



Introduction
The genus Echinochloa includes numerous problematic rice (Oryza sativa L.) weeds, such as Echinochloa crus-galli var. crus-galli (L.) P. Beauv. (EC) (Holm et al. Reference Holm, Pancho, Herberger and Plucknett1979), Echinochloa crus-galli var. mitis (Pursh) Peterm. (ECM), and Echinochloa glabrescens Munro ex Hook f. (EG) (Chen et al. Reference Chen, Tang, Li, Lu and Dong2019) in rice-planting areas in Asia. EC, ECM, and EG are very similar in morphology and are usually treated as a single species in weed management practices. Spikelets of EC are 3 to 4 mm in length, and the lower lemma extends into an awn to 3 cm. Spikelets of ECM are about 3 mm in length and awnless or with awns shorter than 5 mm, while inflorescences of ECM are usually branched. The lower lemma of EG is convex, coriaceous, and shining; while the lower lemmas of EC and ECM are flat and herbaceous (Chen et al. Reference Chen, Tang, Li, Lu and Dong2019; Chen and Phillips Reference Chen and Phillips2006). Our previous studies found that the harmfulness of ECM and EG is not lower than that of EC and even higher in some regions. Zhang et al. (Reference Zhang, Li, Zhang and Yang2014) found that 6 plants m−2 of EC and ECM resulted in 10.8% to 25.3% and 19.2% to 39.7% reduction in rice yield, respectively. Opena et al. (Reference Opena, Chauhan and Baltazar2014) found that EG may cause a yield loss of 7% to 87%.
EC, ECM, and EG propagate by seed (Chen et al. Reference Chen, Chen, Yu, Zhou and Zhuang2022; Li Reference Li1998). Therefore, knowledge on seed germination of these weed species is of key importance for their integrated management (Y Chen et al. Reference Chen, An, Chen and Zhuang2023; Masin et al. Reference Masin, Loddo, Gasparini, Otto and Zanin2014; Rezvani et al. Reference Rezvani, Nadimi, Zaefarian and Chauhan2021). Temperature is the primary factor influencing seed germination. EC did not germinate below 10 C, and its percentage germination increased with temperature increasing from 10 C to 24 C (Loddo et al. Reference Loddo, Ghaderi-Far, Rastegar and Masin2018). The optimal germination temperature for EC was around 25 to 30C (Kovach et al. Reference Kovach, Widrlechner and Brenner2010). EC buried in the autumn had a higher average percentage germination than seeds buried in the spring, and the optimal germination temperature for EC decreased with increasing seed age (Martinkova and Honek Reference Martinkova and Honek2013; Martinkova et al. Reference Martinkova, Honek and Lukas2006). In the case of alternating light and dark, the percentage germination of EG populations at 25/15 C, 30/20 C, and 35/25 C were similar (Opena et al. Reference Opena, Chauhan and Baltazar2014). The average percentage germination of ECM was 54.0% at 25 C and 0% at 17 C.
To date, research on Echinochloa species has been mainly on EC, and most studies on the seed biology of Echinochloa species were based on one to two populations. Yoshioka et al. (Reference Yoshioka, Satoh and Yamasue1998) confirmed soil CO2 was responsible for causing intermittent flushes of germination by testing 50 EC populations. The differences in biological and ecological traits among these three species remain undefined. Considering the widespread distribution of the three Echinochloa species and the seriousness of their damage in rice fields, a comparative study of the variations between EC, ECM, and EG is necessary. It is the basis for differentiated and efficient integrated management.
In 2022, we collected 66, 141, and 120 populations of EC, ECM, and EG, respectively, from rice fields in East China. We compared the germination characteristics of these populations under different temperature regimes. It had been reported that there were significant intraspecific differences in the initial germination temperature of EC (Royo-Esnal et al. Reference Royo-Esnal, Onofri, Loddo, Necajeva, Jensen, Economou, Taab, Synowiec, Calha, Andersson, Uludag, Uremis, Murdoch and Torresen2022). Nevertheless, the existence of intraspecific differences in ECM and EG had yet to be determined. With all 327 populations collected from rice fields in East China, the aims of this study were to (1) reveal intraspecific differences in seed germination of EC, ECM, and EG at different temperatures simulating different rice-planting periods; and (2) compare seed germination of EC, ECM, and EG at different temperatures.
Materials and Methods
Sampling and Investigation
In October 2022, we conducted field surveys on the occurrence of EC, ECM, and EG in rice fields of East China (Supplementary Table S1) and collected seeds from each rice field surveyed. A total of 250 independent rice fields (sites) where EC, ECM, or EG occurred were randomly surveyed, with an interval of >5 km for adjacent sites (Figure 1; Supplementary Table S1). Each site surveyed a rice field covering an area of about 0.1 to 0.2 ha. The identification of Echinochloa species was based on our previous work (Chen et al. Reference Chen, Tang, Li, Lu and Dong2019). A total of 327 Echinochloa populations were collected: 66 EC, 141 ECM, and 120 EG populations. Panicles with mature seeds were randomly collected from more than 100 individuals of each population with a pollen bag (100 mesh, 30 cm by 45 cm), and mature seeds were collected by hand. The relative dominance of Echinochloa species in rice fields was scored 0.1, 0.5, 1, 2, 3, 4, or 5 by visual inspection according to Qiang (Reference Qiang2005) and our previous field surveys (Chen et al. Reference Chen, He and Qiang2013). Seeds were air-dried and stored in our lab at room temperature fluctuating from 15 C to 25 C. From March to June 2023, the 1,000-seed weight of each population was determined by weighing 5 replicates of 100 mature seeds.
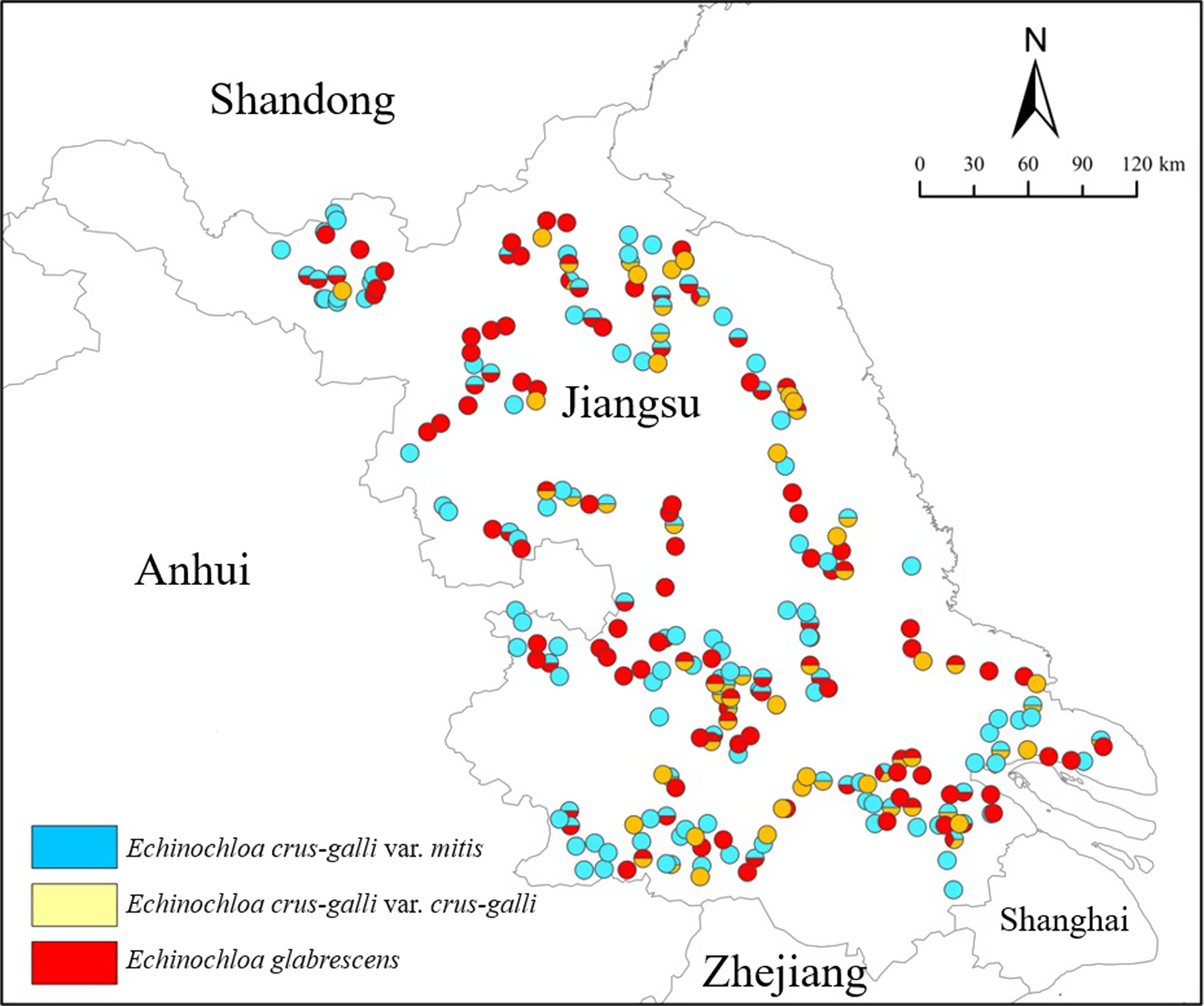
Figure 1. Collection sites of the three Echinochloa species populations in rice fields in China.
Experimental Design
To determine the adaptation of the three Echinochloa species to temperatures at the beginning of rice-planting season in East China, three temperature regimes were set responding to three different rice-cropping patterns in East China: single-cropping rice, double-cropping early rice, and double-cropping late rice. Single-cropping rice is mainly sown or transplanted around June 20, double-cropping early rice is usually transplanted around May 20, and double-cropping late rice is often transplanted around July 20. Therefore, seeds were incubated at alternating temperatures of 28/15 C (day/night), 30/20 C, and 35/25 C with a 12/12-h (light/dark) photoperiod to simulate the temperature of double-cropping early rice, single-cropping rice, and double-cropping late rice, respectively. From April to June 2023, about 7 mo after seed collection, seed germination for each population under the above three temperature regimes was determined in incubators (HBZ-400B, Changzhou Haibo Instrument Equipment, Changzhou, Jiangsu, China) with three replications. Fifty seeds from each population were placed into a 9-cm-diameter petri dish with two pieces of filter paper (Hangzhou Fuyang Beimu Pulp & Paper), which contained 6 ml of distilled water. The germinated seeds with a visible radicle were counted and removed (Rehman et al. Reference Rehman, Choi, Jamil and Yun2011). Percentage germination of each petri dish was determined daily for 21 d. From May to June 2024, 20% of 327 Echinochloa populations were selected randomly for germination repeatedly.
Statistical Analysis
The percentage germination of each dish was determined by the percentage of germinated seeds out of the number of sown seeds (50). The germination index for each dish was determined using Equation 1 (Schmer et al. Reference Schmer, Xue and Hendrickson2012):
 $$GI = \sum (GT/DT) $$
$$GI = \sum (GT/DT) $$
where GT is the number of seed germinations per day; and DT is the germination day corresponding to GT. Coefficient of variations (CVs) for variables were determined using Equation 2 (Munthali et al. Reference Munthali, Chirwa and Akinnifesi2012):
 $$CV = \sigma /\mu \times 100\% $$
$$CV = \sigma /\mu \times 100\% $$
where μ is the sample mean, and σ is the sample SD.
A three-parameter logistic function was fitted to test days and accumulated temperature required for germination (Equation 3), using the drc add-on package in R v. 3.1.3 (Ritz et al. Reference Ritz, Baty, Streibig and Gerhard2015):
 $$Y =a/[1 + (x/e) ^b] $$
$$Y =a/[1 + (x/e) ^b] $$
where Y denotes the total germination (%) at day x of the germination or accumulated temperature x after sowing; a is the upper limit; b indicates the slope; e is the days required for 50% of total germination (GD50) or the accumulated temperature required for 50% of total germination (TD50). Accordingly, the days required for 90% of total germination (GD90) and the accumulated temperature required for 90% of total germination (TD90) were determined.
To determine differences in germination indices among three temperature regimes, data were subjected to ANOVA in SPSS software v. 26.0 (IBM, Armonk, NY, USA), using the one-way ANOVA procedure. Data were checked for normality and constant variance before analysis. Treatment means were separated using the LSD test at P = 0.05. The general linear model (GLM) in SPSS was used to test data of environmental factors and germination indices; Echinochloa species, planting methods, and collection cities were set as dependent variable; GD50, GD90, and percentage germination were set as fixed factors. Correlations among [1] the latitudes and longitudes of seed collection sites and germination indices; and [2] 1,000-seed weight and relative dominance, percentage germination, and GD50 were determined with SPSS using correlation analysis. The independent-sample t-test method in SPSS software was used to compare the differences in percentage germinations of Echinochloa species collected from direct-seeded rice fields and transplanted rice fields at different temperature regimes. The data presented are means ± SEs.
Results and Discussion
Distribution and 1,000-Seed Weight
Among the 250 collection sites (Figure 1), 2 Echinochloa species co-occurred in 68 collection sites, and 3 Echinochloa species co-occurred in 5 collection sites. ECM was the most frequent species (43.0%), followed by EG (36.6%) and EC (20.4%). Average values of relative dominance among seeds collected in rice fields were 2.5, 2.5, and 2.7 (Supplementary Figure S1), for EC, ECM, and EG populations, respectively, which did not show significant differences. Echinochloa species differed in their average 1,000-seed weights. EC were significantly the heaviest (average 1,000-seed weight of 2.92 g), followed by EG and ECM with 1,000-seed weights of 2.40 g and 2.26 g, respectively (Figure 2). Interestingly, the 1,000-seed weight of the 327 Echinochloa populations showed significant and positive correlations with relative dominance, percentage germination, and germination index (Figures 3 and 4).

Figure 2. Thousand-seed weight of Echinochloa crus-galli var. crus-galli (EC), Echinochloa crus-galli var. mitis (ECM), and Echinochloa glabrescens (EG). The horizontal lines represent the average 1,000-seed weight of each of the three Echinochloa species. Different letters indicate significant differences among the three Echinochloa species.

Figure 3. Correlations between 1,000-seed weight and percentage germination and germination index at different temperature regimes for the three Echinochloa species.

Figure 4. Correlations between 1,000-seed weight and relative dominance for the three Echinochloa species.
Interspecific Differences
At 28/15 C, the average percentage germination of EC (67.5%) was significantly higher (P < 0.05) than that of ECM (46.4%) and EG (43.7%) (Figure 5), and the coefficients of variations (CVs) were 35.9, 43.2, and 44.2, respectively (Supplementary Table S2). The average percentage germinations of the three species at 30/20 C were like those of the same species at 28/15 C (Supplementary Table S3). Among the 120 EG populations, the latitude of population collection sites significantly and positively correlated with the percentage germination at 28/15 C and 30/20 C (Table 1); the longitudes of population collection sites significantly and negatively correlated with the percentage germination at 30/20 C. When temperature regime increased to 35/25 C, the average percentage germinations of EC, ECM, and EG were 90.7%, 80.5%, and 80.3%, and the coefficients of variation of EC, ECM, and EG decreased to 11.2, 17.5, and 17.8, respectively (Table 1; Supplementary Table S2).

Figure 5. Percentage germination of Echinochloa crus-galli var. crus-galli (EC), Echinochloa crus-galli var. mitis (ECM), and Echinochloa glabrescens (EG) at 28/15 C, 30/20 C, and 35/25 C with a 12/12-h (light/dark) photoperiod (12L/12D). The horizontal lines represent the average percentage germinations of each of the three Echinochloa species. Different letters indicate significant differences among three Echinochloa species in the same figure.
Table 1. The correlation between the latitude (Lat.) and longitude (Long.) of the collection sites and the germination indices of three Echinochloa species populations. Values are correlation values and not GPS coordinates.

a GD50, days required for 50% of total germination; GD90, days required for 90% of total germination.
* Significant effect at P < 0.05. NS note was removed, because (1) NS does not need to be defined and (2) there are no entries with “NS” in Table 1.
The average germination indices of EC, ECM, and EG were 5.6, 3.8, and 3.6 at 28/15 C, respectively (Figure 6), and the coefficients of variation were 41.0, 55.7, and 52.2, respectively. At 35/25 C, the average germination indices of EC, ECM, and EG were also significantly highest among different temperature treatments, with 13.7, 9.8, and 9.7, respectively. The percentage germination and germination index of EC was higher than that of the other two species regardless of temperature regimes.

Figure 6. Germination index for Echinochloa crus-galli var. crus-galli (EC), Echinochloa crus-galli var. mitis (ECM), and Echinochloa glabrescens (EG) at 28/15 C, 30/20 C, and 35/25 C with a 12/12-h (light/dark) photoperiod (12L/12D). The horizontal lines represent the average germination indices of each of the three Echinochloa species. Different letters indicate significant differences among three Echinochloa species in the same figure.
The percentage germination and germination process of EC were significantly higher and faster than those of ECM and EG at the same temperature treatments, which might be related to seed mass. Zhou et al. (Reference Zhou, Yu, Yang, Chen, Yin and Ding2021) found that the smaller common ragweed (Ambrosia artemisiifolia L.) seeds germinated faster with a study of 26 populations. Here, the 1,000-seed of EC was heavier than those of ECM and EG by 29.2% and 21.7%, respectively (Figure 2). Meanwhile, species of Echinochloa and the city where the population was collected had significant influences on the GD50 and GD90 values and percentage germination (Table 2). Moreover, percentage germination at high temperatures was significantly influenced by the planting method (direct seeding or transplanting) at the site where the seed was collected. Specifically, at 30/20 C, average germination percentage of overall populations collected from transplanted rice fields (62.9 ± 2.6%) were significantly higher than that of overall populations collected from direct-seeded rice fields (52.5 ± 2.1%), as well as the comparison at 35/25 C (85.9 ± 1.7% vs. 76.9 ± 1.5%) (Supplementary Table S4). The chemical control time window against Echinochloa species in rice fields is before planting or sowing to the rice bolting stage, which is about 10 d shorter in transplanted rice fields than in direct-seeded rice fields (Li et al. Reference Li, Sun, He, Wang, Wang and Lou2015; Sun et al. Reference Sun, Lu, Yin and Fan2014). Thus, Echinochloa species populations escaping chemical control applications tended to have a longer growth period in transplanted rice fields than those in direct-seeded rice fields. This longer development time often results in higher-quality seeds (Chen et al. Reference Chen, Sun, Li, Ma, Tian, Chen, Yang and Yin2017), which could lead to a significantly high percentage germination of Echinochloa seeds collected from transplanted rice fields.
Table 2. The effects (F-values) of species, planting method, and city on three germination indices determined by generalized linear models.

a GD50, days required for 50% of total germination; GD90, days required for 90% of total germination.
* Significant effect at P < 0.05.
Compared with the average GD50 and GD90 values for ECM and EG, those of EC were significantly highest among the three temperature regimes (Figures 7 and 8). The GD50 values for Echinochloa species were significantly and positively correlated with the latitude of population collection sites and negatively correlated with the longitudes of population collection at 35/25 C, respectively. Considering the different climate zones in the world, accumulated temperature was more universal (Hou et al. Reference Hou, Liu, Xie, Ming, Ma, Li and Mei2014). At 28/15 C, the average accumulated temperatures of EC, ECM, and EG to 50% total germination were 111.0 C, 127.3 C, and 124.7 C, respectively (Table 3). At 35/25 C, the average accumulated temperatures of EC, ECM, and EG to 50% total germination were 74.3 C, 76.4 C, and 80.2 C, respectively. Six indices of ECM and EG seed germination showed no significant differences at the same temperature treatments, excluding GD90 and TD90 at 30/20 C and 35/25 C (Supplementary Table S5).

Figure 7. Number of days to 50% total germination rate (GD50) of Echinochloa crus-galli var. crus-galli (EC), Echinochloa crus-galli var. mitis (ECM), and Echinochloa glabrescens (EG) at 28/15 C, 30/20 C and 35/25 C with a 12/12-h (light/dark) photoperiod (12L/12D). The horizontal lines represent the average GD50 values of each of the three Echinochloa species. Different letters indicate significant differences among three Echinochloa species in the same figure.

Figure 8. Number of days to 90% total germination rate (GD90) for Echinochloa crus-galli var. crus-galli (EC), Echinochloa crus-galli var. mitis (ECM), and Echinochloa glabrescens (EG) at 28/15 C, 30/20 C and 35/25 C with a 12/12-h (light/dark) photoperiod (12L/12D). The horizontal lines represent the average GD90 values of each of the three Echinochloa species. Different letters indicate significant differences among three Echinochloa species in the same figure.
Table 3. The accumulated temperature to 50% and 90% total germination (TD50 and TD90) of Echinochloa species at different temperature regimes a .
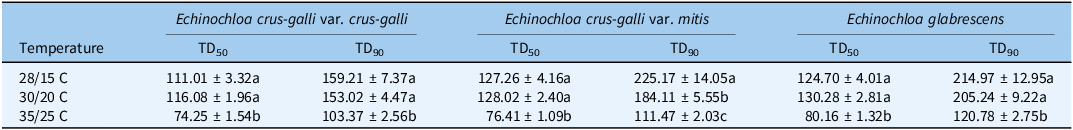
a Different letters in the same column indicate significant differences among the three temperature regimes at P < 0.05.
Interspecific differences were obvious among the three Echinochloa species, and seed germination could be affected by the latitudes and longitudes of the population collection sites (Zhou et al. Reference Zhou, Yu, Yang, Chen, Yin and Ding2021). Cheng et al. (Reference Cheng, Huang, Liu, Zhou, Han, Wang and Zhang2022) found that smooth cordgrass (Spartina alterniflora Loisel.) seeds germinated earlier at higher latitudes. In our results, germination indices of EG were also impacted by the latitudes and longitudes of collection sites (Table 1). EG seeds in the northwest region had a lighter 1,000-seed weight (Supplementary Table S6), the percentage germination tended to be higher, and the germination process tended to be slower. Furthermore, the city where the population was collected significantly affected the seed percentage germination and germination process of tested Echinochloa species populations (Table 2). This effect may be related to the influence of local rice cultivation, field management, climate, and other environmental factors (Li et al. Reference Li, Yin, Liu, Li and Zhou2009). Among these factors, previous crops were the most critical for weed impact, followed by tillage intensity and environmental parameters (Hanzlik and Gerowitt Reference Hanzlik and Gerowitt2011). Additionally, geographic location and agricultural practices could affect the genetic variation of EC (Altop and Mennan Reference Altop and Mennan2011).
Intraspecific Differences
Percentage germination and germination index of the three Echinochloa species increased with temperature increments, which were significant at 35/25 C. At 28/15 C, the average GD50 and GD90 values of EC (5.2 and 7.4), ECM (5.9 and 10.5), and EG (5.8 and 10.0) were significantly highest, followed by treatments at 30/20 C with EC (4.6 and 6.1), ECM (5.1 and 7.4), and EG (5.2 and 8.2). At 35/25 C, the GD50 and GD90 values for EC (2.5 and 3.4), ECM (2.6 and 3.8), and EG (2.7 and 4.0) were all significantly the lowest among different temperatures. The GD50 and GD90 values of EC were the lowest at the same temperature conditions, and the GD90s of ECM and EG had significant differences at 35/25 C (Supplementary Tables S3 and S4).
Seed germination of the 66 EC populations at different temperatures simulating different rice-planting periods varied considerably, and intraspecific differences decreased significantly with increasing temperature (Supplementary Tables S2 and S3). Royo-Esnal et al. (Reference Royo-Esnal, Onofri, Loddo, Necajeva, Jensen, Economou, Taab, Synowiec, Calha, Andersson, Uludag, Uremis, Murdoch and Torresen2022) found that one EC population collected in Norway tended to emerge earlier than another population collected in Italy. Intraspecific variations in seed percentage germination and GD50 were also found in a study with 25 EC populations (Martinkova and Honek Reference Martinkova and Honek1997). The 66 EC populations in the present study produced CVs of 35.9 and 24.3 in percentage germination and GD50 values at 28/15 C, while the CVs of both indices decreased dramatically at 35/25 C. Serra et al. (Reference Serra, Fogliatto and Vidotto2018) found EC populations exhibited varying degrees of adaptability to environmental conditions during the seed germination stage. Intraspecific variations included population-level variations, between-individual variations, and within-individual variations, which were affected by the maternal plant (Albert et al. Reference Albert, Grassein, Schurr, Vieilledent and Violle2011). Our study was at the population-level, so these intraspecific variations would be affected by genotypic compositions of populations and a temporal variability in the environment. Intraspecific variations in the germination process and the percentage germination of EC were notable at low temperatures, and such variations narrowed with increasing temperatures. Increasing temperatures increased the percentage germination and accelerated the germination process (Bastiani et al. Reference Bastiani, Lamego, Nunes, Moura, Wickert and Oliveira2015; Derakhshan et al. Reference Derakhshan, Bakhshandeh, Siadat, Moradi-Telavat and Andarzian2018). Marambe and Amarasinghe (Reference Marambe and Amarasinghe2002) found percentage germinations of EC at 34/31 C were 27% to 29% higher compared with those at 28/24 C with two populations. In this study, the average percentage germination of the 66 EC populations at 35/25 C was 90.7%, which was significantly higher than those at 28/15 C and 30/20 C, respectively; the GD50 of the 66 EC populations at 35/25 C was 2.5 d, which was significantly lower than those at 28/15 C and 30/20 C. The CV of the average percentage germination among the 66 EC populations decreased from 40.7 to 11.2 as with treatment temperature increased from 30/20 C to 35/25 C, as did the germination index, GD50, and TD50.
Seed germination of the 141 ECM and the 120 EG populations also showed notable intraspecific variations, which also decreased significantly with increasing temperature. When the temperature increased to 35/25 C, the CVs of GD50 among the 141 ECM and 120 EG populations significantly decreased to 16.9 and 18.1, respectively, as well as percentage germination, germination index, and TD50. The accumulated temperature could serve as a reference for the growth and development of wheat (Triticum aestivum L.), which required an accumulation of 70 to 80 C to produce each leaf on the main stem (Li et al. Reference Li, Yin, Liu, Li and Zhou2009). In contrast, the accumulated temperatures required for EC, ECM, and EG seed germination at high temperatures were significantly lower than those at low temperatures. This may be the less days that seed germination needed under high temperature regimes. Combined with data from experiment repetition in the second year (Supplementary Table S7), seed germination of the three Echinochloa species response to temperature had significant intraspecific variations.
Management Strategies
In management of rice weeds, different species of Echinochloa species are usually treated as a single species (Guo et al. Reference Guo, Qiu, Ye, Jin, Mao, Zhang, Yang, Peng, Wang, Jia, Lin, Li, Fu, Liu and Chen2017). Nevertheless, our results suggested interspecific differences among EC, ECM, and EG, as well as intraspecific variations among populations of the same species. Hence, management strategies against EC, ECM, and EG should also be customizable according to the target populations, especially when EC co-occurs with either ECM or EG in a rice field. The significant correlation between 1,000-seed weight, percentage germination, and relative dominance of Echinochloa weed populations is a warning to farmers to pay more attention to the rice fields with serious Echinochloa weed damage and formulate reasonable and effective control measures.
At the temperatures simulating the planting period of double-cropping early rice (28/15 C) and single-cropping rice (30/20 C), the percentage germinations of the three Echinochloa species ranged greatly, with a long duration. For long germination periods of the three Echinochloa species, onetime preemergence chemical control is not sufficient to control later-emerging seedlings. For double-cropping late rice (35/25 C), a majority of the 327 Echinochloa populations tested showed more than 90% germination within 3 to 4 d. Therefore, preemergence chemical control at 4 d after rice seeding or transplanting could be highly effective (GQ Chen et al. Reference Chen, An, Chen and Zhuang2023), as could stale seedbed strategies (Chen et al. Reference Chen, Chen, Yu, Zhou and Zhuang2022). When two or three Echinochloa species occurred in the same field at low temperatures (28/15 C and 30/25 C), species identification was crucial. At low temperatures, the same weed management can be applied to ECM and EG in view of their similar germination durations. However, the germination duration of EC was shorter compared with the other two species, making preemergence control should be applied within 5 to 6 d when EC occurrs in a rice field. Moreover, the longer germination periods of ECM and EG suggests they were more likely to partially evade preemergence herbicide treatment in rice fields. Consequently, management practices implemented too early may fail to completely control late-emerging seedlings, whereas management practices implemented too late will suffer from low efficacy (Marschner et al. Reference Marschner, Colucci, Stup, Westbrook, Brunharo, DiTommaso and Mesgaran2024), and thus repeated application of preemergence herbicides will be necessary. Also, considering the significant influence of collection location on seed germination characteristics, weed management strategies employed in local or adjacent areas could be used as important references to improve the efficiency of integrated management against Echinochloa rice weeds in paddy fields (Boddy et al. Reference Boddy, Bradford and Fischer2012; Rezvani et al. Reference Rezvani, Nadimi, Zaefarian and Chauhan2021).
In summary, we collected 66 EC, 141 ECM, and 120 EG populations in 250 rice fields surveyed in East China, and average values of relative dominance of the three species did not show significant differences. The 1,000-seed weight of the 327 Echinochloa populations overall showed significant and positive correlations with relative dominance, percentage germination, and germination index. Significant interspecific and intraspecific variations in seed germination characteristics under different temperatures were identified. EC showed significantly the highest seed biomass and percentage germinations, which made EC damage more probable and timely EC control vital; EG and ECM showed similar germination patterns with each other under all temperature regimes treated (28/15 C, 30/20 C, and 35/25 C). Percentage germination of the three Echinochloa species increased with increasing treatment temperatures, with intraspecific variations narrowing with increasing temperatures. The city where the population was collected had significant influence on the germination of Echinochloa species. At 30/20 C and 35/25 C, average germination percentages of populations collected from transplanted rice fields were significantly higher than those of populations collected from direct-seeded rice fields. The accumulated temperatures required for EC, ECM, and EG seed germination at higher temperatures were also significantly lower. Together, the findings of this study suggest that management strategies against EC, ECM, and EG, such as stale seedbed and preemergence chemical control, should be customized according to the target populations, in particular when the temperatures are not high.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/wsc.2024.104
Acknowledgments
We thank Hongcheng Zhang (Agricultural College, Yangzhou University, Yangzhou, China) for his guidance in this study.
Funding statement
This study was funded by the National Key R&D Program Projects (2021YFD1700100), Jiangsu Key R&D Plan (BE2022338), Jiangsu Agricultural Science and Technology Independent Innovation Fund (CX (20)1012), and a project funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Competing interests
The authors declare no conflicts of interest.














 Open access
Open access