No CrossRef data available.
Published online by Cambridge University Press: 10 December 2024
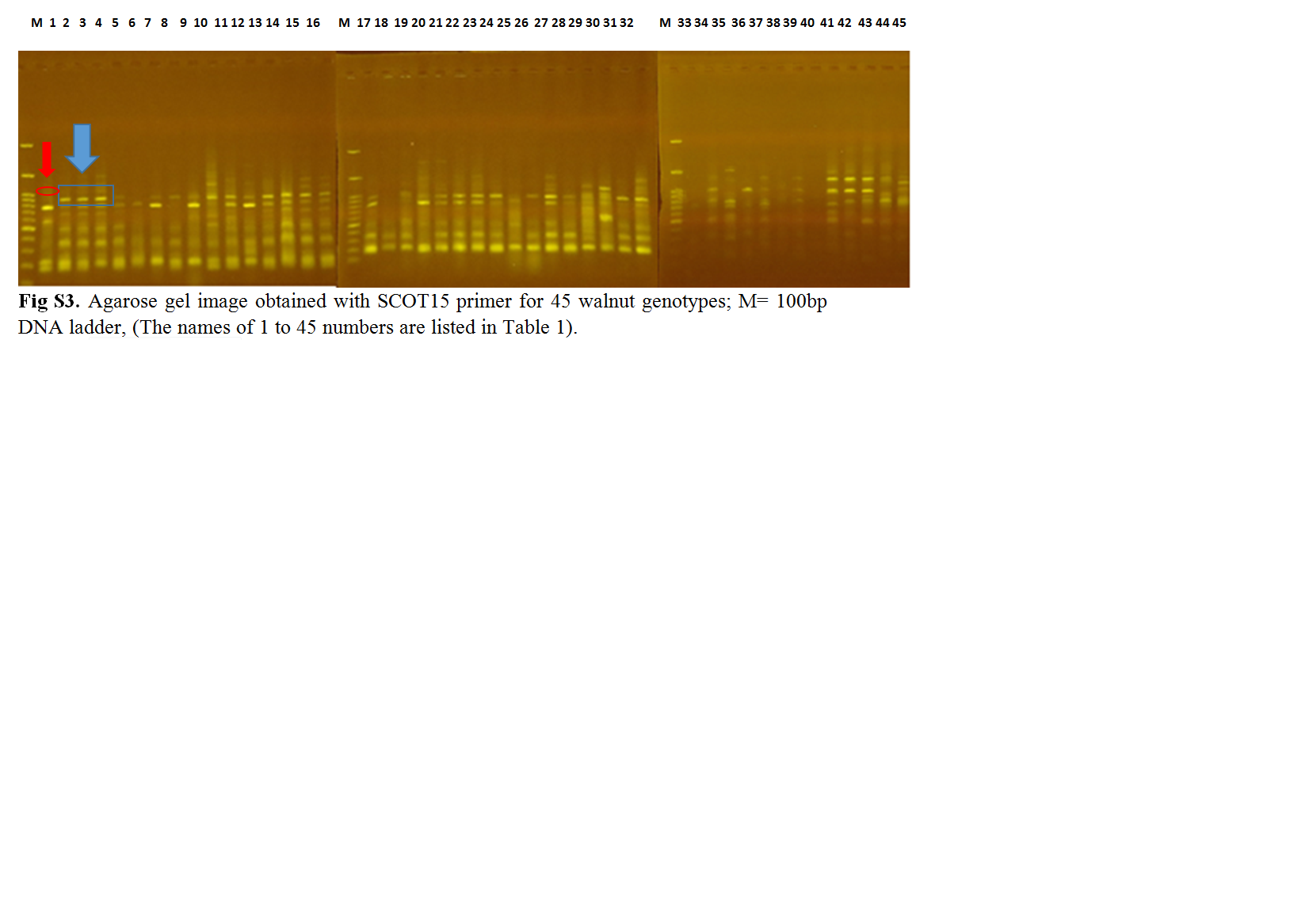
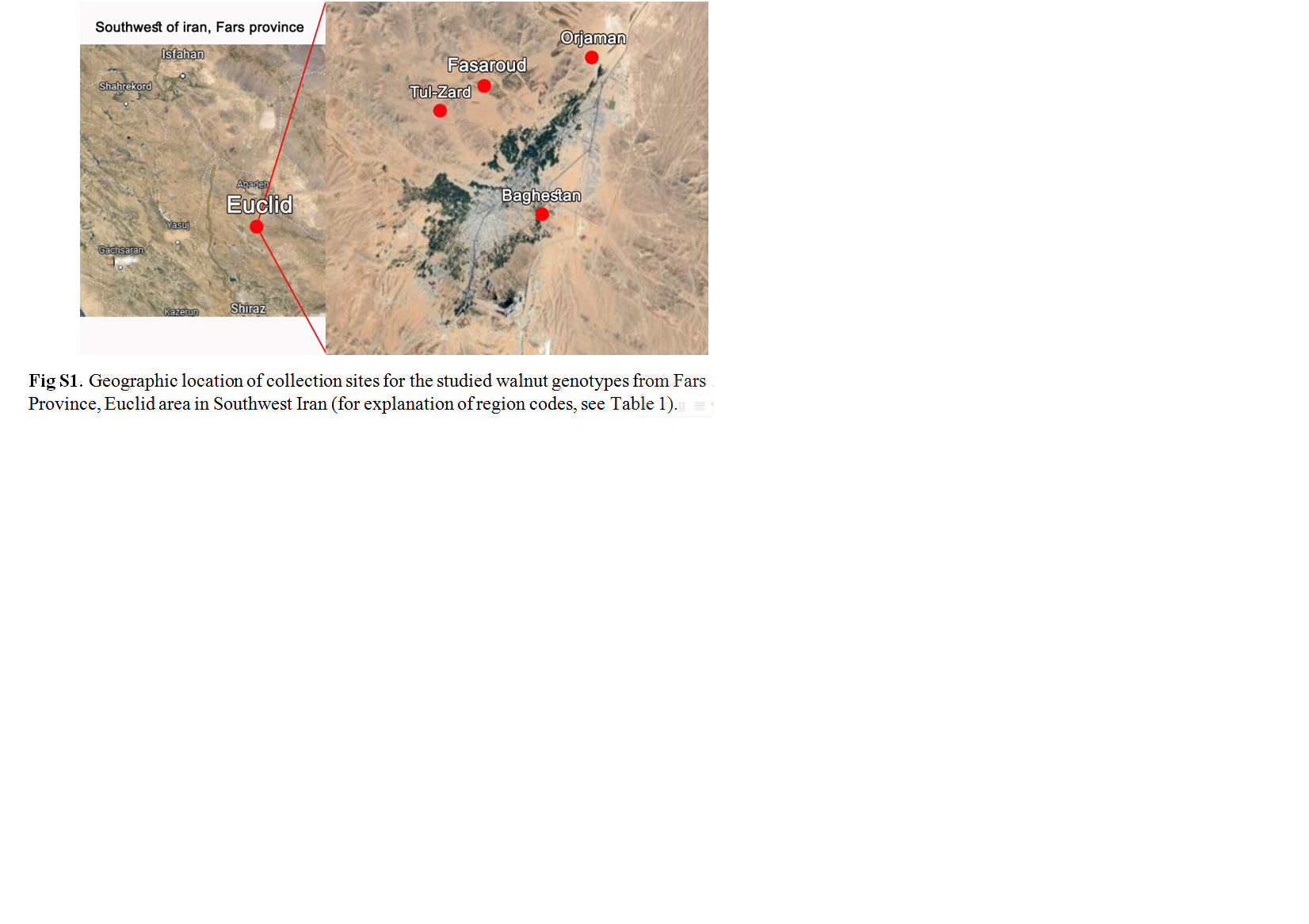
Walnut (Juglans regia L.) is one of the oldest and most economically important edible nut species due to its high nutritional value. Iran is known as one of the most important centres of the origin and diversity of walnuts worldwide. This research was carried out to determine the genetic relationships of some old walnut genotypes in Iran in order to select superior genotypes and better conserve them. The results of morphological evaluation of the fruits showed that the highest coefficients of variation were related to nut weight, kernel weight, percentage kernel, thickness and weight of the shell, and thickness and weight of the packing tissue in both years. The average morphological traits among the studied genotypes were different, such that the highest nut weights in the first and second years were 15.59 and 14.79 g and the corresponding lowest values were 7.37 and 7.79 g, respectively. The highest and lowest kernel weights were observed in the first years to be 8.09 and 3.80 g and in the second years were 6.94 and 2.65 g, respectively. The highest kernel percentages in the first and second years were 64.97 and 59.79% and the lowest percentages were 39.52 and 27.80% in the first and second years, respectively. Molecular evaluation results showed that the 16 start codon of target (SCoT) primers amplified a total of 166 bands with an average of 10.44 bands per primer. The number of polymorphic bands was changed from 2 to 19 in primers SCoT-19 and SCoT-15. The average polymorphism information content value was 0.30 and the highest amount of this index (0.36) was observed in SCoT-14 and SCoT-19 primers. The results of principal component analysis and analysis of molecular variance indicated the presence of relatively high diversity within the investigated genotypes. The amount of genetic diversity within the investigated regions was 84% and that between areas was 16%. Based on the obtained results, it was concluded that SCoT markers could provide helpful information on the genetic relationships among walnut genotypes, which can be used in future walnut breeding and conservation programmes.