatment was significantly higher than that of the control group (Table 1). The mRNA expression of RXRα in the jejunum and ileum of the β-carotene group was significantly higher (P< 0·01) than that of the control group after 21 d of treatment. The mRNA expression of RARα in the jejunum of the β-carotene group after 21 d of treatment was significantly higher (P< 0·05) than that of the control group, but that of RARα in the jejunum of the β-carotene group after 7 d of treatment was significantly lower (P< 0·01). The mRNA expression of RARγ in the jejunum of the β-carotene group after 14 and 21 d of treatment was significantly higher (P< 0·05) than that of the control group, and that of RARγ in the ileum of the β-carotene group after 21 d of treatment was significantly higher (P< 0·05). There were no significant differences in the mRNA expressions of IgA C-region, CCL25 and RARα in the ileum between the groups.
Table 1 mRNA expressions of IgA C-region, CCL25, retinoid X receptor (RXR) α, retinoic acid receptor (RAR) α and RARγ in the jejunum and ileum of the control group after 7 (n 8), 14 (n 8) and 21 (n 6) d of treatment and the β-carotene group after 7 (n 8), 14 (n 8) and 21 (n 6) d of treatment* (Mean values with their standard errors)

GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
* mRNA expression is represented relative to IgA C-region, CCL25, RXRα, RARα and RARγ mRNA expression normalised by the abundance of GAPDH mRNA.
Discussion
IgA antibodies in the intestines are specific to antigens of the intestinal microflora and act to limit the penetration of commensal intestinal bacteria through the neonatal intestinal epithelium( Reference Harris, Spoerri and Schopfer 26 ). After weaning, the intestinal mucosa of mice is exposed to a wide variety of exogenous antigens due to the sudden change in diet and the numbers of Ig-secreting cells are likely to increase ( Reference Van del Heijden, Bianchi and Stok 14 ). Mean faecal IgA concentration of mice at weaning is 3·1 μg/g, but faecal IgA concentration increases rapidly until 35 d of age and thereafter remains almost constant( Reference Takagi, Hasegawa and Sugimoto 27 ). In the present study, however, the concentrations of IgA and the numbers of IgA ASC in the jejunum and ileum of weanling mice increased rapidly with age, although concentrations in the faeces and serum increased after 14 d of treatment and then remained constant.
Supplementation of vitamin A and carotenoids affects the immune cell function during ontogenesis( Reference Garcia, Rühl and Herz 28 ), and vitamin A-depleted mice exhibit impaired IgA secretion in mucosal tissues of the small bowel( Reference Mora, Iwata and Eksteen 17 ). In the present study, supplemental β-carotene at 50 mg/kg in the diet increased the concentrations of IgA, the numbers of IgA ASC and the mRNA expression of IgA C-region in the jejunum of weanling mice after 14 and 21 d of treatment, but in the ileum, supplemental β-carotene only increased the numbers of IgA ASC. On the other hand, supplementation of β-carotene at 50 mg/kg in the diet given to maternal mice during pregnancy and lactation enhanced IgA transfer from maternal milk to neonates, owing to the increase in the numbers of IgA ASC in the mammary glands and ileum of maternal mice, but β-carotene supplementation had no effect on the numbers of IgA ASC and the mRNA expression of IgA C-region in the jejunum( Reference Nishiyama, Sugimoto and Ikeda 12 ). In the previous( Reference Nishiyama, Sugimoto and Ikeda 12 , Reference Nishiyama, Yasumatsuya and Kasai 13 ) and present studies, however, β-carotene supplementation had no effect on IgA concentrations in the serum and faeces of neonatal and weanling mice. Thus, β-carotene supplementation in weanling mice is effective to enhance mucosal IgA induction in the small intestine, owing to the increase in the numbers of IgA ASC in the jejunum and ileum, but supplemental β-carotene at 50 mg/kg in the diet may have a predominant effect on IgA concentrations in the jejunum of weanling mice.
IgA antibodies in the intestines are mainly secreted as dimers after incorporation of the J chain and association with polymeric Ig receptor (pIgR), and the transport of IgA antibodies from IgA ASC into the intestines is regulated by pIgR( Reference Fagarasan and Honjo 1 , Reference Wheeler, Hodgkinson and Prosser 4 ). In the murine small intestine, mature isolated lymphoid follicles are inductive sites for the immune response and nodular lymphoid structures can be observed in the distal small intestine( Reference Lorenz, Chaplin and McDonald 29 ). On the other hand, supplementation of fructo-oligosaccharides in mice after weaning was more effective for enhancing IgA levels in the jejunum rather than in the ileum and colon, and supplemental fructo-oligosaccharides increased not only the concentrations of IgA but also the expression of pIgR in the ileum and colon( Reference Nakamura, Nosaka and Suzuki 30 ). Additionally, coumestrol administration in maternal mice during pregnancy and lactation increased the number of IgA ASC in mammary glands, but had no effect on the concentrations of milk IgA and the mRNA expression of pIgR in mammary glands( Reference Wang, Sugimoto and Ikeda 31 ). These results indicate that the increased concentrations of IgA in the jejunum of weanling mice may be due to the increase in the mRNA expression of IgA C-region and the expression of pIgR caused by β-carotene supplementation.
Gut-associated lymphoid tissue dendritic cells rely on RA to induce IgA class switching, and RA is essential for the imprinting of gut-homing receptors on T and B cells and contributes to IgA production( Reference Iwata, Hirakiyama and Eshima 16 , Reference Mora, Iwata and Eksteen 17 ). CCL25 plays essential roles in intestinal homing of IgA ASC primarily by mediating their extravasation into intestinal lamina propria( Reference Hieshima, Kawasaki and Hanamoto 18 ). RA is important for the induction of the expression of CCR9 on activated T cells, and the blocking of RA receptors decreases the induction of gut-homing receptors( Reference Iwata, Hirakiyama and Eshima 16 ). In the present study, supplemental β-carotene increased the mRNA expressions of CCL25 and RARγ in the jejunum after 14 and 21 d of treatment and those of RXRα and RARα in the jejunum and of RXRα and RARγ in the ileum after 21 d of treatment, although it decreased the mRNA expression of RARα in the jejunum after 7 d of treatment. These results imply that the enhanced mucosal IgA induction caused by β-carotene supplementation is mainly due to the RA-mediated immune response, owing to the increased mRNA expressions of RXRα, RARα and RARγ in the jejunum and of RXRα and RARγ in the ileum, and β-carotene supplementation may influence the expression of chemokine receptor CCR9 on developing IgA ASC in the jejunum.
In conclusion, the present study suggests that supplementation of β-carotene in mice after weaning is useful for enhancing mucosal IgA induction in the intestines, owing to the increase in the concentrations of IgA, the numbers of IgA ASC and the mRNA expression of IgA C-region in the jejunum and the numbers of IgA ASC in the ileum. Foods containing animal products and pro-vitamin A carotenoids are the primary source of vitamin A, but vitamin A deficiency is associated with an increased risk of death from common childhood infections( Reference Ertesvåg, Naderi and Blomhoff 3 , Reference Mora and von Andrian 10 ). Supplementation of β-carotene-enriched formulas may improve the immune system of neonates, because β-carotene has not been detected in four of eight brands of formula preparations and plasma β-carotene of formula-fed children is significantly lower than that of breast milk-fed children( Reference Sommerburg, Meissner and Nelle 6 ). However, for children aged 1–3 years in the USA, the median vitamin A intake from foods and supplements is 1205 μg of retinol activity equivalent/d and exceeds the tolerable upper level of 600 μg/d, but does not exceed the no-observed-adverse-effect level of 6000 μg/d( Reference Allen and Haskell 32 ). Further study is needed to clarify the optimal and toxic levels of β-carotene in formulas to improve the immune system of neonates.
Acknowledgements
The present study did not receive any financial support. Experimental β-carotene was kindly provided by Sceti Company Limited (Tokyo, Japan).
The contributions of the authors are as follows: K. N., M. S., S. I. and S. K. designed the research; K. N. conducted most of the research; K. N. and S. K. contributed equally to the discussion and to the writing of the manuscript.
The authors have no conflicts of interest to declare.




