



Introduction
The study of early agriculture in Central Asia and the role of the populations living along the Inner Asian Mountain Corridor in the spread of domesticated plants and animals across Eurasia have been the focus of extensive scientific debate (e.g. Frachetti et al. Reference Frachetti, Spengler, Fritz and Mar'yashev2010; Frachetti Reference Frachetti2012; Spengler et al. Reference Spengler2014; Motuzaite Matuzeviciute et al. Reference Motuzaite Matuzeviciute2015; Reference Ventresca Miller and MakarewiczVentresca Miller & Makarewicz 2019). The concept of the Inner Asian Mountain Corridor was first used by Frachetti (Reference Frachetti2012) to describe the chain of mountains (Kopet Dag, Hindu Kush, Pamir, Tien Shan, Dzungar Alatau and Western Altai) that acted as an early conduit for the movement of people, technologies, cultures, plants and animals across Eurasia. Indeed, across Central Asia, the earliest records of crop species originally domesticated in either South-west Asia (wheat, Triticum spp. and barley, Hordeum vulgare) or East Asia (millets, Panicum miliaceum and Setaria italica) are found along the Inner Asian Mountain Corridor (Frachetti et al. Reference Frachetti, Spengler, Fritz and Mar'yashev2010; Spengler et al. Reference Spengler2014; Motuzaite Matuzeviciute et al. Reference Motuzaite Matuzeviciute2015, Reference Motuzaite Matuzeviciute2017, Reference Motuzaite Matuzeviciute2018; Spengler Reference Spengler2019; Zhou et al. Reference Zhou2020). By the end of the fourth millennium BC, wheat and barley were present in the Chinese part of the Western Altai Mountains (Zhou et al. Reference Zhou2020), while the earliest broomcorn millet (Panicum miliaceum) remains in Central Asia, dated to 2560–2150 cal BC, were discovered together with wheat grains in the Dzungar Alatau mountains of south-eastern Kazakhstan (Frachetti et al. Reference Frachetti, Spengler, Fritz and Mar'yashev2010). Isotopic analysis employing incremental sampling of ovicaprid enamel bioapatite for carbon (δ13C) and oxygen (δ18O) has shown that, during the third millennium BC, domesticated animals from archaeological sites in the Dzungar Alatau range were foddered, probably with broomcorn millet (Hermes et al. Reference Hermes2019).
In western China, the earliest-known broomcorn millet grains come from the Tongtiandong site, and have been directly radiocarbon dated to 2199–1981 cal BC and 1616–1456 cal BC (Zhou et al. Reference Zhou2020). In fact, across the Xinjian region, the earliest broomcorn millets all date to the period between 2000 and 1600 BC (Tian et al. Reference Tian2021). At the Xiaohe site near Turpan, for example, broomcorn millet is directly dated to 2011–1756 and 1605–1401 cal BC (Flad et al. Reference Flad, Li, Wu and Zhao2010). The earliest-known evidence for a human C4 plant diet in western China comes from the Tianshanbeilu cemetery and is dated to 1949–1765 cal BC (Wang et al. Reference Wang2017). Notably, evidence for broomcorn millet cultivation in the third millennium BC comes from south of the Inner Asian Mountain Corridor, at the Pethpuran Teng site in northern Kashmir (Yatoo et al. Reference Yatoo2020). The later dates for broomcorn millet usage in western Xinjiang and the earlier presence of this crop along the Inner Asian Mountain Corridor and northern Kashmir raise questions concerning the directionality of early crop movements. Other questions concern how the pioneering crops of wheat, barley and broomcorn millet became integrated into local economies and moved across Central Asia: as food for herd animals (Hermes et al. Reference Hermes2019), as food “for the soul” (Frachetti Reference Frachetti, Mair and Hickman2014: 41), or as a human food source travelling alongside population movements.
A multiproxy approach to studying plant remains, as well as stable isotope ratios in human and animal skeletal material, is crucial for examining questions relating to local crop cultivation, food preparation, and direct consumption by humans and animals, as well as broader questions of crop movement across wide geographical areas. Stable isotope analysis of human and animal collagen has identified varying proportions of C4 and C3 plant consumption by humans and animals in Central Asia during the second millennium BC (Ventresca Miller et al. Reference Ventresca Miller2014; Lightfoot et al. Reference Lightfoot2015; Motuzaite Matuzeviciute et al. Reference Motuzaite Matuzeviciute2015; Ananyevskaya et al. Reference Ananyevskaya2018). For the earliest episodes of crop dispersal along the Inner Asian Mountain Corridor during the third millennium BC, however, such studies are few and limit our understanding of the extent of C3 and C4 crop use.
Situated in the middle of the Inner Asian Mountain Corridor, Kyrgyzstan is unique in that it provides a substantial number of human inhumation burials dated to the third millennium BC—that is, the period of the earliest wave of crop dispersal in the wider region. This situation offers the opportunity to apply stable isotope analysis to investigate early human, plant and animal movement through this Eurasian crossroad.
In this article, we present δ13C and δ15N stable isotope research conducted on human and animal remains ranging from the Early Bronze Age (c. 2500 BC) to the Turkic period (fifteenth century AD) in central Tien Shan, in present-day Kyrgyzstan. The stable isotope results are combined with direct radiocarbon dates and collagen peptide fingerprinting, which allows the genus affiliation of fragmented osteological material to be identified. The goal of this research is to understand how millet spread along the Inner Asian Mountain Corridor during the earliest episodes of food globalisation by analysing the intake of C4 plants into human and animal diets. We also aim to elucidate dietary change through time from the start of farming to the medieval period.
Materials and methods
The human and faunal remains analysed here were collected from sites along the Inner Asian Mountain Corridor in Kyrgyzstan, located at the highest elevation ecocline for successful cereal cultivation. Most of the human skeletal remains were sampled from the burial grounds of Aigyrzhal 1–2, Baskya 1–2 and Chechen-Bulak, at an altitude of 2000m asl, along the canyon of the river Naryn (Figure 1). Zooarchaeological identification prior to stable isotope analysis was undertaken by Dr Elina Ananyevkaya.

Figure 1. Map of sites analysed in Kyrgyzstan: 1) Aigyrzhal 1–3; 2) Kochkor burial grounds; 3) Baskya 1; 4) Baskya 2; 5) Boz Adyr; 6) Chalchyk-Bulak; 7) Chap I & II; 8) Chon-Alai; 9) Chechen-Bulak; 10) Kara-Tumshuk; 11) Keden; 12) Kok-Sai; 13) Kok-Tash; 14) Kyrk-Sheyit; 15) Kyzyl-Too; 16) Mechet at-Bashi; 17) Shyldyrak; 18) Uch-Kurbu; 19) Zhapyaryk (figure by the authors, using SRTM (NASA) for DEM, Digital Chart of the World (DCW) for water surface and Global Administrative Areas (GADM) for boundaries).
Stable isotope analysis
A total of 78 human and 84 animal samples from 17 archaeological sites were analysed (see the online supplementary material, OSM A–B). Three main groups of sites are represented: 1) Early–Late Bronze Age (2500–1200 BC, n = 46); 2) Final Bronze Age/Early Iron Age (1200–200 BC, n = 19); and 3) Turkic–medieval period (200 BC to fifteenth century AD, n = 13) (Figure 2). Information on the collagen extraction methodology, as well as measurements and calibration details, can be found in OSM B. We carried out statistical comparisons using the primary results and previously published isotope data from the region (n = 39) (de Barros Damgaard et al. Reference de Barros Damgaard2018; Narasimhan et al. Reference Narasimhan2018). For statistical analysis, we used the Mann-Whitney U test, which allows the comparison of non-parametric and unpaired data.
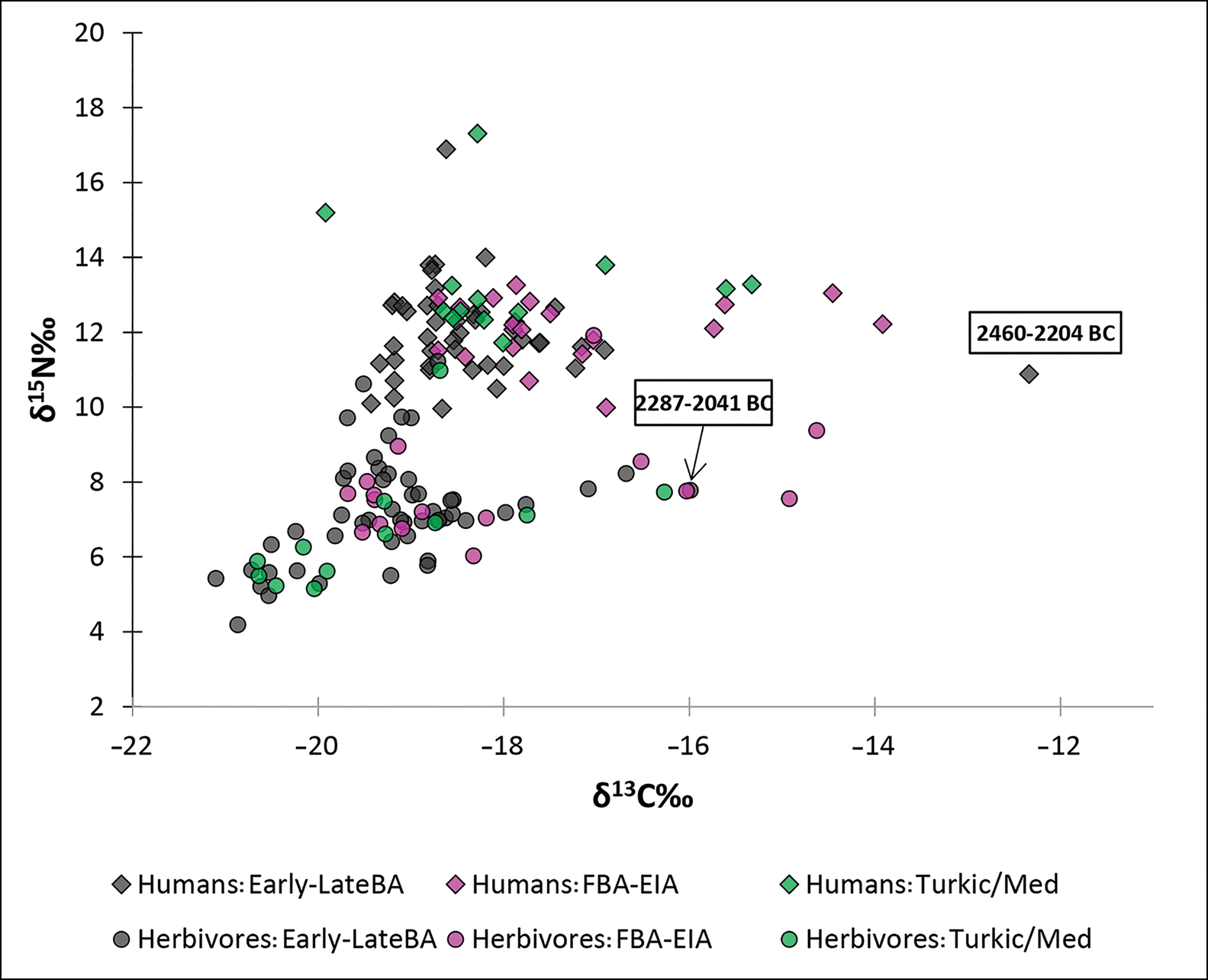
Figure 2. Scatter plot of stable isotope values of human and animal bone collagen from Kyrgyzstan (BA = Bronze Age; FBA = Final Bronze Age; EIA = Early Iron Age) (figure by the authors).
We follow previous estimations that over 20 per cent of dietary protein must originate from C4 sources for the isotopic signatures to be distinguishable from a predominately C3 diet (Hedges Reference Hedges2003), and base our interpretations on previous estimations of the δ13C cut-off value between predominately C3 and mixed C3 and C4 consumers. Predominately C3 consumers tend to demonstrate δ13C values lower than −18‰, while the δ13C value for predominately C4 consumers should be greater than −12‰ (Pearson et al. Reference Pearson2007; Lee-Thorp Reference Lee-Thorp2008).
AMS radiocarbon dating
Radiocarbon dating of human and animal bone collagen was undertaken at the 14CHRONO Centre in Belfast and at the Centre for Physical Sciences and Technology in Vilnius. In total, 13 bone collagen samples from six archaeological sites were AMS-dated. The results were calibrated in OxCal v4.4.2 using the IntCal20 calibration curve (Bronk Ramsey Reference Bronk Ramsey2009, Reference Bronk Ramsey2020; Reimer et al. Reference Reimer2020) (Figure 3).

Figure 3. OxCal v4.4.2 calibration curves of the human and faunal bone dates from Kyrgyzstan (dates calibrated in Oxcal v4.4.2 using the IntCal20 atmospheric curve; Bronk Ramsey Reference Bronk Ramsey2020; Reimer et al. Reference Reimer2020).
Collagen peptide mass fingerprinting (ZooMS)
The collagen peptide mass fingerprinting (ZooMS) method is extremely valuable for species identification of indeterminate skeletal remains, for example for distinguishing between sheep and goat species (Buckley et al. Reference Buckley2010). Eighteen skeletal fragments collected from the mid-third-millennium BC Chap II site were analysed by ZooMS, using 5–20mm bone fragments that could not be identified morphologically. The analysis was carried out at the palaeoproteomics laboratory of the University of Turin (Italy). The details of sample pre-treatment, measurement and results are available in OSM C.
Results
Stable isotope analysis
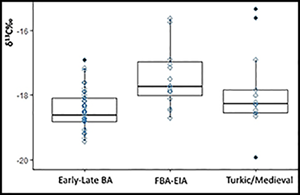
The overall human isotope values from Kyrgyzstan presented here range from −19.9‰ to −12.3‰ in δ13C and from 10.0‰ to 17.3‰ in δ15N. The individual with the highest δ13C value, of −12.3‰, comes from the Aigyrzhal-3 cemetery, Kurgan 139, and is directly dated to 2460–2204 cal BC (OSM B; Figure 3). The highest δ15N value of 17.3‰ is found in an individual from the Aigyrzhal-1 site, Kurgan 14, associated with a Turkic-period burial. Overall, the stable isotope human values for the Early–Late Bronze Age group (n = 46) range from −19.4‰ to −12.3‰, with a mean of −18.3‰ for δ13C, and from 10.0‰ to 16.9‰, with a mean of 12.0‰ for δ15N. The Final Bronze Age/Early Iron Age group (n = 19) demonstrate δ13C values from −18.7‰ to −13.9‰, with a mean of −17.3‰ and δ15N values from 10.0‰ to 13.3‰, with a mean of 12.1‰. The Turkic–medieval group (n = 13) yield δ13C values ranging from −19.9‰ to −15.3‰, with a mean of −17.7‰, while the δ15N values range between 11.7‰ and 17.3‰, with a mean value of 13.3‰ (Figure 4).

Figure 4. Box plots showing the variation of human isotope values through time (BA = Bronze Age; FBA = Final Bronze Age; EIA = Early Iron Age) (figure by the authors).
The overall isotopic values of herbivores (n = 83) range from −21.1‰ to −14.6‰ for δ13C and from 4.2‰ to 11.2‰ for δ15N. The Early–Late Bronze Age group (n = 52) values for δ13C range from –21.1‰ to –16.0‰ and from 4.2‰ to 11.2‰ for δ15N. The Final Bronze Age/Early Iron Age group (n = 16) values for δ13C range from –19.7‰ to –14.6‰ and from 6.0‰ to 11.9‰ for δ15N. The Turkic–medieval group (n = 15) values range from −21.7‰ to −16.3‰ for δ13C and from 5.2‰ to 11.0‰ for δ15N (Figure 5).

Figure 5. Box plots showing the variation of animal isotopic values through time (figure by the authors).
The earliest, least depleted δ13C herbivore value indicative of C4 plant consumption comes from sample BTC-KY-F45, dated to 2287–2092 cal BC (OSM B; see also below), from the Chap II site (Figure 1). This sample was identified through collagen peptide fingerprinting analysis to the genus level Bos (OSM C).
Statistical comparison shows that the overall stable isotope values in the Early–Late Bronze Age populations differ significantly from those of the Final Bronze Age/Early Iron Age group and medieval group for both δ13C and δ15N; values are not significantly different for either δ13C or δ15N between the Final Bronze Age/Early Iron Age and medieval groups (Table 1).
Table 1. Results of Mann-Whitney U test comparing human groups from chronologically different periods. EBA = Early Bronze Age; LBA = Late Bronze Age; FBA = Final Bronze Age; EIA = Early Iron Age.

Collagen peptide fingerprinting results
Of 18 samples measured, only one of the two duplicates of sample BTC-KY-F37 showed a poor-quality spectrum. Other samples yielded credible results, allowing identification to genus level (for spectrum images for each sample, see OSM C). Overall, 12 samples were identified as Bos: these could belong to domestic cattle, yak or any other wild bovid. Three samples were identified as either Ovis or Rupicapra sp., two samples were identified as either Ovis, Rupicapra or Ovibos sp. and one sample was identified as Capra sp.
Discussion
Diet in the highlands of Kyrgyzstan
Early Bronze Age individuals from Kyrgyzstan show a low mean δ13C value of −18.3‰, indicating a diet of predominantly C3 plants. Some individuals during this period, however, also consumed C4 plants (Figure 2). It is notable that the mean δ15N value of 11.9‰ (excluding an outlier, BTC-KY-H5: δ15N = 16.9‰) is slightly lower than the mean values from previously analysed second-millennium BC agro-pastoral communities in south-eastern Kazakhstan (12.7‰) (Motuzaite Matuzeviciute et al. Reference Motuzaite Matuzeviciute2015; Ananyevskaya et al. Reference Ananyevskaya2020a). Our results show the importance of C3 plant food—probably barley and wheat—in the population's diet during the earliest stages of the introduction of the cultigen to the central Tien Shan Highlands.
During the transition from the Final Bronze Age to the Iron Age, between 1300 and 1000 BC, there was a clear increase in C4 plant consumption as millet became a frequent dietary source for the ancient population of Kyrgyzstan. Isotopic measurements of faunal remains also show an increase in C4 plant consumption during this period (Figures 4 & 5). Stable isotope results are supported by previous archaeobotanical research that helps explain less depleted δ13C values in both human and animal bone collagen. At the sites of Uch-Kurbu (in a layer dated to 1366–1124 BC) and Chap I (dated to c. 1065–825 BC), both broomcorn and foxtail (Setaria italica) millet grains have been discovered, together with wild C4 grasses such as Echinochloa spp. and Setaria spp. (Motuzaite Matuzeviciute et al. Reference Motuzaite Matuzeviciute2018, Reference Motuzaite Matuzeviciute2020a, Reference Motuzaite Matuzeviciute, Mir-Makhamad and Tabaldiev2021). Both stable isotope and archaeobotanical results show that locally cultivated millet and ruderal C4 grasses constituted an important component of animal fodder that was probably fed to animals during the winter months at lower elevation winter camps. The increase in millet cultivation from c. 1300 BC onwards seems to be a part of a much wider phenomenon across Eurasia, as during this period broomcorn millet moved to the northern latitudes and high altitudes (Miller et al. Reference Miller, Spengler and Frachetti2016; Ananyevskaya et al. Reference Ananyevskaya2018; Filipović et al. Reference Filipović2018, Reference Filipović2020; Motuzaite Matuzeviciute Reference Motuzaite Matuzeviciute2018; Ventresca Miller & Makarewicz Reference Ventresca Miller and Makarewicz2019; Wilkin et al. Reference Wilkin2020). The rise in δ15N values during the Final Bronze Age and Early Iron Age points towards an intensification of the pastoral economy and increased consumption of animal products. Recent zooarchaeological analysis from Early Iron Age Chap I has shown that caprine animals were exploited for secondary products (wool and milk) as well as meat (Ananyevskaya et al. Reference Ananyevskaya2020b). Advances in agriculture and the intensification of field manuring may also have contributed to the overall rise in δ15N values in humans (Fraser et al. Reference Fraser2011).
There is a slight drop in C4 plant consumption and an increase in δ15N values during the Turkic–medieval period. The significant statistical difference between the Early–Late Bronze Age (mean δ15N = 12‰) and the medieval period (mean δ15N = 13.3‰) among humans contrasts with the animal nitrogen values that decrease during the medieval period (Figures 4 & 5). This could indicate greater crop consumption during the Bronze Age and a heavier reliance on animal-based products in medieval times.
C4 plant consumption from the third millennium BC
An important discovery is that the individual from a burial at Aigyrzhal-3 (Kurgan 139, sample BTC-KY-H48) was a C4 plant consumer in the third millennium BC. Dated to 2460–2204 cal BC, this individual has a δ13C value of −12.3‰. A δ13C value of −12‰ is considered to represent a predominantly C4 plant intake (Pearson et al. Reference Pearson2007; Lee-Thorp Reference Lee-Thorp2008). This identification, to our knowledge, is therefore the earliest evidence of probable millet consumption by humans along the Inner Asian Mountain Corridor of Central Asia. Given previous stable isotope analysis of δ13C values in wild and domestic faunal remains dated to the Bronze Age in neighbouring south-east Kazakhstan and the stable isotope results presented here, we can be confident that C4 plant consumption by the Kurgan 139 individual at Aigyrzhal-3 indicates direct consumption of cultivated plants—probably broomcorn millet—rather than the animals that grazed on wild C4 vegetation. Previous research has reconstructed the carbon composition of the local vegetation in the Dzungar Alatau, which ranges in δ13C value between −26‰ and −22‰, demonstrating the predominance of C3 floral taxa in the wetter mountains and drier, open steppe biomes (Hermes et al. Reference Hermes2019). Stable isotope studies conducted on modern wild plant remains show that most ecotones in Central Asia today consist of C3 plants (Ventresca-Miller et al. Reference Ventresca Miller2019), dominated by Stipa spp., Festuca spp., Carex spp., Artemesia spp., Spiraea spp. and Caragana spp. (Rachkovskaya & Bragina Reference Rachkovskaya, Bragina, Werger and van Staalduinen2012). Extensive archaeobotanical research at the third-millennium BC Chap II site has failed to identify a single grain of millet or wild Panicoid grass among the thousands of recovered wheat and barley grains and wild plant taxa recovered (Motuzaite Matuzeviciute et al. Reference Motuzaite Matuzeviciute, Hermes, Mir-Makhamad and Tabaldiev2020b). The earliest archaeobotanically identified instances of wild and domestic millets (such as Panicum miliaceum, Setaria italica, Setaria viridis and Echinochloa spp.) have only been found on sites in Kyrgyzstan dated to the Final Bronze Age/Early Iron Age (Motuzaite Matuzeviciute et al. Reference Motuzaite Matuzeviciute2018, Reference Motuzaite Matuzeviciute, Mir-Makhamad and Tabaldiev2021). At these sites, the appearance of C4 grasses probably represents weeds growing in broomcorn millet fields, but not endemic vegetation of the Tien Shan (Motuzaite Matuzeviciute et al. Reference Motuzaite Matuzeviciute, Mir-Makhamad and Tabaldiev2021).
Without macrobotanical evidence of C4 plants from third-millennium BC contexts in Kyrgyzstan, it remains unclear how the individual from Kurgan 139 at Aigyrzhal-3 was able to maintain a C4 diet while other members of the Aigyrzhal population seem to have consumed C3 plants. From a dietary perspective, this individual is unusual, and their burial was also atypical for Bronze Age Kyrgyzstan. The deceased was placed on their side in a chamber containing a stone setting (Figure 6). The vessel that was placed next to the legs has no direct parallels in Kyrgyzstan, more closely resembling the pottery styles of regions to the south or Southeast Asia. The Naryn Valley, where the Aigyrzhal site is located, is well connected with the Fergana region to the west, the Chu valley to the north, and the Kashgar and Aksu regions in China to the south and east. The diverse traditions encountered in the Aigyrzhal burials, and the metal weapons, jewellery, and pottery of various styles found in the graves, attest to wide-ranging social connections (Tabaldiev Reference Tabaldiev1996, Reference Tabaldiev2011; Moskalyev & Soltobayev Reference Moskalyev and Soltobayev2003). The region surrounding the Aigyrzhal cemetery could have constituted a hotspot of human mobility. It is likely that our C4 consumer was not local and had travelled from a millet-growing region into Kyrgyzstan. The significant effort required to construct Kurgan 139 may indicate a special status within the community. Currently, the biological sex and ancestry of this individual are unknown, but ongoing aDNA research could provide more evidence for the identity of this person and the role, if any, that they may have played in bringing the cultivation of millet to Central Asia.
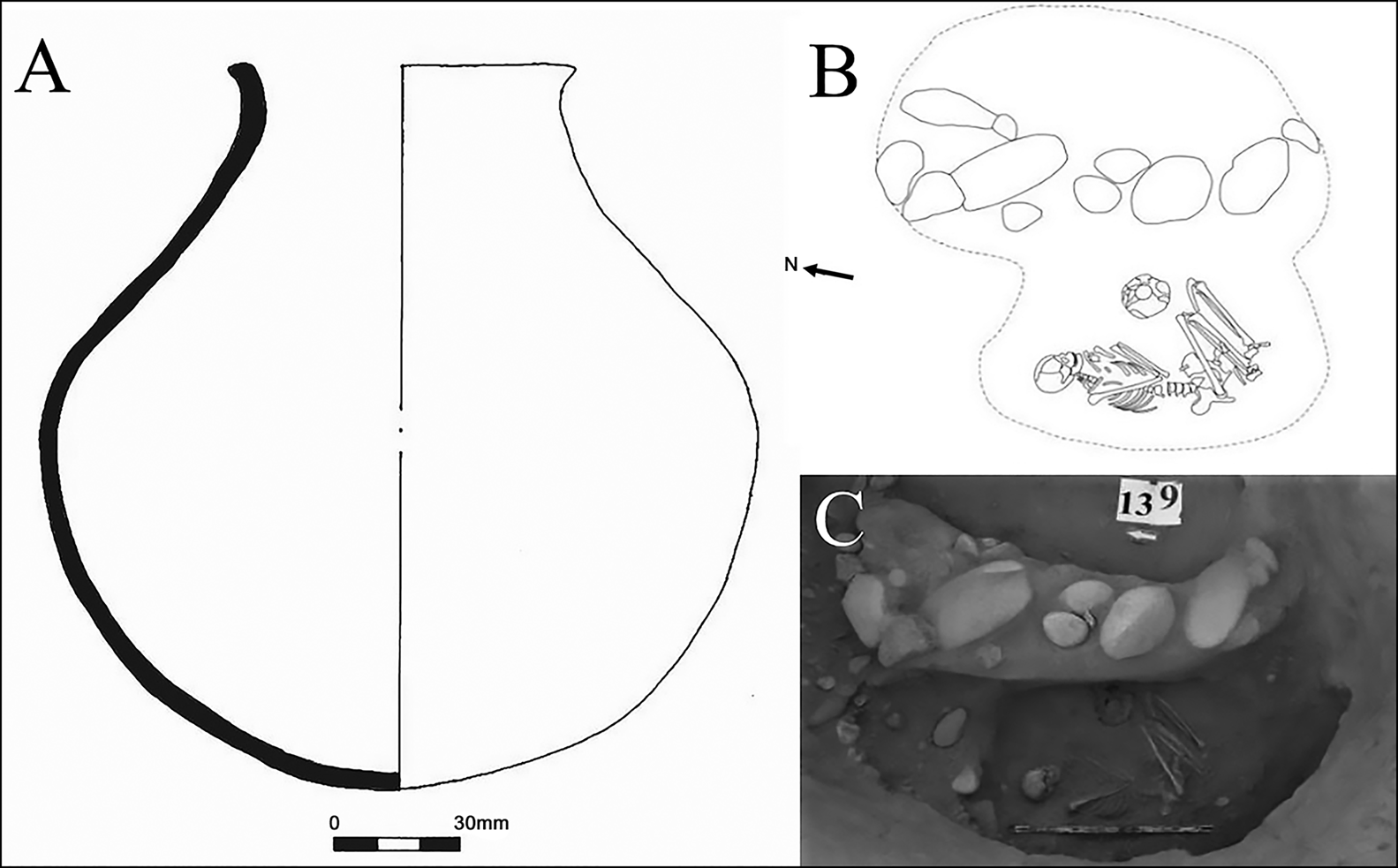
Figure 6. Pottery vessel (A) and human burial (B–C) from Aigyrzhal-3, dated to 2460–2204 cal BC (figure by the authors).
Broomcorn millet macroremains found along the Inner Asian Mountain Corridor and the south-western slopes of the Himalayas show that this crop was consumed by both humans and animals as it spread across Eurasia during the third millennium BC (Figure 7). Genetic research on Bronze Age individuals shows that this was a very dynamic period, with individuals of Iranian, South Asian and western Siberian ancestry identified among populations that brought agriculture to Central Asia (Narasimhan et al. Reference Narasimhan2018). More research is needed in regions south of Kyrgyzstan—towards the mountain valleys of the Hindu Kush, the Pamir and the Himalayas—to understand more fully human movement and crop exchange in prehistory.
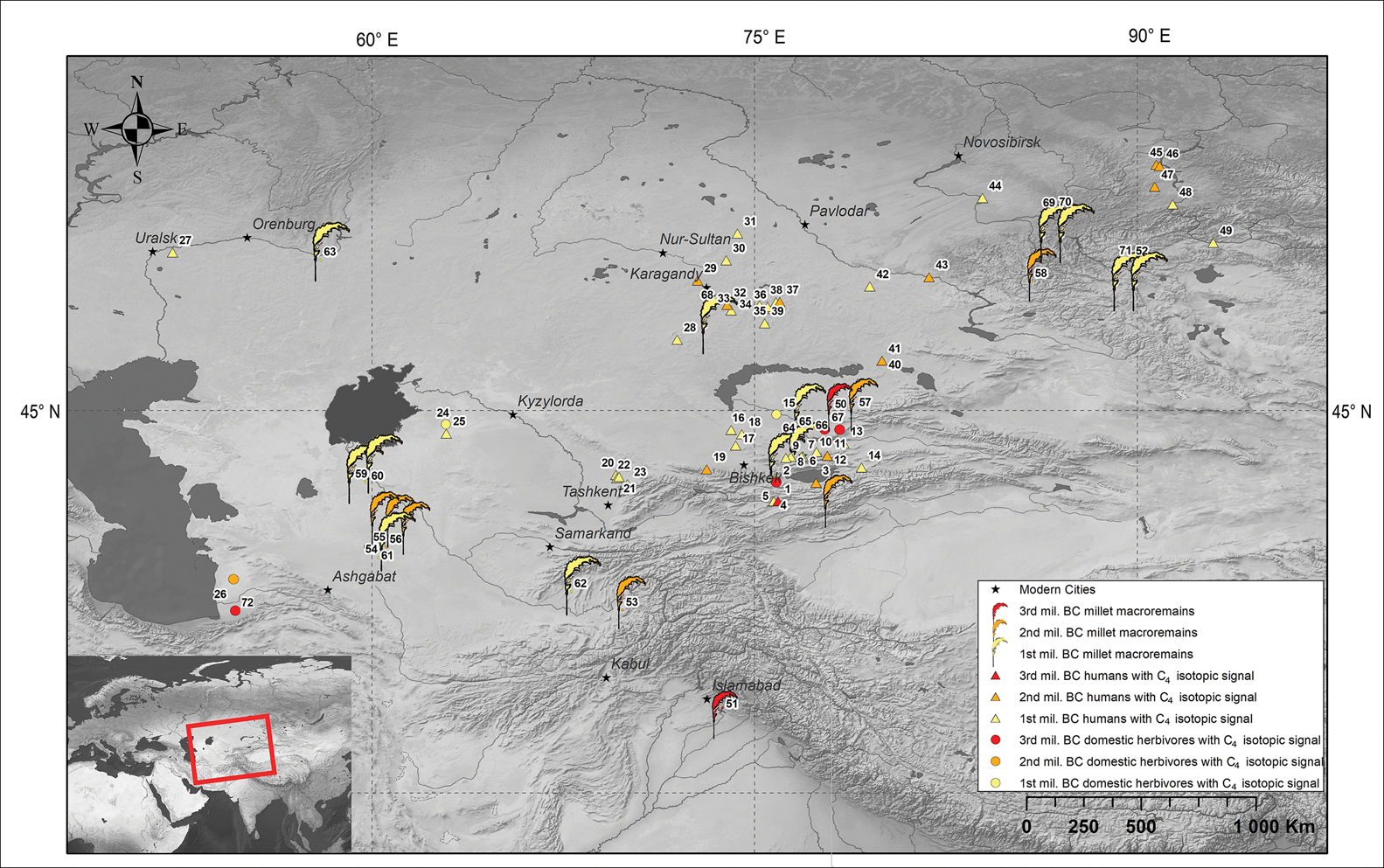
Figure 7. The distribution of broomcorn millet (Panicum miliaceum) macroremains and C4 plant consumers outside China (for detailed references, see the online supplementary material (OSM D)) (figure by the authors).
The Bos sp. (BTC-KY-F45) specimen from the Chap II site exhibit a δ13C value of −16‰ and is AMS-dated to 2460–2204 cal BC (Figure 8). The remains of this C4 plant-eating animal were found in a waste pit amongst thousands of grains of wheat and barley, and the bones of various domesticated animals, including Bos taurus. It is therefore likely that this particular example of the genus Bos is of domestic cattle. The isotope analysis of animal bone fragments from the same context shows a diet based on C3 plants, suggesting that the BTC-KY-F45 animal received different fodder and was therefore probably raised in a different place than the other animals recovered at Chap II. Dating from a similar period, C4 plant-consuming animals were identified at the sites of Dali and Begash in the Semirechye region of south-east Kazakhstan (Hermes et al. Reference Hermes2019), approximately 300km to the north-east of Chap II. The Begash site has also yielded carbonised broomcorn millet (Panicum miliaceum) remains (Frachetti et al. Reference Frachetti, Spengler, Fritz and Mar'yashev2010). A recent study shows a high level of genetic diversity among Bronze Age–Early Iron Age goats from the Inner Asian Mountain Corridor, suggesting repeated interaction, animal movement, and exchange with wide parts of Central Asia during these periods (Hermes et al. Reference Hermes2020). It is likely that the BTC-KY-F45 animal was brought from an area where millet was cultivated—probably from the lower altitude piedmont regions.

Figure 8. Peptide fingerprinting spectrum image of the BTC-KY-F45 sample, showing the taxonomic attribution of this C4 plant-eating animal (δ13C value of −16.0‰) to the genus Bos (2287–2092 cal BC). It is likely that this individual represents domestic cattle, as other bones and horns identified as domestic Bos taurus were found in the same context (figure by the authors).
Conclusions
For the first time, directly dated and isotopically analysed human and animal remains from Kyrgyzstan can be confidently interpreted as belonging to the earliest wave of farming communities in Central Asia. The results of stable isotope analysis of such remains from later periods has then allowed us to track dietary change through time.
Our research indicates that the high-altitude valleys of the Inner Asian Mountain Corridor served as conduits for the spread of broomcorn millet across Eurasia. Cultivated C4 plants were circulating among the mountainous populations of Central Asia as both human and animal food during the third millennium BC. During this period, however, C4 plants—probably broomcorn millet in the highlands of Central Tien Shan—were available only to humans and animals who had probably spent most of their lives in regions where millet was cultivated. One Early Bronze Age, C4 plant-consuming person at Aigyrzhal-3 in Kyrgyzstan may have held a special status, and was accorded burial rites and grave goods that are atypical for this period. Both stable isotope analysis and macrobotanical remains show that it was only during the Final Bronze Age and Iron Ages that millet became a staple food in Central Tien Shan.
Our results also reveal a peak in C4 plant consumption between 1300 and 800 BC, while nitrogen values among humans kept increasing across time. The increasing amounts of nitrogen within the human food chain from the Early Bronze Age to the medieval period could indicate a change from a grain-based to a meat- and dairy-based diet, which, in turn, could point to an increase in mobile pastoralism. Unlike in the Bronze Age, the medieval lifeways represent pastoral, cattle-breeding societies that resemble nomadic Central Asian populations as portrayed in ethnographic accounts.
Acknowledgements
We thank Beatrice Demarchi and Barbara Pergolizzi (University of Turin) for help with MS analyses and Aida Abrykanova for help with sample collection.
Funding statement
This project has received funding from the European Social Fund (project 09.3.3-LMT-K-712-01-0002) under grant agreement with the Research Council of Lithuania (LMTLT).
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.15184/aqy.2022.23











 Open access
Open access