



Introduction
The transition from hunting and gathering to farming and herding is one of the most important episodes in human history. Rather than a sudden revolutionary change, subsequently spreading out from a single core zone, this shift was seemingly a slow and complex process, which took place independently in multiple locations across South-west Asia from c. 9500 cal BC (Fuller et al. Reference Fuller, Willcox and Allaby2011; Zeder Reference Zeder2011). In addition, the management, domestication and adoption of different livestock and plant species was highly localised, influenced by both cultural and environmental factors (Fuller et al. Reference Fuller, Willcox and Allaby2011; Arbuckle Reference Arbuckle2014). While the Zagros Mountains—a region that has been subject to greater archaeological investigation than elsewhere in Iran—are considered to be one of the locations where Neolithic lifeways emerged (Weeks Reference Weeks and Potts2013: 56; Vahdati Nasab et al. Reference Vahdati Nasab, Shirvani and Rigaud2019), it is unclear whether hunter-gatherers beyond the Zagros similarly transitioned to farming independently or whether these changes were introduced from elsewhere (Matthews & Fazeli Nashli Reference Matthews and Nashli2022).
The Caspian littoral of north-eastern Iran is an ecologically diverse area, with climatic conditions supporting a broad variety of plant and animal food resources (Leroy et al. Reference Leroy2019), which could have been favourable for a local transition from hunting and gathering to farming. Coon (Reference Coon1951, Reference Coon1952), based on excavations at the cave site of Hotu on the Caspian southern shore, was the first to suggest that animals were domesticated locally in this region. The cave yielded a long sequence of occupation, including Epipalaeolithic and Neolithic layers. Coon claimed to have identified an increase in the quantities of bone from young goat in the late Epipalaeolithic horizons, which he interpreted as evidence for the start of goat husbandry (Coon Reference Coon1951: 50). He also suggested that domesticated sheep, pig, dog and cattle were present in the Neolithic layers of the site (Coon Reference Coon1951, Reference Coon1957) but no analysis substantiating these statements appeared (Harris & Coolidge Reference Harris, Coolidge and Harris2010: 57), since only a selection of the faunal assemblage was published. It therefore remains unclear whether the sheep and goat at Hotu were locally domesticated or imported, or whether introduced domesticates subsequently mixed with local wild animals.
The present study sheds new light on this issue through the analysis of new zooarchaeological data from recent re-excavation of Hotu Cave. The complete zooarchaeological assemblage, as well as other findings from the 2021 excavations, will be published separately.
Regional context
Epipalaeolithic and Neolithic sites on the Caspian littoral located in close proximity to Hotu Cave contain varied zooarchaeological assemblages. The prehistoric layers from Ali Tappeh (for location, see Figure 1) all belong to the Epipalaeolithic, with radiocarbon dates ranging from 11 200 to 9750 cal BC. Gazelle is the most abundant species in all layers, although the relative abundance of gazelle decreases over time and the presence of seals sharply increases (Harris & Coolidge Reference Harris, Coolidge and Harris2010: 55–56). Other animals exploited were onager, wild sheep, aurochs, wild boar, fox and various birds. Sheep and goat numbers peak in the latest two stratigraphic phases of the Epipalaeolithic occupation (McBurney Reference McBurney1969).

Figure 1. Location of Hotu Cave and other Epipalaeolithic and Neolithic sites in the region (figure by A. Richardson).
Komishani Cave and Komishani Tepe are two recently excavated sites on the Caspian shore (see Figure 1). Komishani Cave was highly disturbed and potentially looted, with a mixture of Iron Age and Bronze Age levels, plus Chalcolithic pottery and Neolithic flint tools; an undisturbed Epipalaeolithic layer, however, survived. Radiocarbon dates from this latter horizon range from c. 12 100 to 11 800 cal BC and 10 800 to 10 700 cal BC (Vahdati Nasab et al. Reference Vahdati Nasab2011). Only the animal bones from the Epipalaeolithic layers have been studied, since the analysis of unstratified material would be uninformative. Gazelle is the most abundant taxon, with wild boar, fox and various birds also identified; no sheep/goat and seal bones have been recovered from the site (Mashkour et al. Reference Mashkour, Chahoud and Mahforouzi2010; Vahdati Nasab et al. Reference Vahdati Nasab2020).
Komishani Tepe, situated on the plain in front of Komishani Cave, was excavated in 2017. Its occupation horizons span from the Late Epipalaeolithic into the Early Neolithic and have been dated to between c. 14 350–14 200 and 9200–8200 cal BC (Leroy et al. Reference Leroy2019). The site yielded few animal bones, but sheep and goat are the most abundant species in both the Epipalaeolithic and the Neolithic layers. Wild boar, aurochs, fox and deer are also present, as well as a large variety of water birds, although there is no evidence for the exploitation of marine resources. In terms of species represented, the zooarchaeological assemblage of the Late Epipalaeolithic does not differ from that of the Neolithic, nor does the carbon and nitrogen stable isotope analysis of sheep and goat bones show any notable differences in diet between the Epipalaeolithic and the Neolithic levels (Leroy et al. Reference Leroy2019: SI).
Kamarband Cave (also known as Belt Cave) (Figure 1) was originally excavated at the same time as Hotu Cave (Coon Reference Coon1951, Reference Coon1952). Investigations at Kamarband identified both Epipalaeolithic and Neolithic layers (McBurney Reference McBurney1969); more recent work (see below) has also identified an aceramic Neolithic phase. The Epipalaeolithic layers at Kamarband date to c. 11 350 cal BC, and the Neolithic occupation begins between c. 8000 and 6000 cal BC (McBurney Reference McBurney1969; Leroy et al. Reference Leroy2019). Seal and gazelle are the dominant species during the Epipalaeolithic, but sheep/goat are also present in the later Epipalaeolithic layers, as were aurochs and deer (McBurney Reference McBurney1969). In the Neolithic, sheep/goat are the most abundant species, making up the bulk of the zooarchaeological assemblage (McBurney Reference McBurney1969). New excavations in 2022 at Kamarband Cave, directed by Hassan Fazeli Nashli, have recovered significant quantities of materials, including zooarchaeological remains, from Epipalaeolithic and Early (aceramic) Neolithic levels, which will be the subject of future research.
The changes in species abundance in the zooarchaeological assemblages during the Epipalaeolithic of the region have been attributed to changes in the water levels of the Caspian Sea (McBurney Reference McBurney1969; Leroy et al. Reference Leroy2019), but little is known about the subsequent transition to the Early Neolithic, which is the focus of the present article.
Hotu Cave
Hotu Cave, set in a cliff on the slope of the Alborz Mountains (for location, see Figure 1), is approximately 30 × 20m in area. It was first excavated in 1951 and 1952 over two field seasons, over the course of nine weeks in total; the recording of the finds lacked the rigour expected today (Harris & Coolidge Reference Harris, Coolidge and Harris2010: 57; Leroy et al. Reference Leroy2019: 350) and only part of the faunal assemblage has been published (Coon Reference Coon1951). The site was re-excavated in 2021 by a team directed by Hassan Fazeli Nashli.
The re-excavation of the site identified 124 stratigraphic units, with archaeological layers spanning from the Epipalaeolithic to the Parthian (c. 12 000 BC– AD 300) period (Figure 2). Based on the stratigraphy and archaeological finds, we define six distinct phases (Figure 2). Samples from the Epipalaeolithic and Neolithic layers have been radiocarbon dated, giving a range from c. 12 000 to 6300 cal BC (Table 1). These dates confirm the previous dating of the site, which placed the Epipalaeolithic layers between 12 800 and 8900 cal BC (Harris & Coolidge Reference Harris, Coolidge and Harris2010: 56) and later layers between 7500 and 4000 cal BC (Leroy et al. Reference Leroy2019: 350).

Figure 2. The stratigraphy of Hotu Cave (figure by the authors).
Table 1. New radiocarbon dates from Hotu Cave (calibrated using IntCal20; Reimer et al. Reference Reimer2020).

It is not yet clear whether occupation of the site was continuous through the Epipalaeolithic–Neolithic transition, or whether there was a hiatus in occupation between the Epipalaeolithic and the Early Neolithic. The Early Neolithic is understood to start from Context 99 (between 7 and 6m; see Figure 2), and the radiocarbon dates from Contexts 99 to 88, which overlap, suggest that the site was occupied intensively in the Early Neolithic phase (Table 1). The earliest pottery—Caspian Neolithic Soft Ware—was recovered from Context 75, directly above a layer containing gravel (Context 76, at approximately 5m, marked ‘gap’ on Figure 2) that is interpreted as an episode of cave collapse and abandonment.
Results
Here, we report the faunal remains from the Epipalaeolithic and Early Neolithic layers of Hotu Cave collected during the 2021 field season. The methods used to analyse the assemblage are detailed in the online supplementary information (OSM). The Hotu Cave zooarchaeological assemblage changes significantly across the Epipalaeolithic and Early Neolithic levels (Figure 3; Table 2). The Epipalaeolithic levels are dominated by gazelle, with 64 per cent of the total NISP (Number of Identified Specimens) of mammals, along with a significant presence of seals and large cervids. Sheep and goat appear widely in the assemblage from Context 99 onwards, while gazelle and seals disappear from the assemblage from this stage onwards. This suggests a major shift in animal exploitation practices by the cave's occupants at the start of the Early Neolithic in the early eighth millennium cal BC. Goat outnumbers sheep in both the Early Neolithic and the Pottery Neolithic levels at a ratio of approximately 2:1 (Figure 3; Table 2). Pig, presumably domestic, appear in the later layers of the Pottery Neolithic (Figure 3; Table 2).

Figure 3. NISP (Number of Identified Specimens) of the faunal assemblage from Hotu Cave by period. Ovis/Capra in dark blue represents sheep/goat, blue is sheep, and blue-grey is goat. Among the large cervids, dark blue is any large cervid, blue is red deer (Cervus elaphus) and blue-grey is fallow deer (Dama mesopotamica) (figure by authors).
Table 2. The NISP of the mammals identified, per phase.

The assemblage includes a significant quantity of perinatal remains, not all of which could be identified, due to fragmentation. In addition to these numerous foetal and neonatal bones, both sheep and goat generally died at a young age in the Early Neolithic (Table 3). The small size of the dataset makes it difficult to compare the mortality profiles of sheep and goat, but both species appear only rarely to have reached an age of four or more years. Once both sheep and goat had survived the foetal and neonate stage, however, they generally reached more than one year in age before death. In the Pottery Neolithic, perinatal remains are almost absent (Table 4). Nonetheless, most of the sheep/goat bones are unfused (Table 3).
Table 3. Fusion data of sheep and goat from Early Neolithic Hotu Cave. The sheep/goat category includes sheep, goat and sheep/goat; p. = proximal, d. = distal (following fusion ages of Zeder Reference Zeder and Ruscillo2006).

Table 4. Fusion data from sheep and goat from Pottery Neolithic Hotu Cave. The sheep/goat category includes sheep, goat and sheep/goat; p. = proximal, d. = distal (following fusion ages of Zeder Reference Zeder and Ruscillo2006).

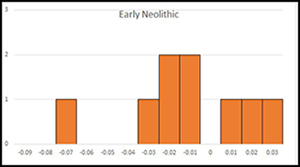
The LSI (Logarithmic Size Index) values of goat in the Early Neolithic show two peaks, which may indicate sexual dimorphism (Figure 4). The graph is slightly skewed, while the kurtosis is negative (Skewness: 0.16, Kurtosis: −0.85), meaning that the mean of the data is greater than the median and the graph is heavy tailed. The dataset from the Pottery Neolithic is smaller and the two peaks are less evident. The skewness of the graph is positive and the kurtosis negative (Skewness: 0.68, Kurtosis: −0.26). The goat bones in the Pottery Neolithic are significantly smaller than those of the Early Neolithic. The LSI of the Early Neolithic sheep shows a flat curve, with a tail at the right (Skewness: −0.52; Kurtosis: 0.44) (Figure 5). Although the number of measurements of Pottery Neolithic sheep is modest, the animals are clearly smaller than the Early Neolithic sheep. The Pottery Neolithic sheep include an outlier on the right-hand side of the graph.

Figure 4. LSI values of goat from Hotu Cave: top) Early Neolithic; centre) Pottery Neolithic; bottom) boxplot of the Early Neolithic (left) and Pottery Neolithic (right), unfused elements in white. The Early Neolithic data include fused elements that fuse before 18 months (n = 15), fused elements that fuse after 18 months (n = 1), elements which do not fuse (astragalus) (n = 5) and unfused elements (n = 3). The Pottery Neolithic includes fused elements that fuse before 18 months (n = 5) and fused elements that fuse after 18 months (n = 2) (for measurements, see Tables S1 & S2 in the OSM) (figure by the authors).

Figure 5. LSI values of the sheep from Hotu Cave: top) Early Neolithic; centre) Pottery Neolithic; bottom) boxplot of the Early Neolithic (left) and Pottery Neolithic (right), unfused elements in white. The Early Neolithic includes elements that do not fuse (astragalus) (n = 9) and fused elements that fuse before 18 months (n = 2). The Pottery Neolithic includes fused elements that fuse before 18 months (n = 3), elements that fuse after 18 months (n = 1), and unfused elements (n = 1) (for measurements, see Tables S3 & S4 in the OSM) (figure by the authors).
Discussion
The absence of sheep/goat in the Epipalaeolithic at Hotu Cave contrasts sharply with their dominance in the faunal assemblage in later levels. At first sight, this pattern might suggest a hiatus in occupation. There is no evidence for a gradual transition towards reliance on sheep and goat, rather than gazelle, cervidae and seal (Figure 3). This shift in the zooarchaeological assemblage could indicate that a decline in the availability of gazelle, for some reason, led local populations to exploit sheep/goat instead. Both wild sheep and wild goat were present in the region during the Epipalaeolithic, as attested at Komishani Tepe and Kamarband Cave, and wild sheep were present at Ali Tappeh and in small numbers at Hotu Cave. It is therefore likely that these animals were also available in the Early Neolithic. Another possibility is that incoming groups brought domesticates with them, or managed animals in some form. One possible scenario is that settlers from the Zagros region of western Iran, where Neolithic developments commenced at a very early date (e.g. management of goat from c. 8000 cal BC), travelled eastwards along the Great Khorasan Road on the northern edge of the central plateau of Iran and introduced Neolithic lifeways (Harris & Coolidge Reference Harris, Coolidge and Harris2010; Roustaei & Gratuze Reference Roustaei and Gratuze2020; Matthews & Fazeli Nashli Reference Matthews and Nashli2022). The presence of domesticated sheep at Obishir in southern Kyrgyzstan by 6000 cal BC (Taylor et al. Reference Taylor2021) suggests a relatively rapid spread of Neolithic herding practices eastwards across north-eastern Iran and southern Turkmenistan. Intriguingly, aDNA from a human individual from Hotu Cave, found during the Coon excavations possibly in an Epipalaeolithic layer, shows genetic affinity with western Iranian farmers (Lazaridis et al. Reference Lazaridis2016).
The sexual dimorphism among mature goats, shown by the two peaks in Figure 4, indicates that both male and female goats were kept or consumed at Hotu Cave. Since bones could all derive from relatively young or mature animals, it is not possible to determine whether male animals were slaughtered earlier than the females. In addition to the perinatal remains, all of the goats were slaughtered at a young age during the Early Neolithic (Table 3). This mortality profile is very different from that of a hunted population, where a focus on prime adults would be expected. The sample size of the Pottery Neolithic is small but is more suggestive of a young male cull (Figure 4). As for the graph of sheep in the Early Neolithic (Figure 5), it seems to be dominated by males. This might represent a focus on prime adults, but the ageing pattern does not endorse such an interpretation (Table 3). In the Pottery Neolithic, a large percentage of the sheep and goat population continues to be slaughtered at a young age, while the percentage of perinatal remains is only four per cent (Table 4). The sample size of the Pottery Neolithic is smaller, but both the sheep and goat LSI values clearly decrease (Figures 4 & 5). It is possible that this is the result of a decrease in the body size in animals or that smaller animals were introduced, but it could also reflect a change in management practices. The large number of young animals in the assemblage could indicate that males were often culled before they were 18 months old (Table 4). A similar size decrease in both sheep and goat has been observed in the Zagros region in the Pottery Neolithic, although the reason for this is unknown (Zeder Reference Zeder, Vila, Gourichon, Choyke and Buitenhuis2008).
The numerous perinatal remains of sheep/goat in the Early Neolithic at Hotu (30 per cent) indicates that they were kept at the cave or close to it, which suggests that people were not solely hunting sheep/goat but also controlled their movement. Caves were used as places to pen sheep and goat at various Neolithic sites in Europe (e.g. in the northern Adriatic basin: Miracle Reference Miracle, Serjeantson and Field2006; or at El Mirador in Spain: Martín et al. Reference Martín, García-González, Nadal and Vergès2016), and frequently the bones of very young caprines have been found in such cave deposits (e.g. at Franchthi Cave in Greece: Munro & Stiner Reference Munro and Stiner2015; in cave sites of the northern Adriatic: Boschin Reference Boschin2020; or at Els Trocs Cave in Spain: Tejedor-Rodríguez et al. Reference Tejedor-Rodríguez2021). It has been suggested that caves were used as locations for breeding and flock management; today, pastoralists often separate pregnant animals from the rest of the flock to give them extra care (Martín et al. Reference Martín, García-González, Nadal and Vergès2016). It is uncertain whether all sheep/goat were kept in or close to Hotu Cave during the Early Neolithic, or whether the cave was perhaps used as shelter for pregnant animals. It is thus possible that, rather than a change in diet, we are witnessing a change in the function of the cave.
The large number of foetal animals and neonates could be related to a sub-group of animals being kept in the cave. The ratio of spontaneous abortions in present-day sheep and goat flocks varies. A survey among sheep/goat farmers in Jordan indicated a perinatal mortality in sheep of 7.5 per cent and 13 per cent in goats (Aldomy et al. Reference Aldomy2009). In cases of infectious disease, this figure can increase to 20–40 per cent (Martín et al. Reference Martín, García-González, Nadal and Vergès2016: 324). Non-infectious causes of spontaneous abortions and stillbirths in sheep and goat can have various causes, such as trauma, which can be brought on by multiple factors such as long journeys, unfamiliar accommodation, climatic changes, carnivore attacks, nutritional deficiencies or the ingestion of toxic plants (Aldomy et al. Reference Aldomy2009; Martín et al. Reference Martín, García-González, Nadal and Vergès2016). Multiple infectious diseases—some also infectious to humans—can lead to stillbirths and abortion (Aldomy et al. Reference Aldomy2009; Fournié et al. Reference Fournié, Pfeiffer and Bendrey2017). Zoonotic diseases are likely to have increased during the Early Neolithic because humans brought animals together in larger, denser herds, leading to increased contact with the animals (Fournié et al. Reference Fournié, Pfeiffer and Bendrey2017). A high proportion of abortions, stillbirths and neonatal deaths might also be due to inexperience or a community experimenting with sheep/goat management. The community at Hotu Cave possibly managed both sheep and goat in the Early Neolithic. Based on bone morphology, it is impossible to make a distinction between foetal sheep and goat, and rarely possible for young animals (Martín & García-González Reference Martín and García-González2015). The assemblage of Hotu Cave is too small to make solid comparisons between the mortality profiles of sheep and goat, but both species were, in general, young at the age of death (Tables 3 & 4), indicating that both were managed. It is common for modern-day herders in the region to keep sheep and goat in the same flock (Ansari-Renani et al. Reference Ansari-Renani2013; Elliot et al. Reference Elliott2015).
Currently, there are no comparable faunal datasets from contemporaneous sites in the region in terms of mortality profiles and animal size through which to gain a better understanding of regional sheep/goat management practices. At Komishani Tepe, both sheep and goat were present in Epipalaeolithic and Neolithic levels, but the assemblage is too small to reconstruct mortality profiles or size patterns (Leroy et al. Reference Leroy2019).
Conclusions
The archaeozoological data from Hotu Cave convincingly demonstrate, for the first time, that sheep/goat management was practised during the Early Neolithic along the Caspian littoral. The cave's faunal assemblage changes dramatically from a predominance of gazelle in the Epipalaeolithic to an almost complete dominance of sheep/goat in the Early Neolithic. The cave's stratigraphy suggests that there was a hiatus in occupation between the Early Neolithic and the Pottery Neolithic, but not between the Epipalaeolithic and the Early Neolithic. Relatively little is currently known about the transition to the Neolithic in Iran beyond the Zagros region; specifically, it is unknown whether domesticated animals and plants were spread across the plateau from the Zagros, or whether there were multiple local centres of domestication. Our analysis suggests that sheep and goat could have been domesticated locally. It is, however, also possible that this switch in the zooarchaeological profile indicates a hiatus in occupation between the Epipalaeolithic and the Neolithic levels of Hotu Cave, with a new population from the west bringing domesticated sheep/goat with it. Further research must therefore be conducted to establish whether local domestication did indeed occur in the region.
The large proportion of perinatal remains of sheep/goat found in the Early Neolithic layers of Hotu Cave indicates that humans kept sheep and goat in or close to the cave. Given that unfused young bones of both species of animal were present, it is certainly possible that sheep and goat were managed during the Early Neolithic, as caves were used to pen herds during the Neolithic in various parts of the world. Further, it is possible that Hotu Cave was used as a shelter for pregnant and young animals. This would explain the presence of large numbers of perinatal remains, which, in turn, could be related to an increase in zoonotic diseases.
Both sheep and goat are also present in the Pottery Neolithic layers of Hotu Cave and the zooarchaeological data indicate that they were closely managed. There is more evidence for a traditional management pattern in the form of a young male cull of both sheep and goat in the Pottery Neolithic, but the sample size is limited. The clear decrease in size in both sheep and goat in the Pottery Neolithic compared with the Early Neolithic might indicate incoming groups, bringing smaller domesticated animals with them; alternatively, it could also be the result of a decrease in sexual dimorphism under the influence of domestication.
Future research, which could shed further light on the Early Neolithic along the Caspian littoral, should include enhanced identification of sheep versus goat using ZooMS (Zooarchaeology by Mass Spectrometry; Pilaar Birch et al. Reference Pilaar Birch, Scheu, Buckley and Çakırlar2019), the determination of the geographical origin of animals and their dietary histories through stable isotope analysis, and the analysis of the aDNA of individual sheep or goats from the Hotu Cave zooarchaeological assemblage, within the wider context of Iran and beyond. Additional radiocarbon dates may also illuminate the development from the Epipalaeolithic to the Neolithic at Hotu Cave. Finally, the study of the faunal remains and other materials from Hotu and Kamarband caves shows great potential in terms of understanding the critical issue of local Neolithisation processes versus the introduction from elsewhere of well-developed animal herding practices.
Acknowledgements
We are grateful to Melinda Zeder for her comments on this article and to the anonymous peer reviewers for their insights.
Funding statement
This project has received funding from the European Research Council under the European Union's Horizon 2020 research and innovation programme (grant agreement ERC AdG 787264), 2018–2024, for the project MENTICA (Middle East Neolithic Transition: Integrated Community Approaches).
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.15184/aqy.2023.37.










 Open access
Open access