



Introduction
Severe population declines amongst African elephants Loxodonta sp. are occurring throughout their range because of poaching for ivory or habitat reduction and fragmentation (Maisels et al., Reference Maisels, Strindberg, Blake, Wittemyer, Hart and Williamson2013; Gobush et al., Reference Gobush, Edwards, Balfour, Wittemyer, Maisels and Taylor2021a,Reference Gobush, Edwards, Maisels, Wittemyer, Balfour and Taylorb). Today, c. 415,000 African elephants remain in 37 countries (Thouless et al., Reference Thouless, Dublin, Blanc, Skinner, Daniel and Taylor2016). After a long-standing debate, African elephants are now considered by IUCN to be two distinct species (Hart et al., Reference Hart, Gobush, Maisels, Wasser, Okita-Ouma and Slotow2021): the savannah elephant Loxodonta africana, categorized as Endangered on the IUCN Red List (Gobush et al., Reference Gobush, Edwards, Balfour, Wittemyer, Maisels and Taylor2021a) and the forest elephant Loxodonta cyclotis, categorized as Critically Endangered (Gobush et al., Reference Gobush, Edwards, Maisels, Wittemyer, Balfour and Taylor2021b).
The two species are known to have their own behavioural and ecological features (Roth & Douglas-Hamilton, Reference Roth and Douglas-Hamilton1991; Grubb et al., Reference Grubb, Groves, Dudley and Shoshani2000; Turkalo & Fay, Reference Turkalo, Fay, Weber, White, Vedder and Naughton-Treves2001). Some anatomical and morphological characteristics such as body size, ear shape, tusk orientation and depth of the temporal fossae are sufficiently different to distinguish the two species (Fallon, Reference Fallon1944; Morrison-Scott, Reference Morrison-Scott1947; Pfeffer, Reference Pfeffer1989; Shoshani, Reference Shoshani1993; Grubb et al., Reference Grubb, Groves, Dudley and Shoshani2000; Debruyne, Reference Debruyne2003; Fowler & Mikota, Reference Fowler and Mikota2006). However, most morphological studies comparing the two species have focused on bones/skulls and not on living individuals (van der Merwe et al., Reference van der Merwe, Bezuidenhout and Seegers1995; Grubb et al., Reference Grubb, Groves, Dudley and Shoshani2000; Debruyne, Reference Debruyne2003). Nevertheless, differences depending on age and sex as well as regional morphological variations observed in the savannah elephant, particularly in the shape and size of their ears and tusks, could limit the use of these morphological characteristics in practice (Hanks, Reference Hanks1972; Pfeffer, Reference Pfeffer1989).
Despite their habitat differences, the geographical distributions of the two species overlap, and both occur in the forest–savannah ecotones of Central and West Africa (Pfeffer, Reference Pfeffer1989; Groves & Grubb, Reference Groves and Grubb2000), although there have been few morphological observations of intermediate elephants reported in these areas (Pfeffer, Reference Pfeffer1989; Groves & Grubb, Reference Groves and Grubb2000; Debruyne, Reference Debruyne2003). To date, genetic studies have provided evidence of hybrids in five locations: Garamba National Park in the north-eastern Democratic Republic of the Congo (DRC) (Roca et al., Reference Roca, Georgiadis, Pecon-Slattery and O'Brien2001, Reference Roca, Georgiadis and O'Brien2005; Comstock et al., Reference Comstock, Georgiadis, Pecon-Slattery, Roca, Ostrander, O'Brien and Wasser2002; Mondol et al., Reference Mondol, Moltke, Hart, Keigwin, Brown, Stephens and Wasser2015; Kim & Wasser, Reference Kim and Wasser2019); along the border of the DRC and Uganda; the northern Central African Republic; the Gourma region in Mali; and along the Pendjari–Arli complex of West Africa on the Benin–Burkina Faso border (Mondol et al., Reference Mondol, Moltke, Hart, Keigwin, Brown, Stephens and Wasser2015; Kim & Wasser, Reference Kim and Wasser2019).
To date, no study combining genetics and morphology has been able to shed light on the phenotype of hybrid elephants. Yet the study of phenotypes (the set of observable characteristics of all or part of an organism) is central to many disciplines in biology (Houle et al., Reference Houle, Govindaraju and Omholt2010). In addition, we have no information about whether hybrids could be phenotypically intermediate between their parents, closer to one parent or outside the range of variation of both parents' species. As hybrids are fertile (Mondol et al., Reference Mondol, Moltke, Hart, Keigwin, Brown, Stephens and Wasser2015), we can assume they have a wide range of phenotypes depending on their hybridization rate and whether their mates are hybrids or pure forest or savannah elephants (Roca et al., Reference Roca, Georgiadis and O'Brien2005; Mondol et al., Reference Mondol, Moltke, Hart, Keigwin, Brown, Stephens and Wasser2015). Our study aims to confirm that the two species of African elephant can be discriminated using a set of six proposed morphological criteria, and that in a hybridization zone both species can be found as well as individuals differing from both the savannah and the forest phenotype, which could emerge from the reproduction of the two species.
This study took place in the hybridization zone at the DRC–Uganda border in the forested Sebitoli area in the north of Kibale National Park, Uganda. This zone is located 60 km north of the nearest area of savannah in Queen Elizabeth National Park and is linked to it by a corridor that facilitates the movement of elephants between the two Parks (Fig. 1). The presence of the two species and the hybrids has already been documented in the south of Kibale National Park. However, neither the proportions of each species nor the history or dynamics of the populations are known (Mondol et al., Reference Mondol, Moltke, Hart, Keigwin, Brown, Stephens and Wasser2015). Local farmers and villagers in the Sebitoli area have also reported the presence of two types of elephants distinguishable by their size, colour and behaviour (Capelot, Reference Capelot2013; SK, pers. comm., 2012; JB, pers. comm., 2019). As direct observations and access to biological samples are challenging because of the dense undergrowth, swamps and mountains of the Sebitoli terrain, all of which add to the danger of closely approaching elephants, which are likely to charge if threatened (Omeja et al., Reference Omeja, Lawes, Corriveau, Valenta, Sarkar, Paim and Chapman2016), we used camera traps and developed an assignment key for the elephant species to document phenotypes in this area.

Fig. 1 The Democratic Republic of the Congo–Uganda border, and the Sebitoli area in Kibale National Park (Uganda), with locations of the camera traps.
Study area
The 795 km2 Kibale National Park in south-west Uganda is a mosaic of mature forest, successional forest (formerly used for agriculture), grassland, woodland, lakes and wetlands (Chapman & Lambert, Reference Chapman and Lambert2000). The movement of elephants between Kibale National Park (forested area) and DRC can only take place in the south via the connection between Queen Elizabeth National Park (savannah area) and Virunga National Park (mixed forest–savannah area) because of the high altitude of the Rwenzori Mountains between the two countries. Migrations between the Virunga and Queen Elizabeth National Parks have been observed since the 1960s, mainly from the DRC to Uganda (Keigwin et al., Reference Keigwin, Wabukawo, Wasser and Chapman2016). In the extreme north of Kibale National Park, the Sebitoli research area, monitored by the Sebitoli Chimpanzee Project, covers c. 25 km2 of forest logged in the 1970s. Today, this area comprises regenerating forest (70%), with only 14% being old-growth forest, surrounded by farmlands and tea and eucalyptus plantations (Bortolamiol et al., Reference Bortolamiol, Cohen, Potts, Pennec, Rwaburindore and Kasenene2014).
Methods
We conducted this study in two stages. Firstly, we tested the reliability of six morphological criteria to distinguish the two species. Secondly, after confirmation of the relevance of these criteria, we studied the Sebitoli elephant population.
Choice and verification of morphological criteria
To distinguish morphologically between the species, we selected criteria that are easily seen on the camera-trap videos and do not require precise measurements but rather observations of orientation or ratios between elephant body parts. We used six criteria visible when an elephant is in profile or facing the camera trap: the position of the lower lobe of the ear compared to the mandible line (Ear, profile); the distance between the tusks and trunk (Space, profile), tusk orientation (TuskProfile, profile and TuskFront, facing), temporal fossae (Forehead, facing), and back curvature (Back, profile). Each criterion is separated into two or three categories (Table 1), as illustrated in Fig. 2 and Supplementary Fig. 1.
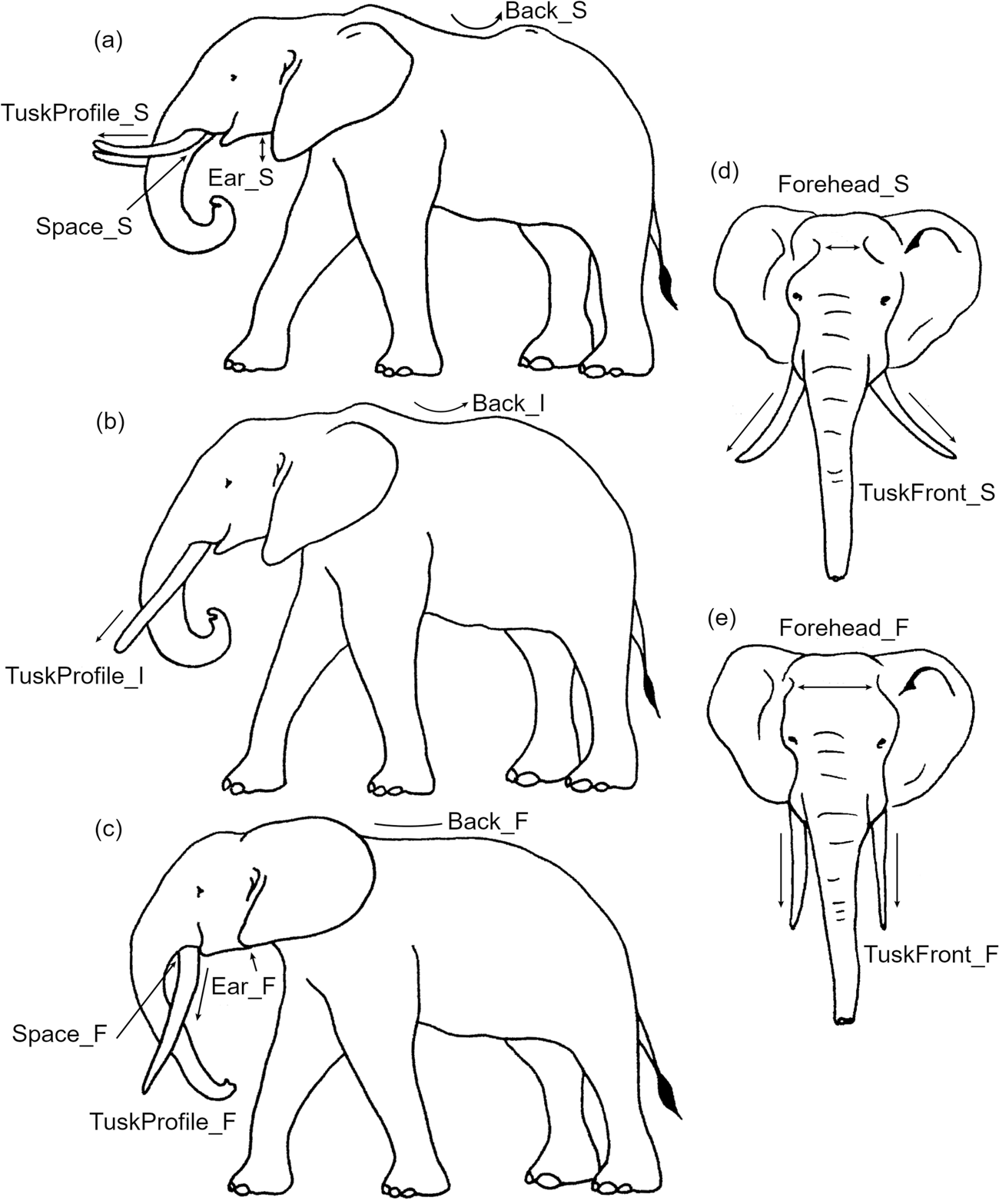
Fig. 2 Illustrations of the six morphological criteria and their states (Table 1) used to distinguish between the two species, showing the typical phenotype of the savannah (a, d) and the forest elephant (c, e), and an example of an intermediate phenotype (b).
Table 1 Summary of the characteristics of the six criteria and their states (S, savannah; F, forest; I, intermediate) used to distinguish savannah Loxodonta africana and forest Loxodonta cyclotis elephants (Fig. 1).

To test our combination of criteria, we selected photographs of 171 savannah elephants and 125 forest elephants (296 in total), from 11 countries (Supplementary Material 1). To quantify the phenotypic distinction between savannah and forest elephants, we applied a supervised machine learning approach to the reference images. We applied the simple and robust K-nearest neighbours algorithm (Ripley, Reference Ripley2007), assigning a class to an object according to the class to which its nearest K neighbours belong, using the following steps: (1) numerically coding the qualitative data in a sparse matrix; this format lends itself to machine learning by suppressing the ordinal effect of the variables; (2) managing missing data by creating a first dataset keeping only specimens whose variables had no missing data, and a second dataset replacing the missing data with the most frequent class of the variable; (3) splitting the data randomly into a training dataset (80%) and a test dataset (20%); and (4) training and testing the model by varying K from 1 to 10. All steps were carried out using the Scikit-learn library (Pedregosa et al., Reference Pedregosa, Varoquaux, Gramfort, Michel, Thirion and Grisel2011) in Python (Python Software Foundation, Wilmington, USA).
Study of elephant morphology in the Sebitoli hybridization zone
We deployed camera traps inside the forest and at its edge (Fig. 1), fixing them to the trunk of a tree c. 1 m above ground level, either on the research trail system or along elephant tracks. There were elephants in 1,408 of the 12,483 videos captured during November 2017–March 2018 by 14 camera traps (13 high-definition Reconyx XR-6 Ultrafire video traps, Reconyx, Holmen, USA; one high-definition Bushnell Trophy Cam HD Maxvideo trap, Bushnell, Overland Park, USA). In these videos, we analysed 1,215 occurrences (i.e. observation of one individual elephant in a single video clip or observations of one individual in several consecutive videos).
To study the global phenotypic variability by linking specimens and variables, we performed a multiple correspondence analysis (MCA; Abdi & Williams, Reference Abdi and Williams2010) using the mca function of the FactoMineR library (Lê et al., Reference Lê, Josse and Husson2008). We conducted this analysis on the complete dataset (reference photographs and the Sebitoli sample), with no missing data. To visualize potential clusters, we performed a kernel density plot on the MCA using the kde function from the package ks (Charcón & Duong, Reference Chacón and Duong2018) in R (Supplementary Fig. 2; R Core Team, Reference R Core Team2019).
We made a species assignment key (Supplementary Table 1) based on the six criteria previously validated in stage 1 of the survey, to facilitate the distinction between forest and savannah elephants from direct and indirect observations in the field. The criteria were divided into two categories (main criteria: C1, C2, C3; secondary criteria: C4, C5, C6) according to their visibility on the camera-trap footage; the secondary criteria were often barely visible. As elephants with intermediate phenotypes could also be seen in these forest savannah ecotones, we included the possibility of intermediate phenotypes in the species assignment key. We also noted when each criterion was not visible. We then used this key to determine the phenotype of the individual. When possible we determined the sex and age class (infant, juvenile, subadult or adult) of the elephant.
To validate our hypothesis that the two species and hybrids are present in the Sebitoli area, we cross-referenced our results from the species assignment key with results from the unsupervised K-means approach (Hartigan & Wrong, Reference Hartigan and Wong1979) using the kmeans function in R. To partition phenotypic variability into K clusters, this algorithm minimizes the sum of the squares of the distances from a point to the average of the points of its cluster. As the K-means algorithm requires the number of groups to be defined, we conducted two analyses, one with two groups and one with three groups, to be able to detect a possible group of intermediate phenotypes. The two approaches were carried out on the dataset comprising the 705 Sebitoli elephants, with missing data. We then tested the sex and the age bias using a Pearson's χ 2 test.
Results
Validation of the reliability of the morphological criteria
The combination of the six morphological criteria facilitated a good description of the species, with 100% being well assigned (K-neighbour = 1) when using observations without missing data and 91.67% being well assigned (K-neighbour = 1) when missing data were replaced with the most frequent class of the variable.
Sebitoli elephant morphology
The multiple correspondence analysis of the six morphological variables revealed that the two most important principal components encompassed 54.17% of the total variance (Fig. 3). Reference savannah elephants showed the lowest variability, reference forest elephants showed medium variability and elephants surveyed in Sebitoli showed high variability. Clustering occurred between the two species, with a marked gap along the first axis, which is explained mostly by the variables Space, TuskProfile and Forehead (Supplementary Fig. 3). The second axis is mostly explained by the variables Space and TuskProfile. The surveyed Sebitoli elephants do not seem to be a cohesive group: they overlap completely with the forest elephant variability and slightly overlap with savannah elephant variability. Thus, there are three groups (Supplementary Fig. 2): (1) savannah elephants from the reference sample and part of the Sebitoli sample, mostly explained by the states of the variables Ear_S, Space_S, TuskProfile_S, TuskFront_S, ForeHead_I and Back_I; (2) forest elephants from the reference sample and part of the Sebitoli sample, mostly explained by the states of the variables Ear_F, Space_F, TuskProfile_F, TuskFront_F, Forehead_F and Back_F; and (3) the rest of the Sebitoli sample.

Fig. 3 Scatter plot of the two first axes of the multiple correspondence analysis (with the per cent of variance accounted for by each axis) performed on the six morphological criteria and their states (Table 1), and the three populations (savannah and forest elephants and the Sebitoli elephant population).
The use of the species assignment key enabled the identification of three types of phenotypes: forest (12.1%), savannah (36.8%) and an intermediate group (51.1%). We then compared the results obtained from this method with results from the K-means analysis (Supplementary Table 2). All of the specimens assigned to the forest species using the species assignment key were assigned to Group 3, in which no savannah elephants but 50% of the intermediate-phenotype individuals were included. Group 2 comprised 78.5% of the individuals assigned to the savannah elephant phenotype, 15.5% of the individuals assigned to the intermediate phenotype and no forest elephant phenotypes. Group 1 includes 34.5% of the individuals assigned to the intermediate phenotype and 21.5% of individuals assigned to the savannah phenotype. The three groups obtained from the K-means analysis partially match the three groups (forest, savannah and intermediate) obtained from the species assignment key analysis, with Group 1 being the intermediate phenotype, Group 2 being the savannah phenotype and Group 3 being the forest phenotype. Sex and age biases were taken into account (Supplementary Material 2).
Discussion
We first validated, using a K-nearest neighbours algorithm, the relevance of the six morphological criteria for discriminating between the savannah and forest phenotypes by testing them on 296 individuals whose geographical origins were known. When applied to a set of 1,408 videos recorded in Sebitoli, in the hybridization area in Uganda, these criteria produced three groups of phenotypes using an unsupervised K-means approach (the expected savannah and forest phenotypes and a third group, present in 51.1% of the videos). This third group could correspond to hybrid individuals, whose presence had been noted previously in the south of our study area (Mondol et al., Reference Mondol, Moltke, Hart, Keigwin, Brown, Stephens and Wasser2015; Bonnald et al., Reference Bonnald, Utge, Kuhner, Wasser, Asalu, Okimat and Krief2021).
To date, there has been no genetic study of individuals whose phenotypes are known. As hybrids are fertile (Mondol et al., Reference Mondol, Moltke, Hart, Keigwin, Brown, Stephens and Wasser2015; Bonnald et al., Reference Bonnald, Utge, Kuhner, Wasser, Asalu, Okimat and Krief2021), we assume there are several hybrid phenotypes depending on the level of hybridization. Moreover, the dominant pattern (savannah state or forest state) of each morphological criterion is unknown, which increases the uncertainty regarding the phenotypes of hybrids.
In our survey we defined the main and secondary criteria and assumed that intermediate states were less informative than the features of forest or savannah elephants, leading to a lower weight being given to the intermediate states in the species assignment key. We made this assumption based on the fact that the intermediate states could be found in both forest and savannah phenotypes and were not restricted to hybrids.
When we compared the results obtained from the species assignment key with those from the K-means analysis, we only obtained a partial match. Although the choice of morphological criteria effectively discriminates between the two elephant species, in all three groups there are individuals with the intermediate phenotype. Our analysis potentially reveals a continuous gradient of phenotypes ranging from the savannah to the forest phenotype via intermediate phenotypes corresponding to hybrid individuals.
The species assignment key could be biased by the ages of individuals. As the forest elephant is a paedomorphic form of the savannah elephant (Debruyne, Reference Debruyne2003), young savannah elephants resemble forest elephants and old forest elephants could be confused with savannah elephants (using features such as ear length and back curvature; Bedetti et al., Reference Bedetti, Greyling, Paul, Blondeau, Clark and Malin2020). Moreover, criteria based on tusks and temporal fossae, which are not yet developed in young individuals (Laws, Reference Laws1966; Sikes, Reference Sikes1966; van der Merwe et al., Reference van der Merwe, Bezuidenhout and Seegers1995), make the assignment of young individuals difficult. Although females have shorter and thinner tusks than males (Elder, Reference Elder1970; Layser & Buss, Reference Layser and Buss1985), sex does not represent a bias here because we did not consider the length and circumference of the tusks.
The IUCN decision to recognize the African elephant as two species aims to reinforce conservation actions, especially for forest elephants, which are Critically Endangered. It also means that we need to gather more information related to hybrids, including number, behaviour and ecology. To be able to carry out effective conservation actions, precise information on the biology, behaviour and reproduction of each species is necessary, which requires being able to distinguish the species of the individuals being studied in areas where the home ranges of forest and savannah elephants overlap. Genetic determination of elephant species is precise, but this process is costly and time consuming. The species assignment key presented here is a simple tool for distinguishing between forest and savannah elephants in environments where both species can be found. However, more studies on the morphology of hybrids are necessary to make the species assignment key more accurate for application in hybridization areas.
Acknowledgements
We thank the Uganda Wildlife Authority and the Uganda National Council for Science and Technology for permission to conduct research in Kibale National Park, Uganda; Jean-Michel Krief, co-director of the Great Ape Conservation Project; Camille Lacroux for the initial checking of the camera trap videos; and all field assistants of the Sebitoli Chimpanzee Project, especially Daniela Zainabu Birungi, Robert Asimwe and Robert Nyakahuma, for the collection and the initial checking of camera trap videos. Financial support for Sebitoli Chimpanzee Project came from Projet pour la Conservation des Grands Singes, Fonds Français pour l'Environnement Mondial, Fondation Prince Albert II, Fondation Nicolas Hulot and Muséum National d'Histoire Naturelle, and the Fondation Ensemble. Julie Bonnald thanks the Muséum National d'Histoire Naturelle, Great Ape Conservation Project and Kinomé for the financial support of her PhD studies and Nicolas Métro and Yohann Fare for the supervision of her work. We thank Frédéric Baer, Jean-François Poudron, Muriel Caslant, Gian-Marco Gesulfo, Camille Lacroux, Nelly Ménard, Sylvie Le Bomin, Shelly Masi, Malenoh Sewuh Ndimbe and Bethan Morgan for providing photographs of elephants; Marion Taylor Baer for editing help; and Régis Debruyne, Martin Fisher and two anonymous reviewers for their insightful comments.
Author contributions
Designing the field programme in Uganda, obtaining the field site, team and authorizations to conduct research: SK, EA; data analysis: JB, RC, MP; analysis, writing: JB, RC, SK; interpretation of results, editing: all authors.
Conflicts of interest
None.
Ethical standards
This research abided by the Oryx guidelines on ethical standards.






 Open access
Open access