


AUDs strongly run in families (Cotton, Reference Cotton1979). While twin and adoption studies have unequivocally demonstrated a substantial genetic contribution to the etiology of AUDs (e.g., Cadoret et al., Reference Cadoret, Troughton and O'Gorman1987; Cloninger et al., Reference Cloninger, Bohman and Sigvardsson1981; Goodwin et al., Reference Goodwin, Schulsinger, Hermansen, Guze and Winokur1973; Heath et al., Reference Heath, Bucholz, Madden, Dinwiddie, Slutske, Bierut and Martin1997; Kaij, Reference Kaij1960; Kendler et al., Reference Kendler, Prescott, Neale and Pedersen1997; Kendler et al., Reference Kendler, Ji, Edwards, Ohlsson, Sundquist and Sundquist2015; Pickens et al., Reference Pickens, Svikis, McGue, Lykken, Heston and Clayton1991; Prescott et al., Reference Prescott, Aggen and Kendler1999; Sigvardsson et al., Reference Sigvardsson, Bohman and Cloninger1996), there remain at least three important unresolved questions about the familial aggregation of AUDs. First, some twin studies suggest modest or substantial familial-environmental influences on risk for AUDs (Allgulander et al., Reference Allgulander, Nowak and Rice1991; Gurling et al., Reference Gurling, Oppenheim, Cook, Farrel and Murray1993; Kaij, Reference Kaij1960; Kendler et al., Reference Kendler, Prescott, Neale and Pedersen1997; Pickens et al., Reference Pickens, Svikis, McGue, Lykken, Heston and Clayton1991), while other reports find no evidence for such effects (Heath et al., Reference Heath, Bucholz, Madden, Dinwiddie, Slutske, Bierut and Martin1997; Prescott & Kendler, Reference Prescott and Kendler1999; Reed et al., Reference Reed, Page, Viken and Christian1996). The magnitude of the shared environmental variance, however, is typically small (Kaij, Reference Kaij1960, is an exception). In the presence of substantial genetic influences, quite large samples of twins are needed to detect evidence for shared environment (Neale et al., Reference Neale, Eaves and Kendler1994).
Second, several early twin and adoption studies reported evidence for quantitative sex effects in the genetic factors for AUD — that genetic effects were stronger in males than in females (Bohman et al., Reference Bohman, Sigvardsson and Cloninger1981; Goodwin et al., Reference Goodwin, Schulsinger, Hermansen, Guze and Winokur1973; McGue et al., Reference McGue, Pickens and Svikis1992). The studies that found differential genetic effects as a function of sex, however, often relied on small samples, especially of females. Most recent larger studies (e.g., Heath et al., Reference Heath, Bucholz, Madden, Dinwiddie, Slutske, Bierut and Martin1997; Prescott et al., Reference Prescott, Aggen and Kendler1999) have been unable to detect genetic differences between the sexes.
Third, five twin studies presented results on resemblance for AUDs in OS dizygotic (DZ) twin pairs, and thus were able to test for the presence of qualitative sex effects for AUDs — that the genetic risk factors were not entirely the same in males and females. This is detected largely by comparing the magnitude of the correlation in same-versus OS DZ pairs. One study found evidence for such an effect for AUDs (Prescott et al., Reference Prescott, Aggen and Kendler1999), two did not (Heath et al., Reference Heath, Bucholz, Madden, Dinwiddie, Slutske, Bierut and Martin1997; Magnusson et al., Reference Magnusson, Lundholm, Goransson, Copeland, Heilig and Pedersen2012), and two did not test for it (Caldwell & Gottesman, Reference Caldwell and Gottesman1991; Knopik et al., Reference Knopik, Heath, Madden, Bucholz, Slutske, Nelson and Martin2004). Demonstration of qualitative sex effects typically requires quite large samples (Prescott & Gottesman, Reference Prescott and Gottesman1993).
A recent meta-analysis of twin and adoption studies of AUD (Verhulst et al., Reference Verhulst, Neale and Kendler2015) addressed all three of these questions. Shared environmental effects were present, although marginally statistically significant, and estimated at 10%. While aggregate heritability estimates were modestly higher in males than females, this difference was not statistically significant. No evidence was found in the meta-analysis for qualitative sex effects.
We here report on a study of AUD in a National Swedish sample of twins with known zygosity (n = 18,829) and reared-together full and half siblings with a maximum of 5 years age difference (n = 769,087). AUD was ascertained from medical, criminal, and pharmacy registries. We examine the role of genetic and environmental risk factors in the etiology of AUD and evaluate specifically the presence of (1) shared environmental effects, (2) quantitative genetic sex effects, and (3) qualitative genetic sex effects.
Before we present these analyses, however, we examine a more foundational question. We obtained our AUD cases from three registers in Sweden: medical, crime, and pharmacy. In our larger (and more informative) sample of males, we address the question of the degree to which AUD ascertained from these three different sources share genetic and environmental determinants.
Methods
Sample
We linked nationwide Swedish registers via the unique 10-digit identification number assigned at birth or immigration to all Swedish residents. The identification number was replaced by a serial number to ensure anonymity.
From the Swedish Twin registry, we selected all twin pairs with birth years from 1960 to 1990 with known zygosity (n = 18,829) and from the Swedish Multi-Generation Registry, we identified Swedish-born full- and half-sibling pairs, born between 1960 and 1990 and within 5 years of each other (n = 769,087). We assessed, using the Swedish national census and total population registries, the cohabitation status of the sibling pairs as the proportion of possible years lived in the same household until the oldest turned 18, the age of majority in Sweden. We included pairs reared together, defined as living together for 80% of the possible years. The Swedish Total Population Registry provided information about: sex, country of birth, migration, and mortality. Individuals who died or emigrated before age 16 were excluded from the dataset. As the modeling assumes that the correlation between pairs is zero, we randomly selected one pair from each family, meaning that if one individual and his or her (full or half) sibling are included in the model, all other possible pairs of full or half siblings from that family are excluded. As detailed elsewhere (Lichtenstein et al., Reference Lichtenstein, de Faire, Floderus, Svartengren, Svedberg and Pedersen2002), zygosity in the same-sex pairs from the twin registry was assigned using standard self-report items from mailed questionnaires which, when validated against biological markers, were 95–99% accurate.
Measures
The following registries were used to create the AUD dataset: the Swedish Hospital Discharge Register, containing hospitalizations from 1964 to 2010; the Outpatient Care register, containing information from all out-patient clinics between 2001 and 2010; the Prescribed Drug register containing all prescriptions in Sweden picked up by patients from 2005 to 2010; the Primary Health Care Register, containing outpatient diagnoses from 2001 to 2007 for 1 million patients from Stockholm and middle Sweden; the Swedish Crime Register, which includes complete national data on all convictions in the lower court from 1973 to 2011; and the Swedish Suspicion Register, which included complete national data on individuals strongly suspected of crime from 1998 to 2011.
AUD was identified in three ways. First was a medical diagnosis from the medical registers: ICD-8 codes: 571.0, 291, 303, 980; ICD-9 codes: V79B, 305A, 357F, 571A, 571B, 571C, 571D, 425F, 535D, 291, 303, 980; and ICD-10 codes: E244, G312, G621, G721, I426, K292, K700, K701, K702, K703, K704, K705, K706, K707, K708, K709, K852, K860, O354, T510, T512, T511, T513, T514, T515, T516, T517, T518, T519, F101, F102, F103, F104, F105, F106, F107, F108, and F109. Second was registration in the prescription registry for well-recognized drugs to treat AUD using the ATC Classification System codes N07BB01 (disulfiram), N07BB03 (acamprosate), and N07BB04 (naltrexone). Third was registration in the suspicion and conviction register for at least two records of drunk driving (suspicion code: 3005 and law 1951:649 and Paragraph 4 and 4A) or drunk in charge of maritime vessel (suspicion code: 3201 and law 1994:1009 and Paragraphs 4 and 5 and Chapter 20). AUD was here defined as registration in one or more of these registries.
Statistical Model
We assumed a liability threshold model with four sources of liability to AUD: additive genetic (A), shared environment (C), shared environment unique to twins (T), and unique environment (E). The model assumes that MZ twins share all of their genes, while DZ twins and full siblings share half, and half siblings a quarter of their genes identical by descent. Shared environment reflects family and community experiences that render the twins and siblings more similar for the phenotype in question, while unique environment includes random developmental effects, environmental experiences not shared by siblings, and random error.
First, in males only, we used a trivariate model to investigate to what extent genetic and environmental factors are the same for all three phenotypes, utilizing both an independent and a common pathway model. In the common pathway model, the common part is represented by a latent phenotype with distinct loadings on the three observed phenotypes. The variance of the latent phenotype is decomposed in four sources of variance and each of the phenotypes also has unique specific variance components, denoted As, Cs, Ts, and Es. In the independent pathway, on the other hand, each of the variance components (A, C, T, and E) for the respective three phenotypes consists of two parts: one that is common to all (Ac, Cc, Tc, and Ec) and one that is specific to each one of them (As, Cs, Ts, and Es). Consequently, the common pathway model is a special case of the independent pathway model.
Second, we used a univariate model of AUD utilizing both sexes and tested for quantitative and qualitative sex effects — that is, do the estimates in our model (A,C,T, and E) differ in magnitude in males and females and are the same genetic and environmental factors influencing risk of AUD in both sexes? Qualitative sex effects are captured by three parameters, here denoted rgs, rcs, and rts that reflect, respectively, the degree to which the genetic (g), shared (c) or twin environmental (t) risk factors are correlated between the two sexes (s).
Estimated path coefficients and variance components are presented with likelihood-based 95% CIs, and models are compared using the Akaike information criterion (AIC).
Results
Sample
For each of the 11 twin-sibling relationships that we examined in this report, Table 1 presents the sample size, prevalence of total AUD, tetrachoric correlation and, for the sibling pairs, the mean percentage of cohabitation. Across relationship types, sample sizes range over two orders of magnitude, being smallest in twin pairs, intermediate in reared-together half siblings, and largest in reared-together full siblings. The prevalence of AUD is approximately twice as great in males as females and lower in the same-sex twin pairs, where zygosity classification depended on the return of questionnaires. Also noteworthy is the substantially higher rates of AUD in the half versus the full siblings and the quite high rates of cohabitation among the full and half siblings included in the analysis.
TABLE 1 Sample Sizes, Tetrachoric Correlations, and Cohabitation Status for Twin and Sibling Samples Used in These Analyses

Trivariate Analysis of Registration for AUD in Male–Male Pairs
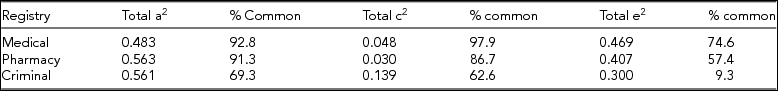
In females, we lacked concordant twin pairs for analyses involving cases of AUD ascertained through the criminal registry, so trivariate modeling was only feasible in males. The tetrachoric correlations within and between twin and sibling pairs for our three forms of AUD are seen in Appendix Tables A1 and A2. Model-fitting results are presented in Table 2 for ACE and ACTE common and independent pathway models. By a considerable margin, the AIC was best for the independent pathway ACE model. Parameter estimates from this model are provided in Tables 3 and 4. The heritabilities of the three forms of AUD were relatively similar, being highest in pharmacy ascertained (56.3%) and lowest for medically ascertained AUD (48.3%). Shared environmental effects were quite small for medical and pharmacy ascertained and somewhat greater for criminally ascertained AUD.
For AUD ascertained through the medical and pharmacy registries, over 90% of the genetic variance was shared. For AUD ascertained through the criminal registry, this figure was ~70%. A similar pattern was seen for shared environmental effects, where the vast majority of variance for medical- and pharmacy-ascertained AUD was explained by the common factor, while for the criminal registry the estimate was lower — here around 60%.
AUD in Both Sexes in Twins and Full and Half Siblings
Turning to modeling total AUD in both males and females, we first note that while the tetrachoric correlations (±SE) were high and equal in male–male and female–female MZ pairs (+0.65±0.05 and +0.65±0.06, respectively), large differences were seen in the DZ pairs, with a much higher correlation in female–female (+0.49±0.10) than in male–male pairs (+0.34±0.08) (Table 1). In full siblings — where the precision of estimation was much greater — we see the reverse pattern, with the correlation being considerably greater in male–male pairs (+0.35±0.01) than female–female pairs (+0.23±0.02). Among half-siblings, the correlations in male–male pairs exceeded only modestly those seen in female–female pairs (+0.21±0.03 vs. +0.19±0.06, respectively).
TABLE 2 Model-Fitting Results for Alcohol Use Disorder Ascertained from Medical, Criminal, and Pharmacy Registries

*best fit model.
TABLE 3 Proportions of Variance Estimated From the Best Fit Trivariate Model

TABLE 4 Total Heritability and Shared Environmental and Individual Specific Environmental Components of AUD as Assessed Via the Medical, Pharmacy, and Criminal Registries
We had three OS pair correlations. In both DZ twins and half siblings, the OS pair correlations were slightly lower than the lowest of the same-sex correlations. In full siblings, however, the OS pair correlations were between those seen for the female–female and male–male pairs.
Model-Fitting Results and Parameter Estimates
We began by comparing the fit of a simple ACE and ACTE model with no sex effects. The fit was much worse with the ACE versus ACTE model (-2,559,904 vs. -2,736,456), so our further model-fitting, as outlined in Table 5, focused solely on ACTE models. Model 1, which contains no qualitative sex effects, so that r gs, r cs, and r ts are constrained to unity, was our base model. In model 2, we equated all parameters of the model to equality across the sexes and the fit of the model deteriorated substantially. In model 3, we constrained r gs, r cs, and r ts to equality and the model estimated them at +0.95. This model fits modestly better than model 1, indicating some qualitative genetic and/or environmental effects on AUD between the sexes. In models 4 through 6, we allowed, respectively, r gs, r cs, and r ts to be free while constraining the other two parameters to unity. Model 4 fit best, although by only a very small margin in the AIC value. In this model, r gs was estimated at +0.91.
TABLE 5 Model-Fitting Results for Total Alcohol Use Disorder in Both Sexes in Twins and Reared-Together Full and Half-Siblings

*best fit model; a refers to additive genetic effects, c to shared environment, t to special twin environment, and e to unique environment. The subscripts r ts, r gs, and r cs refer to correlations between the sexes for, respectively, special twin environment, genetic effects, and shared environment.
The parameter estimates for the best fit model 4 are seen in Table 6. For males, heritability is estimated at 57%, with modest effects of shared and special twin environment estimated at 6% and 2%, respectively. Unique environmental effects were estimated at 35%. Parameter estimates in females differed substantially from that seen in males. Heritability estimates were considerably lower (22%), and estimates for both shared and special twin environmental effects much stronger (12% and 29%, respectively). Unique environmental effects were similar across the two sexes (37% in females, and 35% in males).
TABLE 6 Parameter Estimates and Confidence Intervals from Best-Fitting Model for Alcohol Use Disorders in Males and Females

Given the striking differences seen in parameter estimation in males and females in our entire twin-sibling sample, we repeated model fitting only with the twins. In the twins, we could constrain rg to unity with a slight improvement in AIC. Estimates of a2 in males and females were, respectively, 63 and 31% while estimates of c2 in the two sexes were 3% and 34%.
Discussion
Our initial analyses sought to clarify the degree to which AUD detected through the medical, pharmacy, and crime registries reflected the same versus distinct genetic and environmental risk factors. For AUD cases detected in the medical and pharmacy registry, over 90% of the genetic and 85% of the shared environmental variance reflected the common factor. However, for AUD from the criminal registry, the proportion was lower (69% and 63%, respectively). About a third of the genetic and shared environmental variance, and nearly all of the unique environmental variance for criminal AUD registration, is not shared with AUD detected through medical or pharmacy records and might reflect risk factors specifically associated with externalizing traits. Overall, our independent pathway model suggested that AUD ascertained from the three registries shared a large proportion of the same genetic and environmental risk factors.
We then turned to address the three issues related to twin studies of AUD we outlined above. We examine these in turn. In the recent meta-analysis of twin studies of AUD (Verhulst et al., Reference Verhulst, Neale and Kendler2015), estimates of shared environmental influences were modest and of marginal statistical significance. Consistent with the meta-analytic results, we found evidence for modest c2 effects in both males and females. However, because we examined both twins and siblings, we had the ability to estimate twin-specific environmental effects and found them to be quite large in females, while barely present in males. These results raise the question of whether the familial-environmental effects on AUD might have been under-estimated by prior studies, which by focusing only on twins lacked the ability to detect twin-specific environmental effects.
The meta-analysis of twin and adoption studies of AUD found estimates that were modestly but not significantly higher in males than females (Verhulst et al., Reference Verhulst, Neale and Kendler2015). In our study, substantial differences in heritability of AUD emerged between the sexes, with estimates in males more than twice as high as those observed in females. These results are similar to those reported in a relatively small, clinically ascertained twin sample by McGue et al. (Reference McGue, Pickens and Svikis1992) but inconsistent with the larger population-based studies of AUD in Virginia (Prescott et al., Reference Prescott, Aggen and Kendler1999) and Australia (Heath et al., Reference Heath, Bucholz, Madden, Dinwiddie, Slutske, Bierut and Martin1997). We cannot ascribe our different findings to the presence of siblings and half siblings in our analyses, as when we modeled the results just in twins, we found a similar large difference in heritability across the sexes. What is clear by an examination of the raw correlations is that in our sample we see consistent evidence for strong shared environmental influences (and hence reduced heritability) on AUD in the female–female twin and sibling pairs. In particular, we find (1) DZ twin correlations much greater than 50% of the MZ correlations, (2) DZ twin correlations much greater than sibling correlations, and (3) half-sibling correlations much greater than 50% of the full-sibling correlations. So, the evidence for strong shared environmental effects in females is coming from multiple sources within our sample.
Further research will be required to try to understand the origin of the large sex difference in sources of individual differences in liability to AUD in our sample. Could it result from our mode of ascertainment of AUD? We used the same registries to ascertain drug abuse, and in twin-sibling models found higher estimates of a2 and lower estimates of c2 in females than in males, the opposite of what we have found for AUD (Kendler et al., Reference Kendler, Maes, Sundquist, Ohlsson and Sundquist2013). It seems therefore unlikely that our evidence for quantitative sex effects for AUD arose from any general sex-related bias in our registries. Two questions seem particularly relevant for further research. Would these sex-effects be found if we generalized our sampling further to include sibling, half-sibling, and cousin pairs who both did and did not reside together in childhood, as well as step-siblings? Can we identify social factors governing alcohol exposure and heavy use in males and females in Sweden that might be responsible for these effects?
The meta-analysis also failed to find evidence for qualitative sex effects in AUD — that is, genetic risk factors operating differently in males and females (Verhulst et al., Reference Verhulst, Neale and Kendler2015). We had, in our analyses, evidence for qualitative sex effects on risk for AUD. The best fit model, by a modest margin, suggested that genetic influences on AUD differed slightly in men and women, with an estimated genetic correlation of +0.91. Although perhaps not correlated perfectly, our findings broadly agree with the findings from the meta-analysis that genetic risk factors for AUD are quite highly correlated in males and females.
Limitations
These results should be interpreted in the context of three potentially important methodological limitations. First, we detected subjects with AUD from official registry records. While this method does not require respondent cooperation or accurate recall and reporting, it could produce both false negative and false positive diagnoses, and we cannot precisely estimate these biases. Given that the population prevalence of AUD in this sample is much lower than estimates from most epidemiologic surveys (Grant et al., Reference Grant, Goldstein, Saha, Chou, Jung, Zhang and Hasin2015; Kessler et al., Reference Kessler, McGonagle, Zhao, Nelson, Hughes, Eshleman and Kendler1994) including one from nearby Norway (Kessler et al., Reference Kessler, McGonagle, Zhao, Nelson, Hughes, Eshleman and Kendler1994; Kringlen et al., Reference Kringlen, Torgersen and Cramer2001), false negative diagnoses are much more likely than false positive ones. The validity of our AUD definition is supported by the high rates of concordance for registration across our difference ascertainment methods (Kendler et al., Reference Kendler, Ji, Edwards, Ohlsson, Sundquist and Sundquist2015).
Second, we could only examine same-sex twin pairs whose zygosity was known as a result of at least one member responding to a mailed questionnaire. (This was not needed for opposite-sex twin or sibling pairs.) As expected, AUD is associated with a reduced probability of returning questionnaires so the rate of AUD was lower in pairs with known zygosity. This is known as ‘concordance-dependent’ ascertainment, where the probability of known zygosity will be lowest in pairs concordant for AUD, intermediate in those discordant for AUD, and highest in those where neither twin has AUD (Kendler & Eaves, Reference Kendler and Eaves1989). Prior simulations suggest that given the level of differential ascertainment likely in our data, biases in parameter estimates are probably modest and most likely result in slight under-estimations of genetic and shared environmental effects and overestimation of the effects of the individual-specific environment (Kendler & Eaves, Reference Kendler and Eaves1989).
Third, we did not in our analyses examine in twin and sibling pairs concordant for AUD resemblance in their mode of ascertainment for AUD. Of particular interest, in the siblings where we large enough numbers of pairs concordant for AUD to analyze confidently, resemblance for registration in the medical, criminal, and pharmacy registries, as assessed by a tetrachoric correlation, were moderate in same sex pairs (largely +0.20 to +0.35). However, they were much lower in opposite-sex siblings (~0 to +0.12), suggesting sex effects on the patterns of registration for alcohol problems in the medical, criminal, and pharmacy registries.
Financial Support
This project was supported by the grants AA023534 and AA021399 from the National Institute of Alcohol Abuse and Alcoholism, the Swedish Research Council (K2012-70X-15428-08-3), the Swedish Research Council for Health, Working Life and Welfare (In Swedish: Forte; Reg.nr: 2013-1836), the Swedish Research Council (2012-2378; 2014–10134) and FORTE (2014-0804) as well as ALF funding from Region Skåne awarded.
Conflict of Interest
The authors have no conflicts of interests to declare.
Ethical Standards
The authors assert that all procedures contributing to this work comply with the ethical standards of the relevant national and institutional committees on human experimentation and with the Helsinki Declaration of 1975, as revised in 2008.
Appendix
TABLE A1 Tetrachoric Correlation Matrices in Male–Male Twins for Alcohol Use Disorder Ascertained From Medical, Pharmacy, and Criminal Registries: Monozygotic Twins Above and Dizygotic Twins Below the Diagonal

TABLE A2 Tetrachoric Correlation Matrices in Male–Male Twins for Alcohol Use Disorder Ascertained From Medical, Pharmacy, and Criminal Registries: Full Siblings Reared Together Above and Half-Siblings Reared Together Below the Diagonal









