



Introduction
Palmer amaranth (Amaranthus palmeri S. Watson) is one of the most common and difficult to control weeds in many crops, including soybean [Glycine max (L.) Merr.], due to a range of ecophysiological (Korres et al. Reference Korres, Norsworthy, FitzSimons, Roberts and Oosterhuis2017b; Massinga et al. Reference Massinga, Currie and Trooien2003) and biological characteristics (Korres and Norsworthy Reference Korres and Norsworthy2017) that enhance the adaptive ability of the weed (Korres et al. Reference Korres, Norsworthy, Brye, Vaughn and Mauromoustakos2017a) in a wide range of environments (Korres et al. Reference Korres, Norsworthy, Bagavathiannan and Mauromoustakos2015a, Reference Korres, Norsworthy, Bagavathiannan and Mauromoustakos2015b, Reference Korres, Norsworthy, Young, Reynolds, Johnson, Conley, Smeda, Mueller, Spaunhorst, Gage, Loux, Kruger and Bagavathiannan2018). Furthermore, the continuous establishment of A. palmeri populations throughout the growing season enhances the adaptive and competitive ability of the weed (Bensch et al. Reference Bensch, Horak and Peterson2003; Korres et al. Reference Korres, Norsworthy and Mauromoustakos2019a). It has been shown that early A. palmeri establishment can cause significant soybean yield decreases of between 19% and 80% (Bensch et al. Reference Bensch, Horak and Peterson2003; Korres et al. Reference Korres, Norsworthy and Mauromoustakos2019a).
Manipulation of soybean density can counteract the competitive ability of the weed through canopy closure (Korres et al. Reference Korres, Reddy, Rouse, King, Korres, Burgos and Duke2019b), which results in reduced light transmission to the soil surface (Bell et al. Reference Bell, Norsworthy and Scott2015; Korres and Norsworthy Reference Korres and Norsworthy2017). Low light conditions reduce A. palmeri biomass production, leaf number, specific leaf area, photosynthetic capacity (Korres et al. Reference Korres, Norsworthy, FitzSimons, Roberts and Oosterhuis2017b), and density (Jha and Norsworthy Reference Jha and Norsworthy2009). Harder et al. (Reference Harder, Sprague and Renner2007) reported that soybean densities of 124,000 to 198,000 plants ha−1 had no effect on weed biomass, whereas a 20% biomass reduction was observed at densities of 300,000 to 445,000 plants ha−1 compared with low soybean densities.
Nevertheless, the benefits associated with increased crop competitiveness as a result of increased crop density, especially after a density threshold, which depends on the crop and cropping system, are debatable and must be evaluated based on yield increases, economic returns, and long-term weed management benefits (DeWerff et al. Reference DeWerff, Conley, Colquhoun and Davis2014; Harder et al. Reference Harder, Sprague and Renner2007; Harker et al. Reference Harker, Clayton, Blackshaw, O’Donovan and Stevenson2003). Soybean yield is related to crop density (Fickett et al. Reference Fickett, Boerboom and Stoltenberg2013; Nave and Wax Reference Nave and Wax1971), although compensatory effects in combination with environmental conditions impose a wide range of crop yield responses to density manipulation (Benbella and Paulsen Reference Benbella and Paulsen1998). Norsworthy and Oliver (Reference Norsworthy and Oliver2001) stated that seed cost associated with high crop densities (i.e., >450,000 plants ha−1) can exceed the benefit for better weed control. Bell et al. (Reference Bell, Norsworthy and Scott2015) found that weed control and soybean yield were greater at soybean seeding rates equal to 617,500 seed ha−1 (average density achieved was 400,000 to 580,000 plants ha−1) than at lower soybean densities ranging between 78,000 to 250,000 plants ha−1.
Profitability of agricultural production systems is a function of commodity prices, yield, and cost of production. Increased seed rates, particularly in genetically modified (GM) soybean systems, could increase the production cost significantly due to high seed cost (Place et al. Reference Place, Reberg-Horton, Dunphy and Smith2009; Thompson et al. Reference Thompson, Larson, Lambert, Roberts, Mengistu, Bellaloui and Walker2015). High prices for the seed exert a direct impact on farmers’ decisions related to the range of allowable seeding rates (Epler and Staggenborg Reference Epler and Staggenborg2008; Shi et al. Reference Shi, Chavas and Stiegert2010; Thompson et al. Reference Thompson, Larson, Lambert, Roberts, Mengistu, Bellaloui and Walker2015). Therefore, it is vital to address the effects of increased soybean density in association with soybean grain yield outcome (i.e., the cost of the production system) along with the weed suppression. If weed suppression due to increased crop density does not coincide with yield increases and, consequently, profit margin improvements, then further consideration of this option as a feasible IWM option to control A. palmeri is warranted. The evaluation of cropping systems, for example, as crop density increases, can be facilitated by crop budgeting, a management tool used to estimate costs and evaluate cropping alternatives (Anonymous 2019b).
Therefore, this research aims to determine the extent to which increasing soybean plant density suppresses A. palmeri plants established at various periods of the growing season and to investigate the response of soybean yield to a range of A. palmeri establishment timings. We tested the following hypotheses: (1) whether increasing soybean density reduces the growth and seed production of early-establishing A. palmeri populations; 2) whether increasing soybean density is accompanied with greater yield up to a plateau, at which further yield increases cease to occur; (3) whether increasing soybean density is unable to delimit crop dry weight, pods per plant, and yield reductions caused by early-establishing A. palmeri populations; and (4) whether increasing soybean density compensates for greater production cost compared with lower crop densities. In addition, the differentiation of A. palmeri gender at a range of crop densities was investigated based on height and biomass production.
Materials and Methods
Experimental Setup and Study Site
Two field trials conducted during 2014 and 2015 at the University of Arkansas, Fayetteville, AR (36.095°N, 94.172°W) on Captina silt-loam soil (fine-silt, mixed, active, thermic Typic Albaquults) with pH of 6.7, organic matter 1.5%, and sand, silt, and clay content of 34%, 53%, and 13%, respectively. A four by six factorial experiment was arranged in a randomized complete block design with four replications. The soybean density factor consisted of four treatments: 0 (weedy monoculture), 125,000 or “low” density, 250,000 or “medium” density, and 400,000 or “high” density seeds ha−1. The A. palmeri establishment timing factor, measured in weeks after soybean emergence (WAE), consisted of six treatments: 0, 1, 2, 4, 6, and 8 WAE. The harvestable plot size was 12 m2 (6-m long by 2-m wide), whereas a predetermined plot (9-m long by 2-m wide) outside the harvested area was used for crop destructive sampling throughout the growing season.
Disking followed by a field cultivator (Kongskilde Industries, Hudson, IL, USA) was used on the entire experimental area to prepare the seedbed before soybean planting. Phosphorous and potassium fertilizer (0-40-60) (Slaton et al. Reference Slaton, DeLong, Mozaffari, Shafer, Branson and Richards2005) was applied shortly before planting, assuming that crop removal of phosphorous equals 42 kg P2O5 ha−1 and crop removal of potassium equals 69 kg K2O ha−1 (Anonymous 2019a, 2019b; Flanders Reference Flanders2014; Place et al. Reference Place, Reberg-Horton, Dunphy and Smith2009; Plastina Reference Plastina2019; Schnitkey Reference Schnitkey2018). A glufosinate-resistant (LibertyLink®) soybean cultivar (‘Pioneer® 95L01’, maturity group 4.6, DuPont, Leland, MS, USA) was then seeded in a 10-row plot (20-cm row spacing) using an Almaco cone-drill planter (Almaco, Nevada, IA, USA). Insecticide as zeta-cypermethrin active ingredient was applied at the recommended rate for control of green stink bug (Chinavia hilaris Say) at R2-R3 soybean growth stage (GS).
Experimental plots were hand weeded on a weekly basis for the first 2 mo and regularly afterward to remove unwanted weeds and were irrigated using an overhead sprinkler irrigation system (Valmont Industries, Valley, NE, USA) when rainfall did not occur for an approximately 10-d period, to avoid drought stress.
Plant Material and Experimental Treatments
Amaranthus palmeri seeds were collected in the 2013 to 2014 growing season from a local population at the University of Arkansas farm, Fayetteville, AR, and were stored in sealed vials at 5 C until their use. The germination of A. palmeri seedling grown at 14-h photoperiod and 35/23 C day/night temperature under greenhouse conditions was planned to coincide at 0, 1, 2, 4, 6, and 8 wk in relation to soybean emergence. Sixteen randomly selected plots (i.e., four crop densities by four replications) were used for each A. palmeri establishment timing until all experimental treatments were completed. Amaranthus palmeri seedlings at the 2- to 4-leaf stage were randomly transplanted in the plot area (excluding the edge rows of the plot) targeting a density of 1 A. palmeri plant m−2. Watering the young A. palmeri plants every 2 to 3 d for a 2- to 3-wk period minimized possible stress during their acclimatization period. The entire process of A. palmeri germination and transplanting for each establishment time has been described in detail by Korres et al. (Reference Korres, Norsworthy and Mauromoustakos2019a).
Data Collection
Soybean crop establishment for each seeding rate was evaluated at harvest to calculate the final crop density using a 1-m2 quadrat at three randomly selected sampling points within each experimental plot. In addition, five randomly selected soybean plants from predetermined areas for destructive sampling were collected at R2-R3 and R6 soybean GS, and biomass production (at R2-R3 and R6 soybean GS) and pods per plant (at R6 soybean GS) were recorded after drying the plants at 70 C for approximately a week until no further dry weight reduction was observed.
Soybean grain was harvested with a small-plot combine; yield was adjusted to 13% moisture and recorded (in kg ha−1). Before soybean harvesting, all A. palmeri plants, both male and female, were collected from each plot by cutting the stems at the soil level, and the height of each A. palmeri plant was recorded. The plants were then placed in paper bags and dried at 70 C for approximately a week until no further dry weight reduction was observed. Amaranthus palmeri biomass production was recorded and seeds from each female plant were estimated as described by Korres et al. (Reference Korres, Norsworthy and Mauromoustakos2019a).
In addition, ground cover, leaf area index (LAI), and light interception measurements were recorded throughout the growing period. More particularly, crop canopy photographs for each A. palmeri establishment time along with light interception by the crop canopy and LAI measurements were obtained immediately before each transplanting treatment, and only from the plots designated to that particular A. palmeri transplanting treatment, as described by Korres et al. (Reference Korres, Norsworthy and Mauromoustakos2019a). Digital imagery, a technique reported to be an accurate approach to monitor crop canopy formation (Purcell Reference Purcell2000) was used to estimate ground cover. The Monsi-Saeki extinction coefficient (hereafter called “extinction coefficient”), based on light interception and LAI, was estimated based on Equation 1 (Monsi and Saeki Reference Monsi and Saeki1953):
 $$I = {I_0}{e^{ - kL}}$$
$$I = {I_0}{e^{ - kL}}$$
where k is the extinction coefficient, I 0 is the light intensity above the crop canopy, I is the light intensity below the crop canopy, and L is the LAI of soybean leaves causing the light attenuation. The extinction coefficient relates crop leaf area and canopy architecture with light interception by crop canopy, hence shading conditions beneath crop canopy where the weed is growing.
Data Analysis
A two-parameter exponential decay model was employed to analyze A. palmeri dry weight and seed production, whereas a single linear regression analysis was best fit on A. palmeri height. In addition, a correlation analysis was used to identify the relationships between ground cover and extinction coefficient for each crop density used. Finally, an ANOVA was used to compare A. palmeri dry weight between male and female A. palmeri plants, soybean dry weight, pods per plant, and yield. An LSD test was used for means separation. The employment of ANOVA for the analysis of soybean yield facilitated the crop budget analysis described in the following section. All data analyses were performed using JMP Pro v. 14.0.0 software (SAS Institute, Cary, NC, USA). Values from SigmaScan Pro were exported to SigmaPlot v. 13.0 (Systat Software, CA, USA) to examine the correlation between groundcover and extinction coefficient.
Crop Budget Analysis
In addition to the analysis of the experimental data, a crop budget analysis was performed to evaluate the profitability of the cropping systems under investigation. Soybean production inputs typical to current production systems, that is, fixed and variable costs for preharvest machinery (i.e., moldboard plow, disk/field cultivator, and direct drill), crop husbandry (i.e., seed, fertilizing, irrigation, and crop protection), labor (mechanical weed control, insecticide application, irrigation, and fertilizing), and harvesting and grain storage (i.e., combine and storage) were balanced against crop revenue, net return, and breakeven price for each soybean seeding rate used for both 2014 and 2015. Fixed and variable costs for preharvest machinery were estimated at US$51.38 ha−1 and US$48.66 ha−1, respectively, totaling US$100.04 ha−1 for both 2014 and 2015. Fixed and variable costs for crop husbandry were estimated at US$192.66 ha−1 and US$162.66 ha−1 for low crop density (125,000 seeds ha−1); US$192.66 ha−1 and US$207.69 ha−1 for medium (250,000 seeds ha−1) crop density; and US$192.66 ha−1 and 261.69 ha−1 for high (400,000 seeds ha−1) crop density. The average soybean seed price for LibertyLink® soybean is US$54 per 150,000 seed-containing packages, which also includes a technology fee (Anonymous 2019a; Thompson et al. Reference Thompson, Larson, Lambert, Roberts, Mengistu, Bellaloui and Walker2015). Seeding rates were the same for both experimentation years, with the fixed cost (i.e., US$45, US$90, and US$144 ha−1 for 125,000, 250,000, and 400,000 seeds ha−1, respectively). Variable costs for phosphorus and potassium fertilizer and irrigation fuel were estimated at US$20.3, US$22.35, and US$78 per unit of fertilizer (Anonymous 2019a, 2019b; Flanders Reference Flanders2014; Place et al. Reference Place, Reberg-Horton, Dunphy and Smith2009; Plastina Reference Plastina2019; Schnitkey Reference Schnitkey2018). Labor was estimated at US$14.5 h−1 (Place et al. Reference Place, Reberg-Horton, Dunphy and Smith2009; Plastina Reference Plastina2019). Finally, fixed and variable costs for harvesting and grain storage operations were equal to US$23.24 and US$12.45 ha−1 for low crop density; US$23.81 and 12.97 ha−1 for medium crop density; and US$23.95 and US$13.11 ha−1 for high crop density (Anonymous 2019a; Flanders Reference Flanders2014; Place et al. Reference Place, Reberg-Horton, Dunphy and Smith2009; Plastina Reference Plastina2019; Schnitkey Reference Schnitkey2018). Final grain yield and the total production costs (fixed and variable) were used for the determination of total expenses, breakeven point, and total market revenue for each soybean seeding rate used (Table 1). Inputs in this analysis were held constant, except those associated with seeding rates. Crop revenue under various seeding rates was determined using annual average soybean prices between 2010 to 2018 (USDA-NASS 2019b, 2019c; World Bank 2019); the mean soybean price was estimated at US$0.39 kg−1 (approximately US$10.6 bu−1 of soybean grain). In addition, a budget analysis was developed based on A. palmeri establishment timings and the corresponding soybean yields (Table 2). Because no interaction was recorded between A. palmeri establishment timing and soybean density, the average value of production costs (i.e., fixed and variable) across soybean seeding rates (i.e., 125,000, 250,000, and 400,000 seeds ha−1) from Table 1 was considered for the estimation of total expenses, breakeven prices, crop revenue, and net return depicted in Table 2. Inputs in this analysis were held constant, except those associated with soybean yields at 0, 1, 2, 4, 6, and 8 WAE A. palmeri establishment timings.
Table 1. Effects of soybean seeding rate on total expenses, breakeven price, crop revenue, and net revenue for 2014 and 2015.a

a U.S. soybean grain price (average marketing price 2010–2018) = US$0.39 kg−1 (USDA-NASS 2019b, 2019c; World Bank 2019). All calculations are based on Pendell et al. (Reference Pendell, Perry, Williams, Barnaby and Lubben2003) and Thompson et al. (Reference Thompson, Larson, Lambert, Roberts, Mengistu, Bellaloui and Walker2015).
b Values with the same letter in yield column are not different at P < 0.05.
c Breakeven price = Total crop expenses (US$ ha−1)/soybean grain yield (kg ha−1).
d Crop revenue = Yield (kg ha−1) × soybean grain price (US$ kg−1).
e Net return = Crop revenue − total expenses (US$ ha−1).
Table 2. Effects of Amaranthus palmeri establishment time on total expenses, breakeven price, crop revenue, and net revenue for 2014 and 2015.a
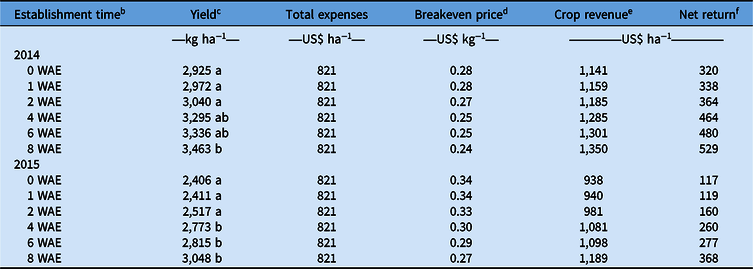
a U.S. soybean grain price (average marketing price 2010–2018) = US$0.39 kg−1 (USDA-NASS 2019b, 2019c; World Bank 2019). All calculations are based on Pendell et al. (Reference Pendell, Perry, Williams, Barnaby and Lubben2003) and Thompson et al. (Reference Thompson, Larson, Lambert, Roberts, Mengistu, Bellaloui and Walker2015).
b Establishment time expressed in weeks after emergence (WAE).
c Values with the same letter in yield column are not different at P < 0.05.
d Breakeven price = Total crop expenses (US$ ha−1)/soybean grain yield (kg ha−1).
e Crop revenue = Yield (kg ha−1) × soybean grain price (US$ kg−1).
f Net return = Crop revenue − total expenses (US$ ha−1).
Results and Discussion
Site Specifications
The 2014 growing season was characterized by more erratically distributed precipitation compared with the 2015 growing season. In 2015 the monthly precipitation was 125% higher than in 2014 and 55% higher than the 30-yr average. The average monthly precipitation between June and July was recorded at 10.2 and 6.1 mm compared with 17 and 20 mm for the years 2014 and 2015 respectively.
Soybean achieved densities in 2014 were 121,300 (±478.7 standard error of mean [SE]), 242,600 (±629.1 SE) and 389,300 (±1,417.6 SE) plants ha−1, representing 97% of the 125,000, 250,000, and 400,000 seeds ha−1 targeted seeding rates respectively. Soybean densities in 2015 were underachieved owing to severe weather conditions. More specifically, for a targeted crop density of 125,000 plants ha−1, the achieved density was 98,500 (±1,121.7 SE), for 250,000 the achieved density was 198,200 (±2,178.1 SE), and for targeted crop density 400,000 plants ha−1 the achieved density was recorded at 323,200 (±4,333.7 SE) plants ha−1.
Does Soybean Density Similarly Affect Growth and Seed Production of Early- and Late-Established Amaranthus palmeri Populations?
The effects of soybean density on A. palmeri biological characteristics became apparent for the weed plants established at 2 WAE establishment timing onward. Great crop density, for example, caused A. palmeri height reductions in comparison to the A. palmeri height in weedy monoculture, particularly at 1 and 2 WAE establishment timings. When A. palmeri establishment time coincided with crop emergence (i.e., 0 or 1 WAE establishment timing), the competitive effects of the crop at low densities almost ceased, as the height of the weed was no different compared with the height of A. palmeri growing under crop-free conditions (Figure 1; Supplemental Table 1).

Figure 1. Effects of soybean density by Amaranthus palmeri establishment time on A. palmeri height (recorded at harvest) in weeks after emergence of the weed relative to soybean emergence.
The effects of soybean density on weed dry weight production were assessed at crop harvest. Significant reductions were recorded when the biomass produced in weedy monoculture was compared with that produced under crop competition, especially on biomass of early A. palmeri establishment (Figure 2; Supplemental Table 2).

Figure 2. Effects of soybean density by Amaranthus palmeri establishment time on A. palmeri dry weight (recorded at harvest) in weeks after emergence of the weed relative to soybean emergence.
The greater the crop density, the greater the reduction of A. palmeri biomass, particularly at early establishment timings of the weed (i.e., 0 and 1 WAE establishment timings). Biomass produced at late establishment times (i.e., 4 to 8 WAE establishment timings) was affected less by crop interference, irrespective of crop density, due to shorter season. It is known that the establishment time affects the size of the plant, because the transition time from vegetative to reproductive growth is shorter, especially for plants like A. palmeri that flower in response to photoperiod (Goyne and Schneiter Reference Goyne and Schneiter1988). Therefore, preventing early-establishing A. palmeri cohorts and enhancing crop competitiveness that results in reducing biomass production would have a direct effect on fecundity of the weed. Korres and Norsworthy (Reference Korres and Norsworthy2017) and Korres et al. (Reference Korres, Norsworthy and Mauromoustakos2019a) have reported the association of biomass and seed production in A. palmeri.
Amaranthus palmeri seed production exhibited a pattern similar to biomass, particularly at the early establishment timings of the weed. Significant seed production reductions were recorded at 0 and 1 WAE establishment timings compared with seed produced in the absence of the crop (Figure 3; Supplemental Table 3). No differences in A. palmeri seed production were recorded at 4, 6, and 8 WAE establishment timings, possibly due to late establishment timing of the weed. Nevertheless, A. palmeri plants at late establishment timings were capable, even when growing with soybean, to produce 60 to approximately 3,500 seeds plant−1 (Figure 3; Supplemental Table 3).

Figure 3. Effects of soybean density by Amaranthus palmeri establishment time on A. palmeri seed production (recorded at harvest) in weeks after emergence of the weed relative to soybean emergence.
Increased crop densities accelerate canopy closure and reduce the amount of light penetrating the canopy and reaching the soil surface, hence lessening weed growth and biomass accumulation beneath the crop canopy (Kudsk et al. Reference Kudsk, Vasileiadis, Sattin, Korres, Burgos and Duke2019). Indeed, the percentage ground cover and the extinction coefficient were greater under great crop density than at low crop density (Figure 4).

Figure 4. Effects of ground cover and extinction coefficient of light interception at low, medium, and high crop densities. Ground cover, leaf area, and light interception were measured immediately before each Amaranthus palmeri establishment time. Note: Average low density achieved for 2014 was 121,302 plants ha−1 (120,311, 122,292) and for 2015 was 98,437 plants ha−1 (96,117, 100,758); average medium density achieved for 2014 was 242,604 plants ha−1 (241,302, 243,906) and for 2015 was 198229 plants ha−1 (193,724, 202,735); and average high density achieved for 2014 was 389,333 plants ha−1 (386,401, 392,266) and for 2015 was 323,167 plants ha−1 (314,202, 332,132). Numbers in parentheses indicate the lower and upper 95% means, respectively. The additional x axis at the bottom of the graph approximates ground cover using A. palmeri establishment time in weeks after crop emergence (WAE). Arrows indicate the corresponding line for each regression equation provided in the graph.
It can therefore be concluded that greater soybean density suppresses growth and seed production of early-establishing A. palmeri. In contrast, greater soybean density offers no suppression advantages for late-establishing A. palmeri. The growth plasticity of late-establishing A. palmeri (i.e., 6 and 8 WAE establishment timings) at greater soybean densities is noticeable. These late-establishing plants were taller compared with A. palmeri plants established at 2 or 4 WAE establishment timing (Figure 1) when ground cover was between 40% and 60% (Figure 4).
Differential Performance between Amaranthus palmeri Gender
There was greater biomass production (P2014, 2015 < 0.001) by the female A. palmeri plants at early establishment timings (i.e., 0 and 1 WAE establishment timings) compared with biomass produced by the male A. palmeri plants at the same establishment timings (Figure 5). These differences dissipated as crop density increased in relation to increases of the establishment timing interval from soybean emergence, except for A. palmeri plants that were grown under crop-free conditions. This trend occurred in both 2014 and 2015 (Figure 5). Differences in height between A. palmeri male and female plants were not found in this work (data not shown).

Figure 5. Differential performance of female and male Amaranthus palmeri plants at different establishment times (0, 1, 2, 4, 6, and 8 weeks after crop emergence [WAE]) and crop densities. Vertical bars represent LSD values for mean separation at P < 0.001. Note: Average low density achieved for 2014 was 121,302 plants ha−1 (120,311, 122,292) and for 2015 was 98,437 plants ha−1 (96,117, 100,758); average medium density achieved for 2014 was 242,604 plants ha−1 (241,302, 243,906) and for 2015 was 198,229 plants ha−1 (193,724, 202,735); and average high density achieved for 2014 was 389,333 plants ha−1 (386,401, 392,266) and for 2015 was 323,167 plants ha−1 (314,202, 332,132). Numbers in parentheses indicated the lower and upper 95% means, respectively.
Nevertheless, differences in height and biomass production between A. palmeri genders have been previously reported (Keeley et al. Reference Keeley, Carter and Thullen1987; Korres et al. Reference Korres, Norsworthy, FitzSimons, Roberts and Oosterhuis2017b; Webster and Grey Reference Webster and Grey2015). The differential response in size between female and male A. palmeri plants might contribute to variation in fitness (Solbrig Reference Solbrig1981), which may convey an adaptive plasticity that is imposed by the effort of the female plants for reproduction (Korres et al. Reference Korres, Norsworthy, FitzSimons, Roberts and Oosterhuis2017b; Obeso Reference Obeso2002).
Effects of Crop Density on Soybean Yield
The lower number of pods per plant at higher crop densities due to different impacts of intraspecific competition on the crop (Yamada et al. Reference Yamada, Sasakura, Nishiwaki, El-Shemy, Mohapatra, Nguyen, Kurosaki, Kanai, Ito and Fujita2011) did not affect yield per unit area (Table 1), with yield increases (P2014, 2015 < 0.05) at medium and high crop densities observed compared with low crop density, in agreement with French (Reference French, Wrigley, Corke and Walker2004).
The greater the crop density, the greater the grain yield (P2014, 2015 < 0.05) averaged across A. palmeri establishment timings. However, grain yield was no different between medium and high crop densities in both years (Table 1). As stated by Board (Reference Board2000), Kane and Grabau (Reference Kane and Grabau1992), and Weaver et al. (Reference Weaver, Akridge and Thomas1991), a positive correlation between grain yield and crop density is not always evident in soybean. Murdoch (Reference Murdoch, Korres, Burgos and Duke2019) reported that one problem with increasing seed rate is that intraspecific competition among crop plants may increase due to the increase in rectangularity of the crop.
Increasing Soybean Density Cannot Diminish Crop Yield Reductions Caused by Early Amaranthus palmeri Populations
Early A. palmeri establishment timings reduced soybean biomass production, an effect that was observed at R2-R3 and R6 crop GS. Crop biomass reductions (P2014, 2015 < 0.05) due to competition by the early-established A. palmeri plants were recorded at R2-R3 and R6 crop GS or 8 and 12 WAE irrespective of crop density (Supplemental Figure 1), indicating the importance of weed control at the early soybean growth stages (Hartzler and Battles Reference Hartzler and Battles2004; Korres et al. Reference Korres, Norsworthy and Mauromoustakos2019a; VanAcker et al. Reference VanAcker, Swanton and Weise1993).
Nevertheless, soybean response to A. palmeri competition in terms of biomass production was determined by the establishment time of the weed. Greater soybean biomass production at late A. palmeri establishment timings, that is, 6 and 8 WAE, resulted in more (P2014, 2015 < 0.05) pods per plant (Supplemental Figure 2), especially at lower crop density. In medium or high crop densities, the number of pods per plant was no different.
Establishment time of Amaranthus spp. regulates the extent of competition with crops (Korres et al. Reference Korres, Norsworthy and Mauromoustakos2019a). Soybean yield was significantly (P2014, 2015 < 0.05) affected by the A. palmeri establishment timing. The sooner the establishment time of the weed in relation to soybean emergence, the greater the yield reduction (Table 2). On the contrary, no differences in soybean yield occurred at A. palmeri establishment times of 2 WAE onward (Table 2). Amaranthus palmeri plants established with the crop (0 WAE) caused soybean reductions of 15% and 22% for 2014 and 2015, respectively, compared with yield recorded at 8 WAE A. palmeri establishment timing (Table 2). Yield reductions declined progressively with late weed establishment, that is, 12%, 5%, and 4% for 2, 4, and 6 WAE in 2014 and 17%, 9%, and 8% for 2, 4, and 6 WAE in 2015 when compared with the yields recorded at 8 WAE (Table 2). According to Ciuberkis et al. (Reference Ciuberkis, Bernotas, Raudonius and Felix2007), there is a positive relationship between the timing of weed emergence and the degree of the pressure exerted on the crop, which usually causes crop yield losses. In addition, Keramati et al. (Reference Keramati, Pirdashti, Esmaili, Abbasian and Habibi2008), Suryanto et al. (Reference Suryanto, Tohari Sulistyaningsih, Susila Putra, Kastono and Alam2017), and Korres et al (Reference Korres, Norsworthy and Mauromoustakos2019a) reported that late weed infestations (including A. palmeri) long enough after crop emergence rarely cause significant yield reductions in soybean.
It can be concluded that early establishment time of A. palmeri in relation to the crop (i.e., 0 and 1 WAE establishment timings) reduces soybean dry weight and pods produced per plant, a result that is particularly notable at low soybean density. In addition, earlier A. palmeri establishment (0, 1, and 2 WAE establishment timings) causes greater soybean yield reductions irrespective of crop density, as no interactions between A. palmeri establishment time and soybean density were recorded in both 2014 and 2015.
Effects of Soybean Plant Density and Weed Establishment Timing on Production Cost and Economic Returns
The soybean seed cost is one of the greatest production inputs, especially after the introduction of GM soybean cultivars in 1996, and subsequent attempts by technology and seed companies to protect their intellectual property (Epler and Staggenborg Reference Epler and Staggenborg2008; Shi et al. Reference Shi, Chavas and Stiegert2010; Thompson et al. Reference Thompson, Larson, Lambert, Roberts, Mengistu, Bellaloui and Walker2015). As farmers attempt to use seed inputs more effectively, decisions on seeding rate should be reevaluated occasionally (Thompson et al. Reference Thompson, Larson, Lambert, Roberts, Mengistu, Bellaloui and Walker2015). Inputs in this analysis were held constant, except those associated with seeding rates; hence the differences in final production costs for each cropping system (i.e., US$772 for 125,000 seeds ha−1, US$818 for 250,000 seeds ha−1, and US$873 for 400,000 seeds ha−1) are due to costs linked with increased seeding rate. Although, seed prices can vary widely due to seed traits such as conventional versus GM cultivars (Popp et al. Reference Popp, Edwards, Purcell and Manning2006), herbicide-tolerant (HT) soybeans dominate U.S. (94% by 2018) and worldwide (57% of the entire area under GM cultivation or 64% of the cultivated area worldwide in 2006) soybean cropping systems (Bonny Reference Bonny2008; USDA-NASS 2019a). HT soybean cultivation in the United States is approximately 35 million ha (USDA-FSA 2019) with LiberyLink® soybean occupying 7 million ha with a 25% potential increase in 2018 onward (Bayer 2017). Therefore, the estimations presented in this work are widely applicable, even though the budget analysis presented here aims to highlight the importance of a judicious seed rate as a sustainable tool for A. palmeri control. Increases in seeding rate resulted in increases of grain yield followed by increased crop revenue, and thus increased net returns: US$290, US$486, and US$493 ha−1 for 125,000, 250,000, and 400,000 seeds ha−1, respectively, in 2014 (Table 1). In 2015, net return was US$88, US$322, and US$226 ha−1 for 125,000, 250,000, and 400,000 seeds ha−1, respectively (Table 1). It is worth mentioning that the greatest net return of 250,000 seeds ha−1 compared with 400,000 seeds ha−1, due to high yields in both years, indicating the importance of selecting a judicious seed rate as an added-value agronomic and weed management tool. The lower the grain yield, the greater the breakeven price, as in the case of the low seeding rate of 125,000 seeds ha−1 compared with the 250,000 and 400,000 seeds ha−1 seeding rates (Table 1). According to Hofstrand (Reference Hofstrand2018) and Dillon (Reference Dillon1993), breakeven prices denote the lower price level at which a farmer can market the product and still be able to cover the production costs. In this study, lower breakeven prices, such as those obtained for the two high seeding rates (i.e., 250,000 and 400,000 seeds ha−1), permit the establishment of a solid marketing foundation and facilitate marketing decisions that favor the optimization of the production system (Dillon Reference Dillon1993) through the selection of competing production alternatives. The inclusion of miscellaneous overhead, crop insurance, interest, fixed machinery cost, fixed building cost, land real estate taxes, land interest, and other costs in the crop budget analysis would have most probably increased the breakeven prices, as the numerator for its calculation would have increased if they were included in the analysis.
Earlier A. palmeri establishment timings (i.e., 0, 1, and 2 WAE establishment timings) resulted in greater yield reductions compared with later A. palmeri establishment timings (i.e., 4, 6, and 8 WAE) for both 2014 and 2015, with subsequent decreases in crop revenue and net returns (Table 2). As stated by Bensch et al. (Reference Bensch, Horak and Peterson2003), Keramati et al. (Reference Keramati, Pirdashti, Esmaili, Abbasian and Habibi2008), Korres et al. (Reference Korres, Norsworthy and Mauromoustakos2019a), and Suryanto et al. (Reference Suryanto, Tohari Sulistyaningsih, Susila Putra, Kastono and Alam2017), late weed infestations (i.e., an 8 WAE weed establishment time) rarely cause significant soybean yield losses. Therefore, lower yields resulted in greater breakeven prices, as in the case of A. palmeri establishment times of 0, 1, and 2 WAE (Table 2). The disadvantages of lower breakeven prices, as discussed earlier, also apply in this scenario.
Disclaimer
This research was supported in part by an appointment to the Agricultural Research Service (ARS) Research Participation Program administered by the Oak Ridge Institute for Science and Education (ORISE) through an interagency agreement between the U.S. Department of Energy (DOE) and the U.S. Department of Agriculture (USDA). Any opinions, findings, conclusions, or recommendations expressed in this publication are those of the author(s) and do not necessarily reflect the view of the DOE or USDA. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement. USDA is an equal opportunity provider and employer.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/wsc.2020.41










 Open access
Open access