



Management Implications
With hotter temperatures and more dry years predicted for California, Salsola spp. (Russian thistle, also known as tumbleweed) has the potential to become more persistent in arid and semiarid rangelands. This has important implications for management. At sites with adequate soil cover and competition with other plants, active Salsola control may not be required, as the site may naturally transition to a more desirable vegetation state. On the other hand, control may be economically prudent on sites where Salsola persists, for example, on arid or semiarid sites with bare ground, or during extended droughts, which may become more common in the future. Additionally, if a land manager finds mature, spiny Salsola plants difficult for humans, livestock, horses, and working dogs to travel through, or if skeletons are collecting against and weakening ranch fences, this may provide further impetus to incur the cost of controlling them. Our results suggest that herbicide was the only treatment that reduced Salsola cover. Where long-term heavy grazing has occurred, reducing stocking rates to moderate levels is also expected to assist with control. Salsola spp. seeds are viable for up to 3 yr, so intense management of Salsola should be employed at least over that time frame, if a manager determines the benefit of control outweighs the cost. Because Salsola populations on rangeland can fluctuate from year to year, one approach could be to begin herbicide treatment during a year when Salsola cover is lower (and thus requires less herbicide) and continue diligent control efforts for 2 to 3 yr or until the seedbank is depleted.
Introduction
Russian thistle, also known as tumbleweed (Salsola spp.), is a problematic C4 weedy annual forb that has invaded almost every state in the United States from its native habitat in southeastern Europe, central Asia, and Australia. It first arrived in the United States in South Dakota in the late 1870s (Dewey Reference Dewey1893), probably as a contaminant in flaxseed from Europe. Multiple species are commonly referred to as Russian thistle or tumbleweed, including Salsola australis R. Br., prickly Russian thistle (Salsola tragus L.), and Salsola ryanii (Hrusa & Gaskin), which is a fertile hybrid of S. australis and S. tragus. However, in North America, the taxonomy and number of species in the Salsola genus has been inconsistent over the years, and debate about its taxonomy continues (Hrusa Reference Hrusa2012; Ryan and Ayres Reference Ryan and Ayres2000).
Salsola spp. are able to thrive across habitats outside their native range due to the ability to quickly establish in resource-poor systems and at sites with disturbed soils. For example, individual Salsola plants can produce 100,000 to 200,000 seeds that are viable up to 3 yr (Chepil Reference Chepil1946; Robbins et al. Reference Robbins, Crafts and Raynor1952). They require very little soil moisture (as little as 9.1%) to germinate and can germinate and emerge from depths up to 7.5 cm below the soil surface (Wallace et al. Reference Wallace, Rhods and Frolich1968). Salsola spp. primarily occur in dry conditions and do not compete well with established plants (Dewey Reference Dewey1893; Young Reference Young1991). With more hot and dry years expected in California in the future (Pierce et al. Reference Pierce, Kalansky and Cayan2018), this species may become even more problematic.
Salsola spp. invasion can threaten rangeland ecosystems by excluding native species and enhancing fire risk (Allen Reference Allen1982; Young Reference Young1991). Salsola spp. plants can be difficult to move through once they develop spines, and can also cause problems when they become caught on fences (Dewey Reference Dewey1893; Goeden and Ricker Reference Goeden and Ricker1968). As a result, many management approaches have been developed to control the spread of Salsola, such as mechanical control, livestock grazing, biological control, and herbicide (DiTomaso et al. Reference DiTomaso, Kyser, Oneto, Wilson, Orloff, Anderson, Wright, Roncoroni, Miller, Prather and Ransom2013; Lecce et al. Reference Lecce, Paolini, Tronci, Gültekin, Di Cristina, Korotyaev, Colonnelli, Cristofaro and Smith2008; see Supplementary Table S1 for examples of effective herbicides for Salsola control). Of these, livestock grazing and herbicide applications are among the most popular, as these tools can be used across large areas of infestation in a logistically and more monetarily feasible manner than other control methods (Dewey Reference Dewey1893; McGinnies Reference McGinnies1968; Robbins et al. Reference Robbins, Crafts and Raynor1952). However, both of these approaches are associated with challenges. For example, high stocking rates of livestock can enhance Salsola biomass (Antill et al. Reference Antill, Naeth, Bork and Westhaver2012; Bleak and Plummer Reference Bleak and Plummer1954). Therefore, moderate stocking rates should be used for controlling Salsola cover. Also, frequent herbicide application can not only facilitate herbicide resistance (Barroso et al. Reference Barroso, Gourlie, Lutcher, Liu and Mallory-Smith2018) but can also cause unintentional negative effects on desired plant species, resulting in bare ground that can easily be reinvaded. One strategy to address issues associated with reinvasion after weed control is by employing an integrated pest management (IPM) approach in which multiple techniques are used in tandem (e.g., Gornish et al. Reference Gornish, Case, Valle, Bean and Moore-O’Leary2018a). Specifically, using a combined strategy of herbicide treatment followed by seeding desirable (native or nonnative) plants (Hergert et al. Reference Hergert, Mealor and Kniss2015) is an approach that is becoming more common in arid systems. This IPM approach can be successful, because seeded species serve to provide competitive pressure. Although the use of interactive management approaches to reduce Salsola dominance and increase native grasses has not been widely researched, a few studies have tested this approach. For example, McGinnies (Reference McGinnies1968) suggested that in normal rainfall years, herbicide is effective for controlling Salsola, helping to establish grass stands. However, in wetter years, McGinnies (Reference McGinnies1968) found that seeded grasses established well, even without herbicide application. Another study tested herbicide treatment and seeding (Antill et al. Reference Antill, Naeth, Bork and Westhaver2012), among other control strategies. Herbicide treatment provided excellent control. However, seeding was not successful. Fick et al. (Reference Fick, Decker, Duniway and Miller2016) were able to control Salsola with herbicide as well. They were also able to increase native perennial grass populations through seeding and the use of small barrier structures to prevent loss of litter, seed, and soil. Although seeding was successful, it did not appear to impact Salsola density. Reducing or completely limiting grazing at sites with poor range conditions was also suggested as a method for allowing grass to recover and litter to build up in order to shade out Salsola seedlings (Antill et al. Reference Antill, Naeth, Bork and Westhaver2012).
To explore the utility of using an IPM approach to reduce Salsola cover, we conducted an experiment in a grazed California grassland. Our study included the complete crossing of three weed control approaches, including grazing, herbicide application (chlorsulfuron + 2,4-D in spring, followed by glyphosate in fall and then 2,4-D the following spring), and seeding. We asked, across 5 yr, how do the singular and interactive effects of grazing, herbicide, and seeding affect (1) seeded species cover; (2) total native cover; (3) Salsola cover; and (4) total cover of other nonnative plants? We expected cattle grazing and herbicide application alone would reduce the cover of Salsola. However, we also expected the addition of seeding to enhance resistance of invasion by nonnative grasses and forbs.
Materials and Methods
The experiment was conducted on two adjacent ranches in San Benito County, CA, located predominantly on mixed dry annual grassland. The long-term average annual precipitation (October 1937 to September 2020) at nearby Pinnacles National Park is 41.9 cm (Western Regional Climate Center 2021). See Supplementary Figure S1 for annual rainfall at Pinnacles National Park from 2001 through 2020 and Supplementary Figure S2 for monthly rainfall during the years that data were collected for this project (2016 to 2020) (RAWS 2021). Both figures are based on the hydrologic year (October 1 to September 30). Experimental plots were located across areas composed of Mocho loam, 2% to 9% slopes (fine-loamy, mixed, superactive, thermic Fluventic Haploxerolls); Docas clay loam, 2% to 9% slopes (fine-silty, mixed, superactive, calcareous, thermic Typic Xerofluvents); and Sorrento silt loam, 0% to 2% slopes (fine-loamy, mixed, superactive, thermic Calcic Haploxerolls). While there is no Range Site or Ecological Site defined for two of the three soils, the Docas clay loam 2% to 9% soil is on the Dry Loamy Range Site/Ecological Site (NRCS 2021). Common nonnative plants in the area included foxtail barley (Hordeum murinum L.), ripgut brome (Bromus diandrus Roth), soft chess (Bromus hordeaceus L.), red stemmed filaree [Erodium cicutarium (L.) L’Hér. ex Aiton], and summer mustard [Hirschfeldia incana (L.) Lagr.-Foss.]. Native species included Menzies’ fiddleneck (Amsinckia sp.), California mustard [Guillenia lasiophylla (Hook. & Arn.) Greene], and clovers (Trifolium spp.) See Supplementary Table S2 for a full list of species and their native status. Both ranches graze stocker cattle during the growing season. Salsola australis and S. ryanii were found in high density (60% to 80% cover) at the sites before experimental deployment.
We deployed a nested block design with a total of nine replicate blocks across two adjacent ranches in February 2016. Six replicates were on one ranch and three replicates were on the other. Each block was split in half, with one side fenced to exclude grazing and one side left unfenced to allow cattle access to the plots. We deliberately deployed blocks in areas characterized by high Salsola density. On each side, we randomly deployed a complete cross of two treatments. One treatment was herbicide application, consisting of two levels: “no” herbicide application and “yes” herbicide application. The second treatment was seeding, consisting of three levels: no seeding, seeding with forage species, and seeding with native species. Because all treatments were crossed, each side of the block had six plots, for a total of 12 plots per block (108 plots total). Each plot was 2.5 m2 and a distance of 10 m separated the grazed and ungrazed treatments of each replicate. The farthest distance between replicates was 3.3 km in distance.
Treatment Application
Herbicide
The herbicide treatment was applied on March 22, 2016, at one ranch and April 4, 2016, at the other, using a backpack sprayer with chlorsulfuron (Telar® XP 75% w/w, Bayer, 100 Bayer Boulevard, Whippany, NJ 07981) at 0.052 kg ai ha−1 combined with 2,4-D (DMA® 4 IVM 456 g l−1, Corteva, Chestnut Run Plaza 735, Wilmington, DE 19805) at 2.13 kg ai ha−1. We used a four-nozzle boom with TeeJet® 8005 air-induction nozzles (TeeJet Technologies, 1242 Calle Bonita, Camarillo, CA 93012) at 206.8 kPa using a backpack CO2 applicator. The nozzles were spaced 29.2 cm (18.5 in.) apart. The material was applied in the equivalent of 448.8 L ha−1 (41 gal acre−1) of water. Herbicide was evenly sprayed across each herbicide treatment plot. Salsola plants were young, with soft leaves when treated. Grasses were green with seed heads. Plots were seeded (excluding unseeded controls) on November 8, 2016. On the same day as seeding, but just before putting down seed, 2% v/v (5.9 g ai l−1) of glyphosate (Roundup PowerMax®, Bayer) was sprayed only on herbicide treatment plots to limit competition for the native and forage mix seeding from any emerging plants. The glyphosate was applied with the same setup as described before. Plots that were not previously treated with chlorsulfuron + 2,4-D did not receive the glyphosate application. On March 29, 2017, 1.07 kg ai ha−1 of 2,4-D DMA was sprayed in the herbicide treatment plots to reduce competition from forbs that had already germinated.
Grazing restrictions for the herbicides used in this study were as follows. For chlorsulfuron, there are no grazing restrictions for any livestock with application rates up to 0.052 kg ai ha−1, and no exclosure requirements for any animals (Anonymous n.d.). For 2,4-D, a maximum application rate of 2.24 kg ai ha−1 is allowed in grazed areas (Anonymous 2013). For glyphosate, a maximum of 3.30 kg ai ha−1 is allowed on rangeland, with no required waiting period between application and grazing (Anonymous 2017).
Seeding
Native plots were hand seeded with perennial grasses: blue wild-rye (Elymus glaucus Buckley; 11.2 kg ha−1), California brome (Bromus carinatus Hook. & Arn.; 11.2 kg ha−1), and Nevada blue grass (Poa secunda J. Presl; 5.6 kg ha−1). Seed from all three species were mixed and seeded together for a total seeding rate of 28 kg ha−1 (Koukoura and Menke Reference Koukoura and Menke1995). Forage plots were hand seeded with Flecha tall fescue (Festuca arundinacea Schreb., a perennial cultivar) at a rate of 11.2 kg ha−1. The annual B. hordeaceus was added to the forage seeding plots in the fall of 2017 at 16.8 kg ha−1. Seeds were either procured from seed nurseries or collected by hand in the field. All seeding rates were based on bulk rates. Germination tests were not conducted.
Grazing
One half of each replicate was randomly fenced to prevent grazing using T-posts, hog wire, and cattle panels. Stocking rates during the experiment can be found in Supplementary Table S3. It was clear that cattle consumed vegetation in the grazed plots. In addition to observing cattle on our plots, the ungrazed plots clearly had taller vegetation than the grazed plots. It did not appear that grazing in the seeded plots was heavier than in the rest of the field.
Data Collection and Analysis
Each year, starting in 2016 until 2020, we collected the standard of absolute percent cover of all species in all plots (e.g., Bean et al. Reference Bean, Davy, Kyser and Gornish2021). Percent cover was visually estimated in 1% increments within a 1-m square. We allowed for overlapping canopies and thus potentially more than 100% total cover for a given plot. Sampling occurred at the end of April each year. We visually assessed percent cover of each species, bare ground, and thatch within a 1-m2 quadrat centered on the plot. Cattle dung, rocks, and sticks were considered bare ground. Salsola spp. and H. incana skeletons from prior years were considered thatch. All plant species present in the study area are listed in Supplementary Table S2.
To determine whether seeded species successfully recruited into the treated plots, we used a linear model with seeding treatment, grazing treatment, herbicide treatment, and their interactions as predictor variables and total cover by seeded species as the response variable. Year was included as a predictor variable to test for trends in seeded species cover, and initial Salsola cover (ISC) was included as a predictor variable to account for variable initial plot conditions on seeded species recruitment.
To test the effects of the herbicide, grazing, and seeding treatments and their interactions on total native species cover, Salsola cover, and total cover by nonnative species, we used linear models with herbicide treatment, grazing treatment, seeding treatment, and their interactions as predictor variables. Year and initial Salsola cover were also included as predictor variables. Total native species cover was calculated as the cumulative percent cover of each native species present in the plots. Total cover by nonnative species was calculated as the cumulative percent cover of each nonnative species present in the plots. All native and nonnative species present in the plots are listed in Supplementary Table S2. We did not use precipitation as a factor in our models, as preliminary analyses showed that both previous year total precipitation and current year total precipitation were not significant contributors to Salsola cover.
We performed model selection using the dredge function in the MuMIn package in R software v. 4.0.3 (Barton Reference Barton2020; Grueber et al. Reference Grueber, Nakagawa, Laws and Jamieson2011; R Core Team 2020). The full model for each response variable included herbicide treatment, grazing treatment, seeding treatment, and their interactions, as well as year and initial Salsola cover as predictor variables. Models were ranked using a corrected Akaike information criterion (AICc), and all models with delta AICc < 2 were considered significant (Burnham and Anderson Reference Burnham and Anderson2002). The percent cover data were not normally distributed, so all percent cover data were converted to proportion of plot covered on a scale of 0 to 1 and logit transformed before statistical analysis (Damgaard and Irvine Reference Damgaard and Irvine2019). Because statistical analyses were performed on logit-transformed data, we exponentiated model coefficients to interpret results (Damgaard and Irvine Reference Damgaard and Irvine2019).
Results and Discussion
Salsola spp. is a problematic invasive plant across natural and working landscapes. Employing an IPM approach is likely to demonstrate utility in reducing Salsola dominance and achieving other management goals such as reducing other undesirable nonnative species and increasing native or forage species. We tested the singular and interactive treatments of grazing, herbicide, and seeding to explore how these approaches might ultimately restrict Salsola across a 5-yr experiment.
The best-fit model for total cover by seeded species included year, initial Salsola cover, and seeding treatment (F(4, 535) = 35.42, P < 0.001, adjusted R2 = 0.2034). Year had an overall positive effect on total cover by seeded species (P < 0.001). Mean total cover by seeded species increased by 5.6% from 2016 to 2017, 2.8% from 2017 to 2018, and 3.19% from 2018 to 2019, and declined by 3.26% from 2019 to 2020 (Figure 1). Initial Salsola cover had a negative effect on total seeded species cover (reduced by 1%, P < 0.001; Figure 1). Plots seeded with native species had 18% less total seeded species cover (P < 0.01) and unseeded plots had 28% less total seeded species cover than plots seeded with forage species (P < 0.001; Figure 1). Percent cover of each seeded species is listed in Supplementary Table S4. One seeded species (B. hordeaceus) was already present at the study site, so it was present in the plots before seeding.

Figure 1. Mean ± SE cumulative percent cover by seeded species through time across seeding treatments; untransformed data are shown. Individual percent cover of each seeded species is listed in Supplementary Table S4. Asterisks denote significant differences among treatments within year (P < 0.05).
Many studies suggest that including seeding in an invasive plant control program can enhance outcomes (e.g., Farrell and Gornish Reference Farrell and Gornish2019). In our study, the native grasses E. glaucus and B. carinatus were seeded because they are fairly robust and may compete with Salsola for space (Seabloom et al. Reference Seabloom, Harpole, Reichman and Tilman2003). Another native grass, P. secunda, was added to the mix because although it is a much smaller-statured grass, it may establish better and recruit more successfully long term than the other two native species in the dry local environment (R O’Dell, personal communication). Festuca arundinacea was chosen as the forage treatment because it is robust and may compete well with Salsola, considering its successful establishment and persistence in long-term studies (Davy et al. Reference Davy, Dykier, Turri and Gornish2017). Average rainfall at the Davy et al. (Reference Davy, Dykier, Turri and Gornish2017) study sites was higher than at our sites, 57.9 cm compared with 41.9 cm. However, 2 of the 5 yr during their study had rainfall below 41.9 cm, making F. arundinacea a reasonable choice for our experiment. Bromus hordeaceus was chosen because it reseeds successfully on ranches in the local area. It was the most successful of the seeded species, increasing from 1.6% cover in 2016 to 19.3 % cover in 2019 and 16.4% cover in 2020.
For total nonnative cover, two models had significant support. The best model (F(4, 535) = 105.9, P < 0.001, adjusted R2 = 0.4378) included year, grazing treatment, herbicide treatment, and a grazing*herbicide treatment interaction (Table 1). Herbicide treatment did not have a significant effect on total nonnative cover (P = 0.48), and nonnative cover increased by 26% per year (P < 0.001; Figure 2). Nonnative cover was 27% lower overall in grazed plots compared with ungrazed plots (P < 0.001) and 31% higher in grazed plots with no herbicide treatment than in grazed plots treated with herbicide (P < 0.001; Figure 2). The next best model also included initial Salsola cover (Table 1).
Table 1. Ranked models for total nonnative species cover.
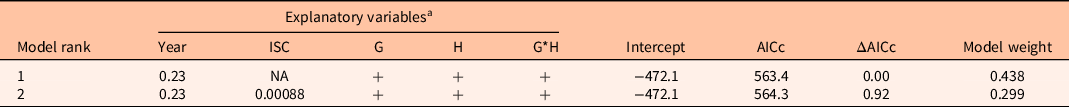
a Parameter estimates are included for continuous variables (Year, ISC). “+” means the variable is included in the model; “NA” means the variable was not included in the model. ISC, initial Salsola spp. cover; G, grazing treatment; H, herbicide treatment; G*H, interaction between grazing and herbicide treatments; AICc, corrected Akaike information criterion.
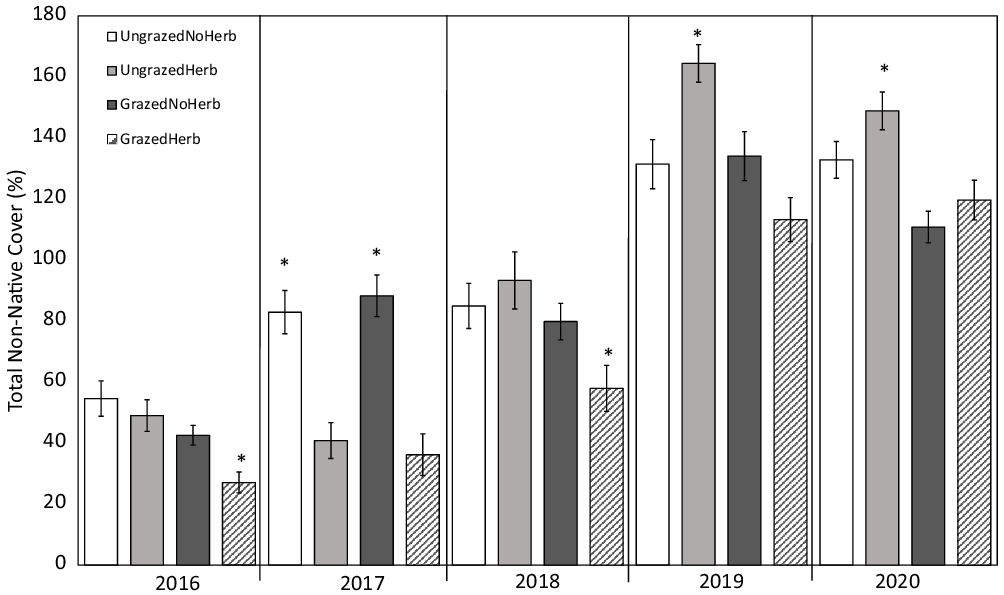
Figure 2. Mean ± SE total cumulative percent cover by nonnative species through time across grazing and herbicide treatments; untransformed data are shown. Individual percent cover of each nonnative species is listed in Supplementary Table S5. Asterisks denote significant differences among treatments within year (P < 0.05).
For total native cover, six models had significant support (Table 2). The best model (F(5, 534) = 13.33, P < 0.001, adjusted R2 = 0.10) included year, grazing treatment, herbicide treatment, and seeding treatment (Table 2). Total native species cover increased by 5% per year (P < 0.001). Grazed plots had 9% lower native cover than ungrazed plots (P < 0.001). Plots with no herbicide application had 14% higher native species cover than plots with herbicide (P < 0.001). This may be a result of overall higher cover in plots without the herbicide treatment. Native species cover was 7% lower in unseeded plots than in plots seeded with the forage species mix (P = 0.056). Native species cover was 7% higher in plots seeded with a native species mix than in plots seeded with the forage species mix (P = 0.039). The remaining models are listed in Table 2.
Table 2. Ranked models for total native species cover.
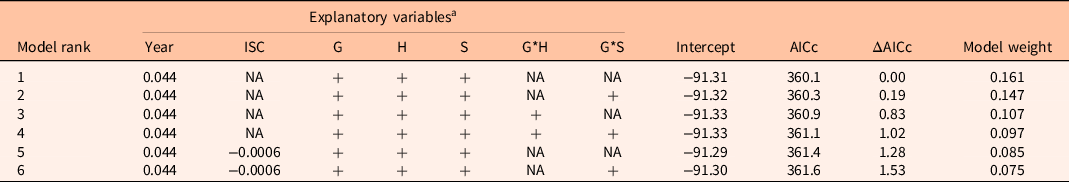
a Parameter estimates are included for continuous variables (Year, ISC). “+” means the variable is included in the model; “NA” means variable was not included in the model. ISC, initial Salsola spp. cover; G, grazing treatment; H, herbicide treatment; S, seeding treatment; G*H, interaction between grazing and herbicide treatments; G*S, interaction between grazing and seeding treatments; AICc, corrected Akaike information criterion.
For total Salsola cover, two models were supported. The best model (F(3, 536) = 94.67, P < 0.001, adjusted R2 = 0.3427) included year, initial Salsola cover, and herbicide treatment (Table 3). Year had an overall negative effect on total Salsola cover (P < 0.001). Mean Salsola cover decreased by 12.81% from 2016 to 2017, 0.81% from 2017 to 2018, 0.76% from 2018 to 2019, and 0.03% from 2019 to 2020 (Figure 3). Within each year, total Salsola cover was 26% higher in plots with no herbicide application than in plots with herbicide application (P < 0.001; Figure 3). Initial Salsola cover had a statistically significant positive effect on total Salsola cover (P < 0.001), but this effect was small (total Salsola cover increased by 0.5%) and likely not biologically significant. The next-best model (adjusted R2 = 0.259) included grazing treatment in addition to all of the explanatory variables in the best model (Table 3).
Table 3. Ranked models for total Salsola spp. cover.

a Parameter estimates are included for continuous variables (Year, ISC). “+” means the variable was included in the model; “NA” means variable was not included in the model. ISC, initial Salsola spp. cover; G, grazing treatment; H, herbicide treatment; AICc, corrected Akaike information criterion.
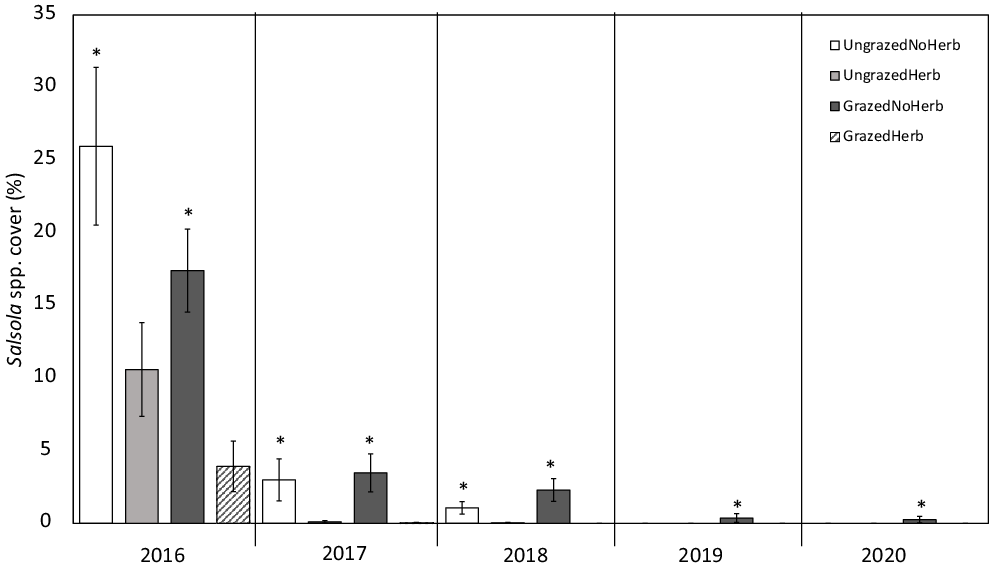
Figure 3. Mean ± SE total percent cover of Salsola spp. through time across grazing and herbicide treatments; untransformed data are shown. Asterisks denote significant differences among treatments within year (P < 0.05).
The herbicide treatment was not repeated over multiple years, potentially limiting the ability to test interaction between herbicide and other treatments. However, due to legacy effects associated with the herbicide, we would expect there to be differences in the spray+graze treatment, for example, compared with the grazing treatment in year 2 to year 5. To see a more a pronounced effect of herbicide+grazing interactions, spraying would need to be conducted multiple years in a row. Many managers want to reduce their reliance on herbicide, so a comprehensive weed management approach often includes a single year of herbicide spray followed by successive years of burning or grazing.
Our results were similar to other studies that have found that seeding of native species overall is not successful in reducing Salsola invasion (Antill et al. Reference Antill, Naeth, Bork and Westhaver2012). This might be due to the relatively low establishment of seeded native species (Figure 1). Native species can be difficult to establish by seed in arid rangeland habitat (George and Davy Reference George and Davy2020; Kildisheva et al. Reference Kildisheva, Erickson, Merritt and Dixon2016). Also, we broadcast seeded our plots. Seeding may have been more successful if seed had been covered by soil. Uncovered seed is less likely to succeed at sites with lower rainfall (George and Davy Reference George and Davy2020) and due to predation by rodents and birds (Nelson et al. Reference Nelson, Wilson and Goebel1970). Only the forage species mix resulted in any appreciable difference in seeded species cover compared with unseeded controls. Because forage species tend to demonstrate high establishment success, identifying species from this group that overlap with Salsola in resource use might provide utility for control.
Grazing is a feasible large-scale option for weed management and native plant enhancement by producers on dryland systems (Gornish et al. Reference Gornish, Eastburn, Oneto and Roche2018b; Hart and Ashby Reference Hart and Ashby1998). Grazing demonstrated high utility for controlling nonnative plant cover in general (Figure 2), particularly when paired with herbicide application. However, grazing alone did not have any effect on Salsola cover (Figure 3). This was unexpected, because while cattle avoid mature plants, young, tender Salsola individuals are palatable (Dittberner and Olson Reference Dittberner and Olson1983; USDA 1937) and nutritious (Hageman et al. Reference Hageman, Fowler, Suzukida, Salas and Lecaptain1988). In fact, they contain almost 19% crude protein, which is similar to early cut alfalfa (Medicago sativa L.) (Hageman et al. Reference Hageman, Fowler, Suzukida, Salas and Lecaptain1988). It is possible that the dramatic decrease in Salsola through time obfuscated any grazing effects.
Because grazing is effective at reducing many common rangeland weeds and enhancing natives in many situations (Bartolome et al. Reference Bartolome, Allen-Diaz, Barry, Ford, Hammond, Hopkinson, Ratclif, Spiegal and White2014; DiTomaso et al. Reference DiTomaso, Kyser, Oneto, Wilson, Orloff, Anderson, Wright, Roncoroni, Miller, Prather and Ransom2013), we advocate for employing grazing within a larger IPM strategy for Salsola control. This might be particularly important in situations where Salsola invasion includes multiple Salsola species, because each species responds dissimilarly to control approaches (Bruckart et al. Reference Bruckart, Cavin, Vajna, Schwarczinger and Ryan2004; Sobhian et al. Reference Sobhian, Ryan, Khamraev, Pitcairn and Bell2003). If grazing is to be used, stocking rates should be moderate, as Salsola can become a problem in situations where high levels of grazing creates bare ground and limits establishment of other vegetation that could create competitive situations (Antill et al. Reference Antill, Naeth, Bork and Westhaver2012).
Herbicide treatment has been shown to be successful in reducing Salsola cover (Antill et al. Reference Antill, Naeth, Bork and Westhaver2012; Supplementary Table S1), although the magnitude of control can vary depending on herbicide and applicator type (Young et al. Reference Young, Yenish, Launchbaugh, McGrew and Alldredge2008). Herbicide is an important weed management tool on rangelands where large areas covered by Salsola can be treated by ground or aerial treatment. Herbicides are particularly useful for Salsola, because both established plant cover and seed germination can be reduced (Young and Whitesides Reference Young and Whitesides1987). Specifically, Young and Whitesides (Reference Young and Whitesides1987) achieved 73% control of flowering Salsola plants treated with chlorsulfuron, glyphosate plus chlorsulfuron, paraquat, and bromoxynil plus metribuzin in crop fields after harvesting dryland small-grains. These same herbicides, in addition to others tested in their experiment, also reduced Salsola seed germination. Herbicide treatment (e.g., 2,4-D) may be most effective when conducted when Salsola plants are immature (McGinnies Reference McGinnies1968). Clearly, herbicide is an effective approach for Salsola management. However, application techniques can modify control success and should be considered before use. One example is widespread herbicide resistance reported for Salsola (e.g., Kumar et al. Reference Kumar, Spring, Jha, Lyon and Burke2017; Saari et al. Reference Saari, Cotterman, Smith and Primiani1992). Chlorsulfuron is an acetolactate synthase inhibitor (2(B) category, Herbicide Resistance Action Committee, https://hracglobal.com/tools/hrac-moa-2020-revision-description-and-master-herbicide-list) and has a different mode of action than 2,4-D (4(O)); alternating the use of these two herbicides could reduce the risk of herbicide resistance developing. Surfactant type has also been shown to affect control success (Harbour et al. Reference Harbour, Messersmith and Ramsdale2003) and should be carefully identified with the assistance of experts before application. Finally, to limit unintended herbicide exposure on desired adjacent plants, season of application should be limited to when Salsola is actively growing and other species are dormant (e.g., Lyon et al. Reference Lyon, Barroso, Thorne, Gourlie and Lutcher2021).
In addition to the treatments we deployed, precipitation may have influenced Salsola cover. While data from this study do not demonstrate a relationship between precipitation and Salsola cover, we hypothesize this may have been a factor in reduced Salsola cover over time. Dewey (Reference Dewey1893) suggested that it is not the rain itself, but the increased competition with other plants in wet years that constrains Salsola populations. Young (Reference Young1991) also indicated that Salsola does not compete well in the presence of other established plants. At our site in 2015, ranchers were concerned that Salsola was growing in areas they had not seen it before and population density at sites where it previously grew had increased. This was the third year of a severe drought during which rainfall was approximately 21.3 cm in 2012 to 2013, 15.8 cm in 2013 to 2014, and 23.1 cm in 2014 to 2015 (Supplementary Figure S1). Average annual rainfall from 2001 to 2020 was 28.4 cm. It may be that Salsola was able to expand due to low rainfall and minimal competition from other plants. Rainfall in 2015 to 2016, which was our first year of data collection, was 36.5 cm, which is above average for the 2001 to 2020 time period, and the following year was even higher, which may in part account for the sharp reduction in Salsola cover we saw between years, even in no-herbicide and no-grazing plots. In fact, Figure 2 shows that by 2019 and 2020, nonnative cover increased substantially, coinciding with the substantial drop in Salsola cover. We anticipate that longer-term collection of weather and Salsola cover data will clarify this relationship.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/inp.2022.9
Acknowledgments
We thank Fred Hrusa for identifying Salsola specimens. We thank Bill Whitney from the Topo Ranch and Todd and Wendy Sans from Sans-Topo Ranch for hosting our plots. We also thank Hedgerow Farms and L.A. Hearn Seed Company for seed donations. No conflicts of interest have been declared.









 Open access
Open access