



Introduction
Weed management in flax (Linum usitatissimum L.) is challenging due to its uncompetitive nature, resulting in yield losses ranging from 20% to 30% (Liu et al. Reference Liu, Mahoney, Sikkema and Swanton2009; Sánchez Vallduví and Sarandón Reference Sánchez Vallduví and Sarandón2011; Stevenson and Wright Reference Stevenson and Wright1996). Weed competition not only reduces flax seed yield but can also reduce seed oil content and iodine levels (Bell and Nalewaja Reference Bell and Nalewaja1968; Friesen Reference Friesen1986). Weed control in flax is further hampered by limited herbicide options for in-crop weed management. At present, Group 1 (acetyl CoA carboxylase [ACCase] inhibitors), Group 4 (synthetic auxins), and Group 6 (photosystem II inhibitors) herbicides are the widely used herbicides for flax in Canada (Flax Council of Canada 2017). The efficacy of these herbicides, especially ACCase herbicides, is declining due to the evolution of herbicide resistance (Beckie et al. Reference Beckie, Shirriff, Leeson, Hall, Harker, Dokken-Bouchard and Brenzil2020; Heap Reference Heap2021), further impairing weed management in flax.
Wild oat (Avena fatua L.) has been a troublesome weed in many cropping systems on the Canadian Prairies due to its competitive characteristics (Kirkland Reference Kirkland1993; Willenborg et al. Reference Willenborg, May, Gulden, Lafond and Shirtliffe2005) and its resistance to ACCase- and acetolactate synthase (ALS)-inhibiting herbicides (Beckie et al. Reference Beckie, Ashworth and Flower2019). Weed surveys have shown that A. fatua was found in 42% of flax fields surveyed, even after an in-crop herbicide had been applied (Leeson et al. Reference Leeson, Thomas, Hall, Brenzil, Andrews, Brown and Van Acker2005). Cleavers (Galium spp.) is the ninth most abundant weed species on the Canadian Prairies and has been increasing in relative abundance at a faster rate than almost any other weed over the past 30 yr (Leeson Reference Leeson2012; Leeson et al. Reference Leeson, Thomas, Hall, Brenzil, Andrews, Brown and Van Acker2005). Of the many Galium species, false cleavers (Galium spurium L.) is the primary species of concern on the prairies (De Roo et al. Reference De Roo, Eckstein, Benaragama, Beattie and Willenborg2019). Galium spurium can impact the quantity and quality of yield in flax crops due to its climbing and tangling nature, which causes lodging and difficulties in harvesting operations (Malik and Vanden Born Reference Malik and Vanden Born1987). Management of Galium species is complicated by the ability of populations to act as both winter or summer annuals under prairie conditions (Defelice Reference Defelice2002; Malik and Vanden Born Reference Malik and Vanden Born1987). Moreover, Galium’s germination time can highly vary within a season (De Roo Reference De Roo2016). Galium species have also developed resistance to ALS-inhibiting herbicides in some regions (Beckie et al. Reference Beckie, Shirriff, Leeson, Hall, Harker, Dokken-Bouchard and Brenzil2020), and these resistant populations appear to be increasing in frequency. Perhaps more concerning is that next to kochia [Bassia scoparia (L.) A.J. Scott], A. fatua and G. spurium have the highest risk rating for developing glyphosate resistance (Beckie Reference Beckie2010), thus demanding long-term sustainable weed management strategies.
Managing herbicide-resistant weeds calls for integrated weed management systems that restrict weed emergence, reduce weed growth and reproduction, minimize weed interference with crops, and reduce the build-up of weed seedbanks (Harker et al. Reference Harker, Mallory-Smith, Maxwell, Mortensen and Smith2017). To achieve these objectives, cropping systems need to be designed using multiple weed management strategies that are both short-term and long-term in nature. Short-term integrated weed management strategies such as competitive cultivars, high crop density, and narrow row spacing are the predominant strategies that have proven to be effective when integrated into cereal and broadleaf crops (Harker and O’Donovan Reference Harker and O’Donovan2013; Shirtliffe and Benaragama Reference Shirtliffe and Benaragama2014), including flax (Kurtenbach et al. Reference Kurtenbach, Johnson, Gulden, Duguid, Dyck and Willenborg2019). Such cultural strategies are effective in A. fatua management (Harker et al. Reference Harker, O’Donovan, Irvine, Turkington and Clayton2009; O’Donovan et al. Reference O’Donovan, Newman, Harker, Blackshaw and McAndrew1999) but less is known about G. spurium.
In addition to integrating short-term cultural weed control strategies, diversifying crop rotations can diversify available herbicide modes of action and reduce the number of herbicide applications required for weed control, both of which are important for long-term herbicide resistance management (Beckie and Harker Reference Beckie and Harker2017; Harker et al. Reference Harker, Mallory-Smith, Maxwell, Mortensen and Smith2017). Increasing crop rotational diversity imposes a greater number of stress and mortality factors at different stages of a weed’s life cycle (Liebman and Staver Reference Liebman, Staver, Liebman, Mohler and Staver2001; Teasdale Reference Teasdale and Zimdahl2017) and has been identified as an effective long-term strategy to manage weeds. In a global meta-analysis, Weisberger et al. (Reference Weisberger, Nichols and Liebman2019) found that diversifying crop rotations away from simple crop rotations reduced weed density by 49%. Further, their analysis showed that diversifying rotations with more functionally diverse crops, rather than just increasing species diversity, was crucial for weed management. Moving from less diverse to more functionally diverse cropping systems allows growers to rely on more indirect tactics such as competitive crops, varied planting dates from fall to delayed seeding in the spring, and harvesting dates. Having a silage crop allows an early harvest that prevents weed seed production and/or returns to the seedbank (Liebman and Nichols Reference Liebman and Nichols2020). Therefore, differences in crop growth, phenology, and management practices are critical in developing effective crop rotations for weed management. Rotating functionally diverse crops is even more critical when weeds show a wide diversity in morphological and physiological traits. Examples include A. fatua (Miller et al. Reference Miller, Nalewaja and Mulder1982) and G. spurium (De Roo et al. Reference De Roo, Eckstein, Benaragama, Beattie and Willenborg2019; Malik and Vanden Born Reference Malik and Vanden Born1987).
Crop rotations have been an essential weed management tool for farmers on the Canadian Prairies. Yet these crop rotations are mainly diversified using annual spring crops. Adoption of functionally diverse crop rotations, including perennials and annual crops, remains low (Leeson and Beckie 2014). Cropping systems with diverse annual crop rotations, including spring and winter cereals harvested for grain and forage combined with integrated cultural practices, could be viable options for extensively grain-based farmers. Initial work done by O’Donovan et al. (Reference O’Donovan, Harker, Turkington and Clayton2013) and Harker et al. (Reference Harker, O’Donovan, Turkington, Blackshaw, Lupwayi and Smith2016) identified that combining diverse crop rotations with high seeding rates and reduced herbicides rates was effective in managing A. fatua in barley (Hordeum vulgare L.) and canola (Brassica napus L.), respectively. Likewise, incorporation of perennial crops such as alfalfa (Medicago sativa L.) was reported to be an effective strategy to manage weeds in some cropping systems, especially with regard to A. fatua (Entz et al. Reference Entz, Bullied and Katepa-Mupondwa1995; Ominski et al. Reference Ominski, Entz and Kenkel1999). How such integrated systems (where diverse crop rotations are integrated with high seeding rates) can fit into flax-based rotations has not yet been explored. Flax is often grown in rotation with cereals to prevent diseases, and at least 3 yrs are recommended between the cultivation of two successive flax crops (Flax Council of Canada 2017). This allows for longer rotations with more functionally diverse crops to manage challenging weeds. Currently, adequate weed control cannot be achieved in flax if herbicides are excluded; thus, integrated cropping systems with reduced herbicides may be a good alternative. This study aimed to determine the effect of functionally diverse flax-based crop rotations with reduced herbicides and high seeding rates on the longer-term management of weeds. Avena fatua and G. spurium were used as test species to represent problematic monocot and dicot weeds on the Canadian Prairies. We hypothesize that functionally diverse crop rotations with reduced herbicides and with high seeding rates can replace conventional low-diversity flax rotations with herbicides for sustainable weed management.
Materials and Methods
Site Conditions
Field experiments were carried out at three locations in western Canada. One site was located at the Kernen Research Farm at Saskatoon, SK, Canada (52.152861°N, 106.544861°W) on a Black Chernozemic loam (pH: 7.7; organic matter: 2.9%), another at the Indian Head Research Farm, SK, Canada (50.53305°N, 103.65125°W) on a Black Chernozemic clay (pH: 7.4; organic matter: 3.4%), and one site at Carman Research Farm at Carman, MB, Canada (49.48836°N, 98.03913°W) on a Gleyed Black Chernozem (pH: 5.5; organic matter: 6%). The experiment was conducted for 5 yr, from 2015 to 2019.
Experimental Treatments and Design
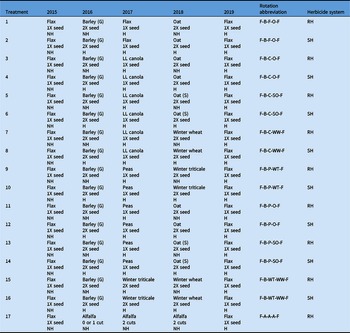
The experiment consisted of 17 treatments (crop rotations) having eight unique crop rotations with two herbicide regimes, giving 16 crop rotation combinations. Of the 16 combinations, eight rotations were under a standard herbicide system (SH), where in-crop herbicides for A. fatua and G. spurium control were applied in all phases except in the first-year flax phase. For comparison, the same rotations also were under a reduced herbicide system (RH), where no in-crop herbicides were applied during the second and fourth years. Crop rotations under both systems received in-crop herbicides in the final year of the flax phase. The 17th crop rotation treatment was considered the most functionally diverse crop rotation, as it comprised a 3-yr alfalfa crop under the RH system. The crop rotation flax–barley–flax–oat–flax under a standard herbicide system (Table 1) was considered the conventional crop rotation, and it was used as the rotation of comparison throughout the study. All crop rotations under both herbicide systems used a 2X seeding rate during the second and fourth years, while a 1X seeding rate was used during other phases. The only exception to this was the rotation with two consecutive winter cereal crops (flax–barley–winter triticale [×Triticosecale Wittm. ex A. Camus (Secale × Triticum)]–winter wheat [Triticum aestivum L.]–flax), in which both winter cereal phases were seeded at a 2X seeding rate. Thus, the seeding rate effect was uniform among all other crop rotations and was not considered as a treatment. All treatments were established in a randomized complete block design with four replicates. All crop rotations began with a flax crop and ended with a flax crop to determine changes in weed density and biomass as a function of the intervening treatments.
Table 1. Cropping systems (crop rotations and herbicide combinations) description at Kernen, SK, Carman, MB, and Indian Head, SK, Canada (2015–2019). a
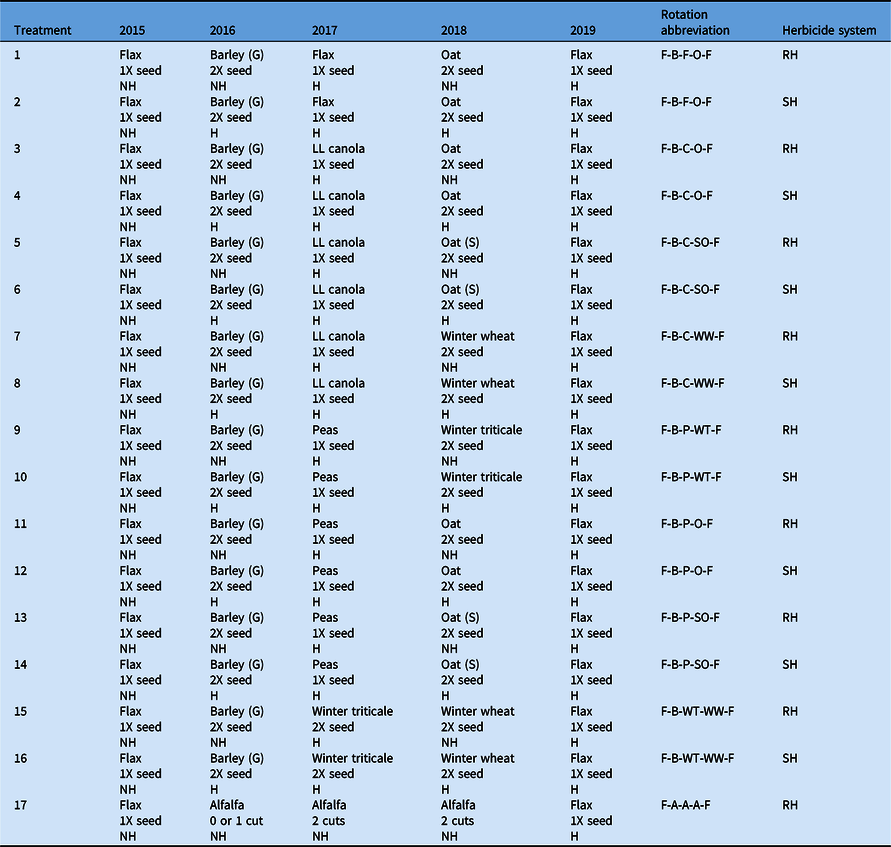
a Abbreviations: G, grain; H, herbicide applied; NH, no herbicide applied; S, silage ;SH, standard herbicide system; RH, reduced herbicide system; 1X, standard seeding rate; 2X, double the standard seeding rate; LL, LibertyLink®.
Crop Establishment and Weed Management
In the first year (2015), A. fatua was cross-seeded and G. spurium was broadcast over the entire experimental area at densities of 150 and 300 seeds m−2, respectively. At all three locations, crops were seeded using no-till seeders with hoe openers at 20-, 25-, and 30-cm row spacing at Carman, Kernen, and Indian Head, respectively. In all years, except in 2019, all standard herbicide treatments (SH system) received a glyphosate burndown before crop emergence. In 2015, no in-crop herbicides were applied to any plot during the first year to allow for weed establishment. In 2015, flax (‘CDC Glas’; SeCan, Kanata, ON, Canada, K2K 0E3) was direct-seeded into standing cereal stubble at a standard rate of 450 seeds m−2 in all plots. In 2016, treatments 1 to 16 were seeded to barley (‘CDC Copeland’; SeCan, Kanata, ON, Canada, K2K 0E3) at a 2X seeding rate (400 seeds m−2), and treatment 17 was seeded to alfalfa (‘Algonquin’; Cailliau Seed Company Ltd, Enchant, AB, Canada, T0K 0V0) at 9 kg ha−1. Barley under the RH system did not receive any A. fatua or G. spurium herbicides in the second year (2016), while barley under the SH system treatments received standard herbicides (Table 2). Herbicides were not applied to alfalfa in any year of the study. Alfalfa was cut twice during the growing season when the crop was at 10% bloom to simulate haying events. In the fall of 2016, treatments 15 and 16 were seeded to winter triticale (‘Luoma’; Corns Seeds, Grassy Lake, AB, Canada, T0K0Z0) at a 2X seeding rate (400 seeds m−2). Significant winter injury was evident in the spring of 2017 at both the Kernen and Indian Head locations; therefore, treatments were reseeded to spring triticale (‘Bunker’; FP Genetics, Regina, SK, Canada, S4N 6E1) at a 2X seeding rate (400 seeds m−2).
Table 2. In-crop herbicides used for the individual crops at Kernen, SK, Carman, MB, and Indian Head, SK, Canada. a
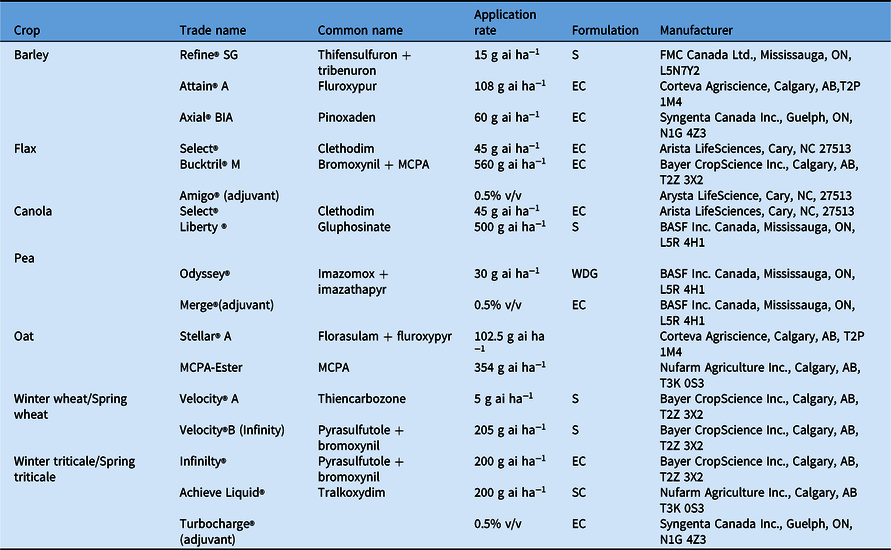
a Abbreviations: S, suspension; SG, soluble granule; SC, suspension concentrate; EC, emulsifiable concentrate; WDG, water-dispersible granules.
In 2017, treatments 1 and 2 were seeded to flax (CDC Glas) at 450 seeds m−2; treatments 3 to 8 were seeded to LibertyLink® canola (‘L130’; Bayer CropScience, Bayer CropScience Inc., Calgary, AB, Canada, T2Z 3X2) at 125 seeds m−2, and treatments 9 to 14 were seeded to field pea (‘CDC Meadow’; Sask Pulse Growers, Saskatoon, SK, Canada, S7N 3R3) at 90 seeds m−2. Treatments containing winter cereals were seeded with ‘Emerson’ (Canterra Seeds, Winnipeg, MB, Canada, R3T 1Y7) winter wheat and ‘Metzger’ (Haney Farm Ltd, AB, Canada, T0K 1V0) winter triticale at 400 seeds m−2. Again, low plant survivability was observed for overwintering cereals at Kernen in the spring of 2018, so spring wheat (‘Utmost’; FP Genetics, Regina, SK, Canada, S4N 6E1) and spring triticale (‘Bunker’; FP Genetics, Regina, SK, Canada, S4N 6E1) were seeded into winter cereal treatments to supplement winter cereal stands. Therefore, treatments 7, 8, 9, 10, 15, and 16 (Table 1) at Kernen and Indian Head were considered different from those treatments at Carman, as there was no winter cereal crop at Kernen and in some years at Indian Head. The crop rotations with winter wheat or winter triticale in the fourth year at Kernen and Indian Head were replaced with spring wheat and spring triticale, respectively, under both herbicide systems. Further, the crop rotation with two winter cereals (flax–barley–winter triticale–winter wheat–flax) was replaced with flax–barley–winter triticale–spring wheat–flax at Indian Head and flax–barley–spring triticale–spring wheat–flax at Kernen.
In 2018, ‘Triactor’ (Canterra Seeds, Winnipeg, MB, Canada, R3T 1Y7) was seeded in the grain oat (Avena sativa L.) treatments and ‘Haymaker’ (SeCan, Kanata, ON, Canada, K2K 0E3) was seeded into silage oat treatments, both at a density of 400 seeds m−2. Herbicides were not applied under the RH system in 2018, while they were under the SH system. Treatments seeded to silage oat were cut when the plants reached the soft dough stage at Zadoks 85 (Zadoks et al. Reference Zadoks, Chang and Konzak1974). In 2019, flax (CDC Glas) was direct seeded into all plots at a rate of 450 seeds m−2. Pre-seed burnoff was applied using 675 g ae ha−1 glyphosate + 18 g ai ha−1 carfentrazone at Kernen, 1.3 kg ae ha−1 glyphosate at Indian Head and 157 g ai ha−1 clopyralid + 880 g ai ha−1 MCPA-Ester at Carman.
Data Collection and Analysis
Weed Sampling
In all years, weed (A. fatua and G. spurium) emergence counts were taken at 2 to 3 wk after planting in two 1-m2 quadrats placed randomly at the front and back of each plot. Weed counts were taken at the species level to identify A. fatua and G. spurium. Aboveground weed biomass was taken when crops reached the late flowering stage at Zadoks 65. Shoot biomass of both weeds was sampled in 0.5-m2 quadrats placed randomly at the front and back of each plot. All biomass samples were placed in paper bags and oven-dried at 80 C for 48 h and then weighed.
Statistical Analysis
Data were analyzed using a repeated-measures ANOVA model using the PROC GLIMMIX procedure in SAS (SAS Institute 2011). Due to differences in crop rotations (differences in winter cereal treatment) among sites, data were analyzed separately for each site. Galium spurium and A. fatua density and biomass at all crop phases were considered for the analysis. Therefore, the weed density and weed biomass reported in this study are the average across all 5 yr of the study for each treatment. Cropping systems were considered as a fixed effect, while block was deemed to be random. Year (time) was considered a repeated factor, while plot was considered the repeated subject. After considering several covariance structures, a compound symmetry covariance structure was used with Akaike’s information criterion value. The assumptions (homogeneous variance and normal distribution of residuals) of the ANOVA were tested using PROC UNIVARIATE with the Shapiro-Wilk test and residual plots. Data were either log transformed or modeled with a negative binomial distribution depending on the residual analysis. All cropping systems were compared with the conventional crop rotation (flax–barley–flax–oat–flax under the SH system) using a Dunnett’s test. Treatment comparison was followed by a preplanned contrast carried out to compare different treatment groups. All data were presented on a back-transformed scale.
Results and Discussion
Growing Season Precipitation
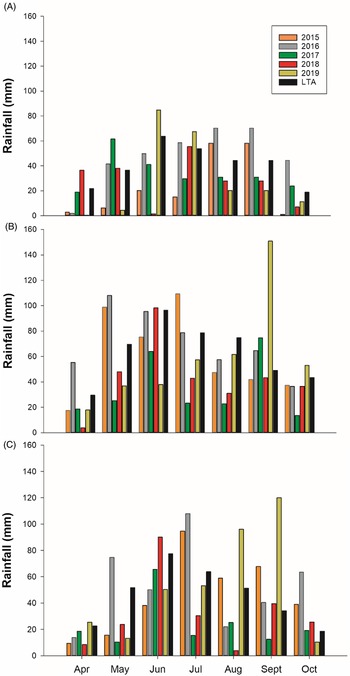
Total growing season precipitation at Kernen was lower than the long-term average (30-yr) in all years, with 2015 being the driest of the 5 yr (43% drier) (Figure 1A). At Carman, total precipitation was near the long-term average in 2015 and 2016, whereas 2017 and 2018 were 46% and 31% lower than the 30-yr long-term average, respectively (Figure 1B). Greater rainfall amounts were received in 2019 than in 2017 and 2018, but it was 13% drier than the 30-yr long-term average. At Indian Head, the years 2015, 2017, and 2018 were 30% lower than the long-term average (Figure 1C).
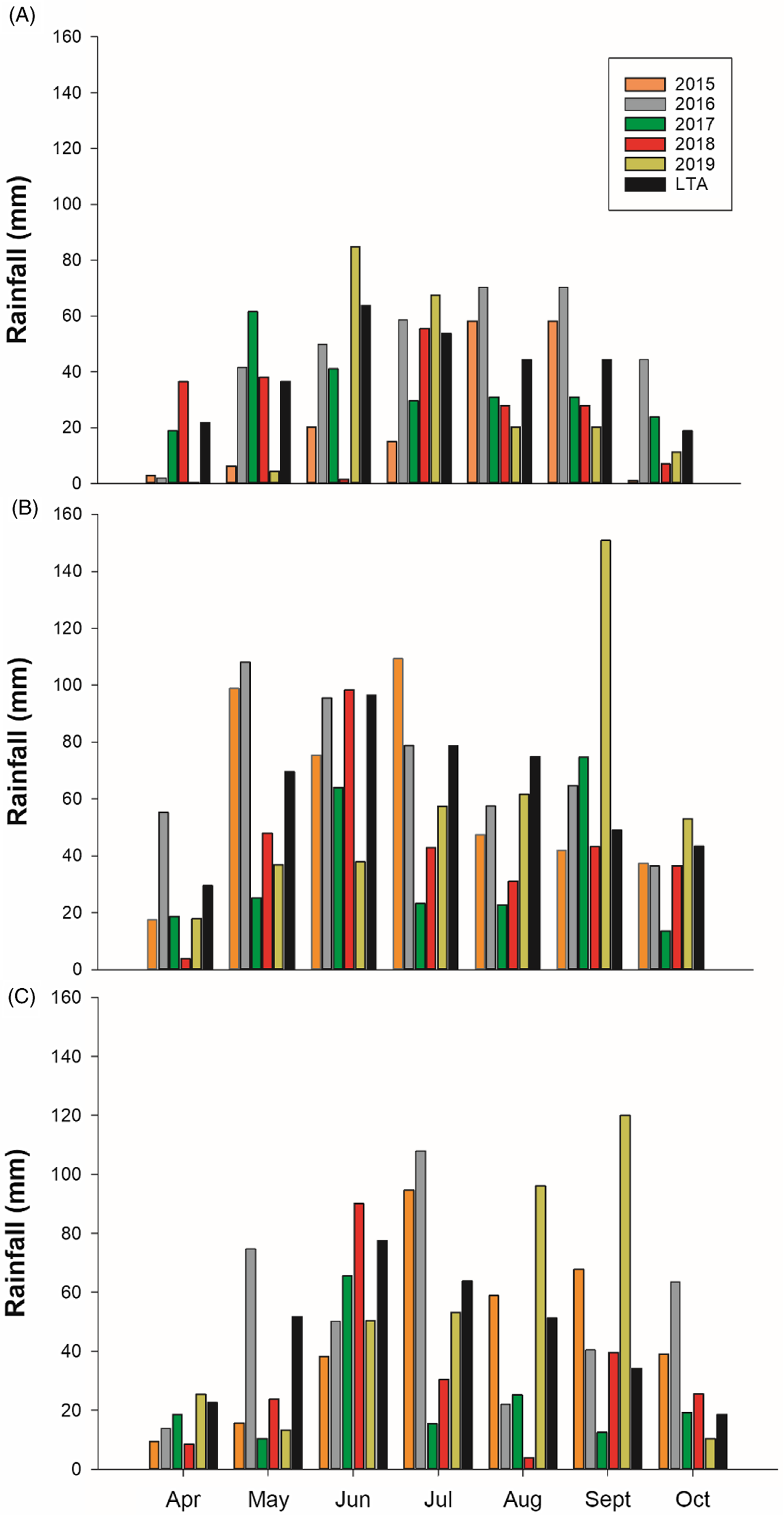
Figure 1. Monthly precipitation from April to October from 2015 to 2019 at (A) Kernen, SK, (B) Carman, MB, and (C) Indian Head, SK, Canada. LTA, long-term average.
Cropping Systems Effect on Weed Control at Each Location
Kernen
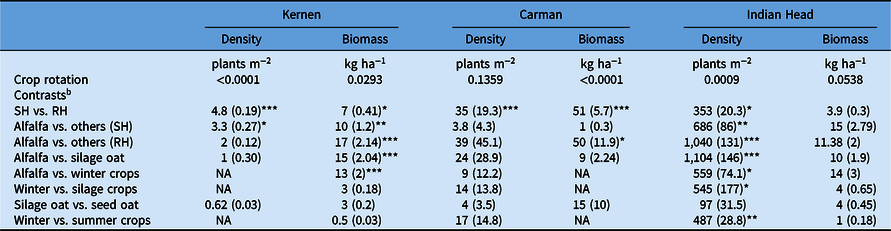
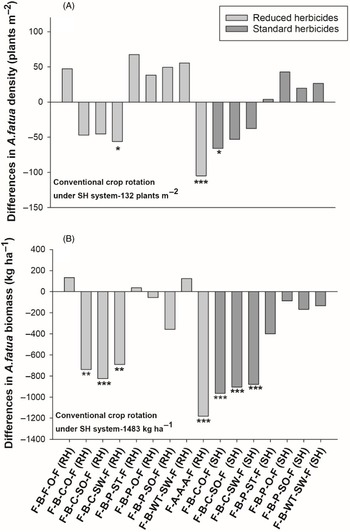
Crop rotations had differences in A. fatua density at Kernen. According to contrasts, the rotations under the SH system had 73% lower A. fatua density compared with the RH system (Table 3), indicating better weed control with the use of standard herbicides. All crop rotations in the RH system had significantly greater A. fatua density than the conventional rotation (flax–barley–flax–oat–flax under the SH system), which had a mean density of 17 plants m−2 (Figure 2A). The only exception was the alfalfa rotation, which had a statistically similar and low A. fatua density (28 plants m−2), indicating better A. fatua control than the other crop rotations compared with the conventional rotation. Contrasts indicated that the alfalfa rotation had a low A. fatua density compared with the average of all rotations under the RH system (84 plants m−2) and was comparable to the rotations under the SH system (22 plants m−2). Furthermore, the alfalfa rotation was found to have significantly lower A. fatua density than the rotations containing either silage crops or winter crops (Table 3). Across both herbicide systems (RH and SH), having silage oat in the rotation did not impact A. fatua densities differently than when oat was harvested for grain (Table 3).
Table 3. Probability values for crop rotations and mean differences for treatment contrasts for Avena fatua density and biomass at Kernen, SK, Carman, MB, and Indian Head, SK, Canada (2015–2019). a
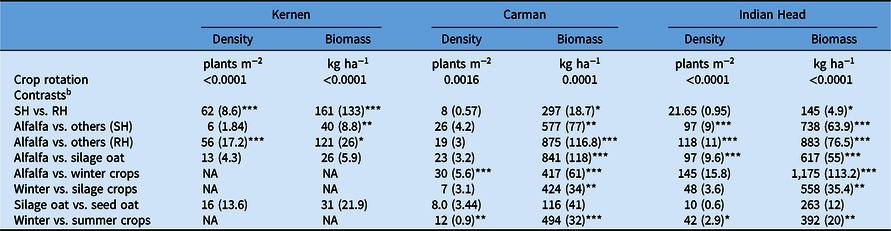
a Values in parentheses are standard errors for mean differences. Significant at: *P = 0.05; **P = 0.01; ***P = 0.001. NA, not applicable.
b Abbreviations: RH, no herbicides applied in the second and fourth year but herbicides applied in the fifth-year crop (flax); SH, full rate herbicides applied in the second-, fourth- and fifth-year crops.
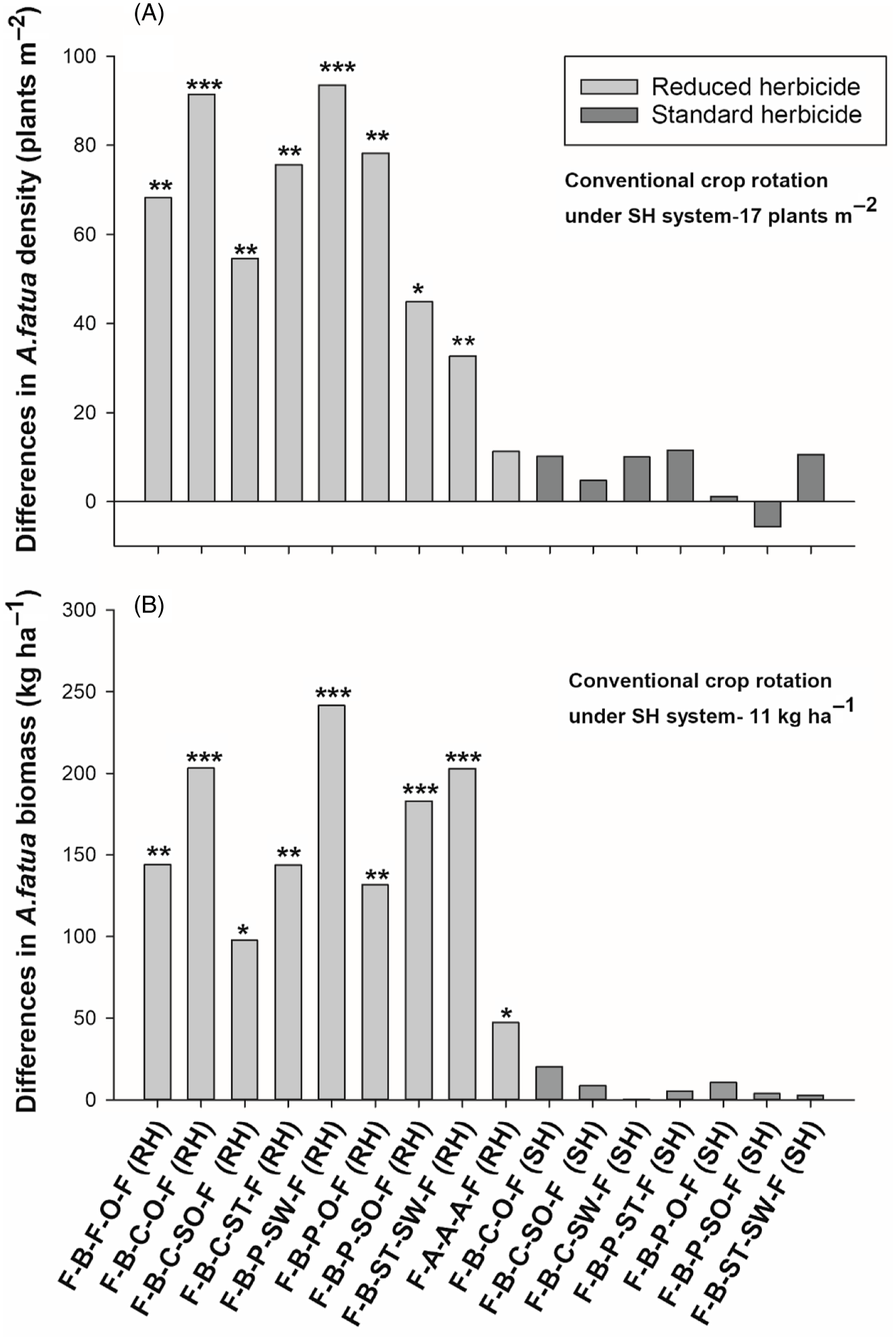
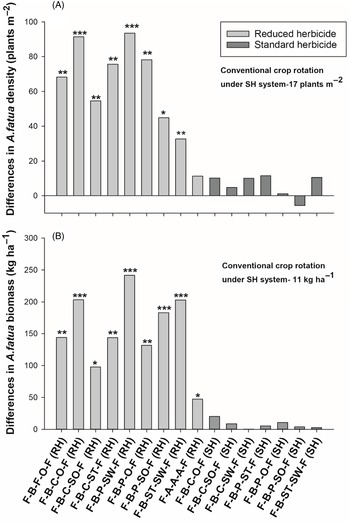
Figure 2. The effect of crop rotation on (A) Avena fatua density and (B) Avena. fatua biomass at Kernen, SK, Canada. Bars represent mean differences between the conventional crop rotation F-B-F-O-F (SH) and other crop rotations. Bars with asterisks indicate significantly different from the conventional crop rotation at *P = 0.05, **P = 0.01, and ***P = 0.001, obtained from Dunnett’s test. ST, spring triticale; SW, spring wheat; F, flax; A, alfalfa; B, barley; O, oat; SO, silage oat; P, pea; C, canola; RH, reduced herbicide; SH, standard herbicide.
Crop rotations also had significant differences in A. fatua biomass at Kernen (Table 3). Contrasts revealed that A. fatua biomass was lower in crop rotations in the SH system than in the RH system (90% reduction), indicating better A. fatua management was achieved with herbicides. In addition, all rotations under the RH system had significantly greater A. fatua biomass than the conventional crop rotation (11 kg ha−1) (Figure 2B). Further, contrasts revealed that the alfalfa rotation had lower A. fatua biomass (58 kg ha−1) compared with the average of all other rotations under the RH system (179 kg ha−1).
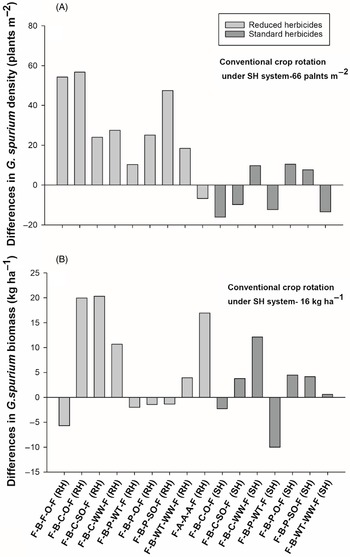
Crop rotations showed differences in G. spurium densities at Kernen, where the rotations under the SH system had a lower G. spurium density than under the RH system (Table 4). Galium spurium densities were greater in most rotations under the RH system compared with the conventional crop rotation under the SH system, including the grain oat, silage oat, spring triticale, spring wheat, and alfalfa rotations (Figure 3A). All other crop rotations had G. spurium densities similar to those of the conventional crop rotation under the SH system. Galium spurium biomass differed among crop rotations at Kernen (Table 4). Crop rotations with three flax crops and the rotations with spring triticale followed by spring wheat under the RH system had greater G. spurium biomass and density than the conventional crop rotation under the SH system. All other crop rotations had G. spurium biomass similar to that of the conventional rotation (Figure 3B). According to contrasts, the alfalfa rotation had lower G. spurium biomass (14 kg ha−1) than the average of all rotations under the RH system (64 kg ha−1).
Table 4. Probability values for crop rotations and mean differences for contrasts for Galium spurium density and biomass at Kernen, SK, Carman, MB, and Indian Head, SK, Canada (2015–2019). a
a Values in parentheses are standard errors for mean differences. Significant at: *P = 0.05; **P = 0.01; ***P = 0.001. NA, not applicable.
b RH, no herbicides applied in the second and fourth year but herbicides applied in the fifth-year crop (flax); SH, full rate herbicides applied in the second-, fourth- and fifth-year crops.
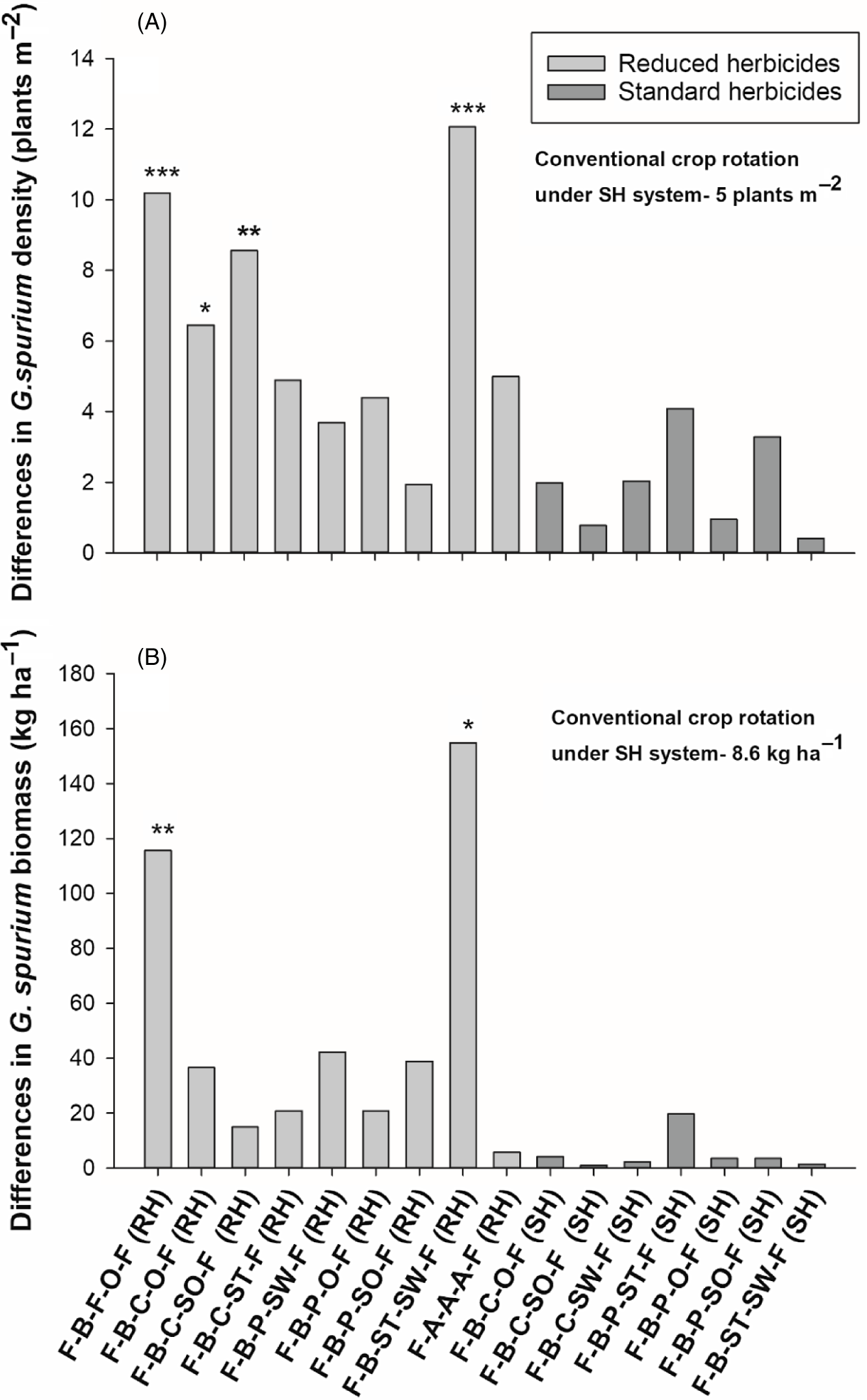
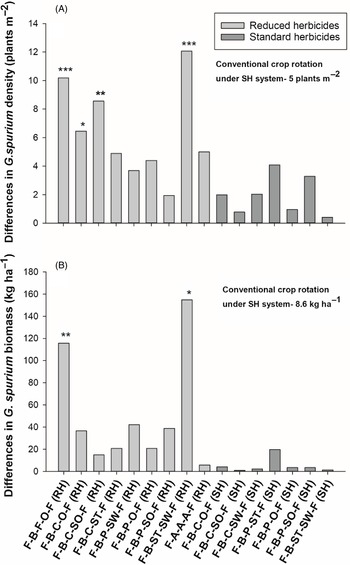
Figure 3. The effect of crop rotation on (A) Galium spurium density and (B) G. spurium biomass at Kernen, SK, Canada. Bars represent mean differences between the conventional crop rotation F-B-F-O-F (SH) and other crop rotations. Bars with asterisks indicate significantly different from the conventional crop rotation at *P = 0.05, **P = 0.01, and ***P = 0.001, obtained from Dunnett’s test. ST, spring triticale; SW, spring wheat; F, flax; A, alfalfa; B, barley; O, oat; SO, silage oat; P, pea; C, canola; RH, reduced herbicides; SH, standard herbicides.
Carman
Avena fatua density also differed among crop rotations at Carman (Table 3). Unlike at Kernen, A. fatua densities did not differ between the SH (65 plants m−2) and the RH systems (75 plants m−2) (Table 3). Crop rotations at the Carman site that had lower A. fatua densities than the conventional crop rotation included the rotation with two consecutive winter cereals as well as the rotations with grain oat and silage oat (Figure 4A). Lower A. fatua densities in the two winter cereal crop rotations under both herbicide systems were also found to be more effective at reducing A. fatua density (50% reduction) than having 3 yr of alfalfa. Further, the average of all winter crop rotations under both herbicide systems (61 plants m−2) had lower A. fatua density than the average of all crop rotations that included only summer annuals (74 plants m−2) (Table 3). Unfortunately, the Carman site was the only one that produced two winter crops in the rotation without losing them to early-season frost.
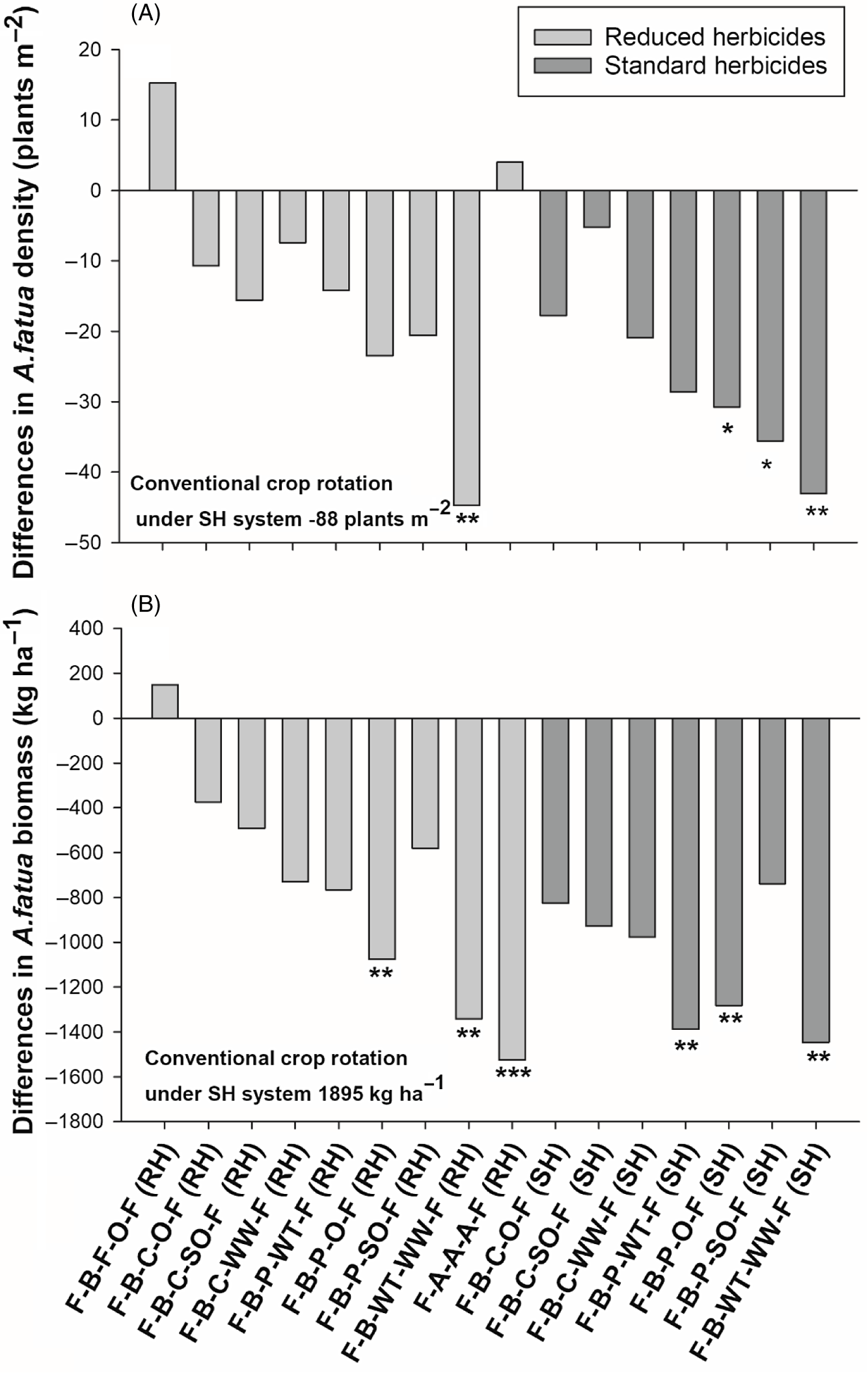
Figure 4. The effect of crop rotations on (A) Avena fatua density and (B) A. fatua biomass at Carman, MB, Canada. Bars represent mean differences between the conventional crop rotation F-B-F-O-F (SH) and other crop rotations. Bars with asterisks indicate significantly different from the conventional crop rotation at *P = 0.05, **P = 0.01, and ***P = 0.001, obtained from Dunnett’s test. F, flax; A, alfalfa; B, barley; O, oat; WT, winter triticale; WW, winter wheat; SO, silage oat; P, pea; C, canola; RH, reduced herbicides; SH, standard herbicides.
Avena fatua biomass at Carman was lower (23 %) in the rotations under the SH system compared with the RH system (Table 3). The crop rotations that had grain oat following pea and the rotation with two consecutive winter cereals had lower A. fatua biomass than the conventional crop rotation, regardless of the herbicide system (Figure 3B). Further, the alfalfa and single winter cereal crop rotation also exhibited lower A. fatua biomass compared with the conventional crop rotation (Figure 4B). The rotation containing alfalfa was found to have the greatest effect (80% biomass reduction) compared with all other systems in reducing A. fatua biomass. Further, the alfalfa rotation had lower A. fatua biomass than the average of all the rotations containing silage crops and winter cereal crops.
Overall, rotations in the SH system at the Carman site had lower G. spurium density than the rotations in the RH system (Table 4). Compared with the conventional crop rotation, only the alfalfa rotation exhibited a reduction (60 %) in G. spurium density (Figure 5A). Furthermore, contrasts indicated that the alfalfa rotation had a lower density of G. spurium compared with the average of all other crop rotations under both herbicide systems. This was also true when compared with the average of all the winter cereal–based rotations, which exhibited a similar G. spurium biomass (572 kg ha−1) compared with the conventional crop rotation (Figure 5B). The rotation with a single winter cereal crop and the rotation with grain oat following pea under the SH system also exhibited a similarly low G. spurium biomass compared with the conventional crop rotation. Similar to the Kernen site, contrasts indicated that the alfalfa rotation had lower G. spurium biomass than the average of all crop rotations regardless of herbicide system. Winter crop–based rotations showed better G. spurium management than the crop rotations with summer annual crops (Table 4).
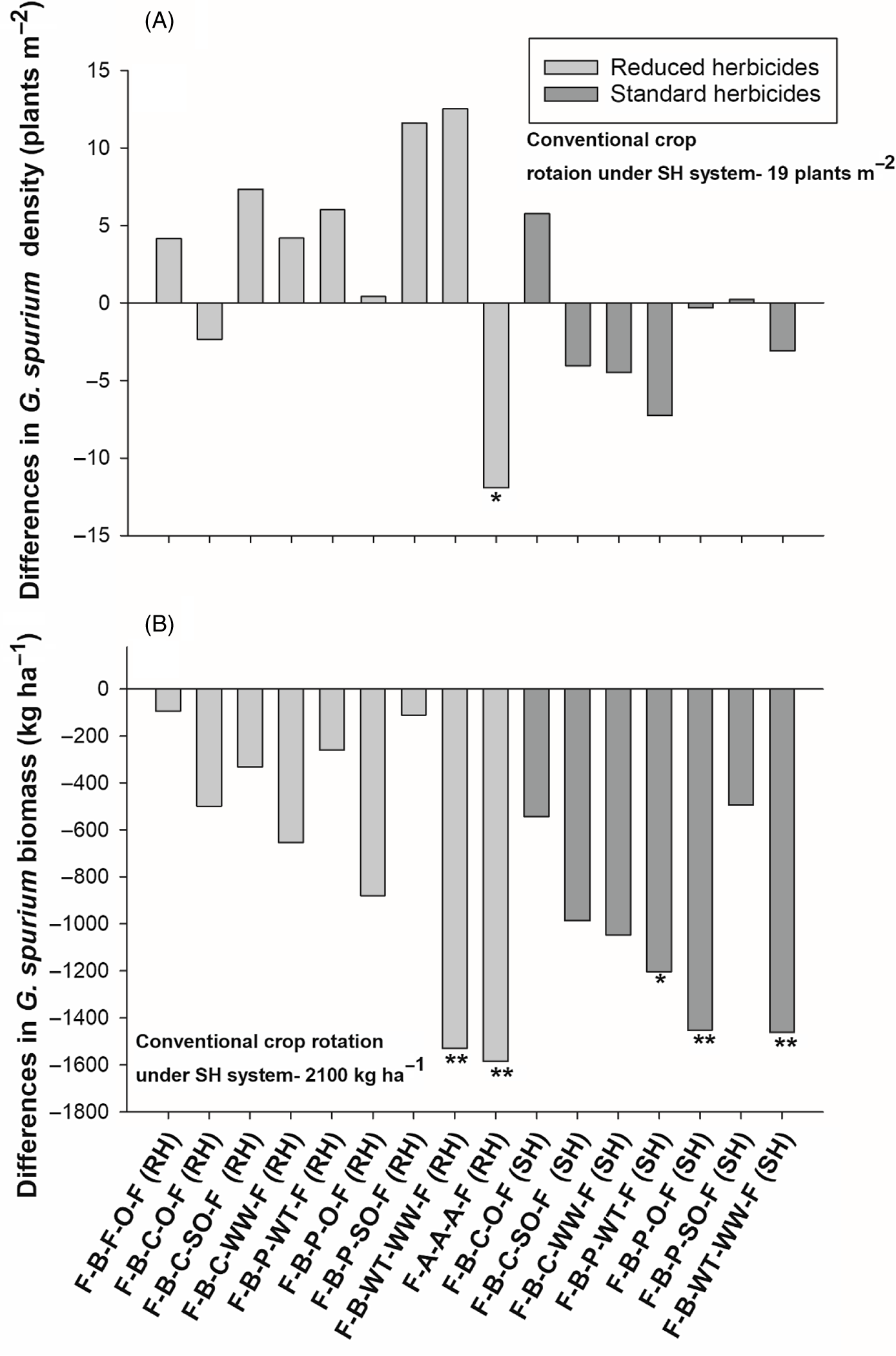
Figure 5. The effect of crop rotation on (A) Galium spurium density and (B) G. spurium biomass at Carman, MB, Canada. Bars represent mean differences between the conventional crop rotation F-B-F-O-F (SH) and other crop rotations. Bars with asterisks indicate significantly different from the conventional crop rotation at *P = 0.05 and **P = 0.01, obtained from Dunnett’s test. F, flax; A, alfalfa; B, barley; O, oat; WT, winter triticale; WW, winter wheat; SO, silage oat; P, pea; C, canola; RH, reduced herbicides; SH, standard herbicides.
Indian Head
Results at the Indian Head site differed somewhat from those of the other two sites. Avena fatua densities did not differ between the two herbicide systems based on contrasts (Table 3), indicating herbicides were less effective at reducing A. fatua density at that site. Among all crop rotations, the rotation with spring wheat following canola (flax–barley–canola–spring wheat–flax) under the RH system, the alfalfa rotation (flax–-alfalfa–-alfalfa–-alfalfa–-flax) and the rotation with seed oat following canola (flax–barley–canola–oat–flax) under the SH system had a significant reduction (42%, 80%, and 49%, respectively) in A. fatua densities compared with the conventional rotation (Figure 6A). Contrasts revealed that the alfalfa rotation had lower A. fatua density (27 plants m−2) compared with the average of all rotations under both herbicide systems (146 and 124 plants m−2, respectively). Furthermore, A. fatua density in the alfalfa rotation was lower than the average of all rotations containing both winter crops (173 plants m−2) and silage crops (125 plants m−2) under both systems.
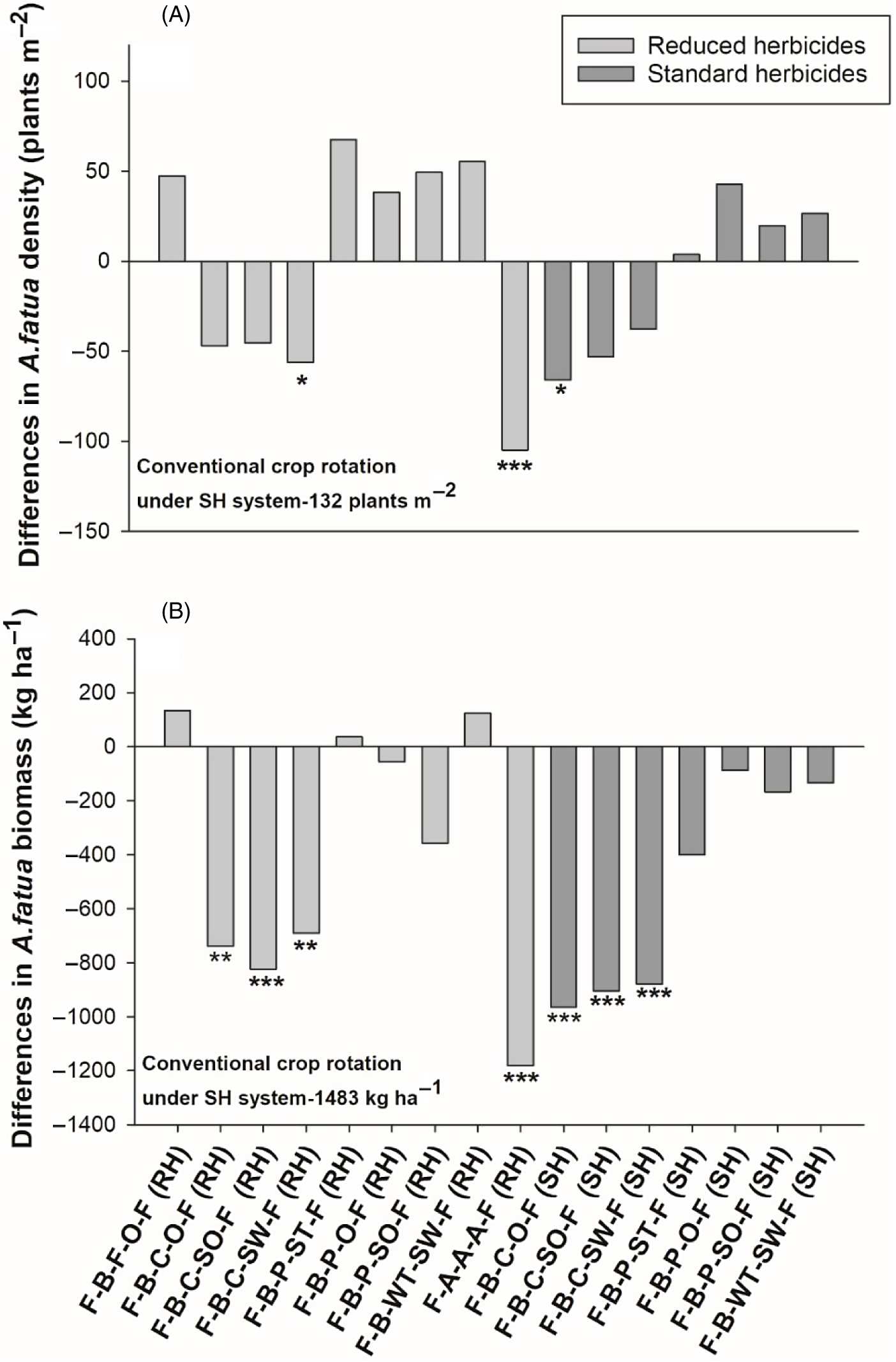
Figure 6. The effect of crop rotation on (A) Avena fatua density and (B) A. fatua biomass at Indian Head, SK, Canada. Bars represent mean differences between the conventional crop rotation F-B-F-O-F (SH) and other crop rotations. Bars with asterisks indicate significantly different from the conventional crop rotation at *P = 0.05 and **P = 0.01, obtained from Dunnett’s test. ST, spring triticale; SW, spring wheat; F, flax; A, alfalfa; B, barley; O, oat; WT, winter triticale; WW, winter wheat; SO, silage oat; P, pea; C, canola; RH, reduced herbicides; SH, standard herbicides.
Avena fatua biomass also differed among crop rotations at Indian Head (Table 3). At this site, the crop rotations with grain oat following canola, silage oat following canola, and spring wheat following canola all had significantly lower A. fatua biomass under both herbicide systems (Figure 6B). The alfalfa rotation had the greatest reduction in A. fatua biomass (79%) relative to the conventional crop rotation, as observed at the other two sites. Contrasts showed that the rotation containing alfalfa exhibited significantly lower A. fatua biomass (300 kg ha−1) compared with the average of all other rotations under both SH (1,041 kg ha−1) and RH (1,187 kg ha−1) systems (Table 3). The alfalfa rotation also had a lower A. fatua biomass than the rotations with winter crops and silage crops under the RH system. Further, contrasts revealed that crop rotations with winter crops were better at reducing A. fatua biomass than rotations with silage crops (Table 3).
Crop rotations system did not exhibit differences in G. spurium density or biomass at Indian Head (Table 4). Further, none of the crop rotations showed significant differences in G. spurium density or biomass compared with the conventional crop rotation at Indian Head (Figure 7A and B). Contrasts indicated that rotations under the SH system had lower G. spurium density than rotations under the RH system, but no differences in biomass were identified (Table 4).
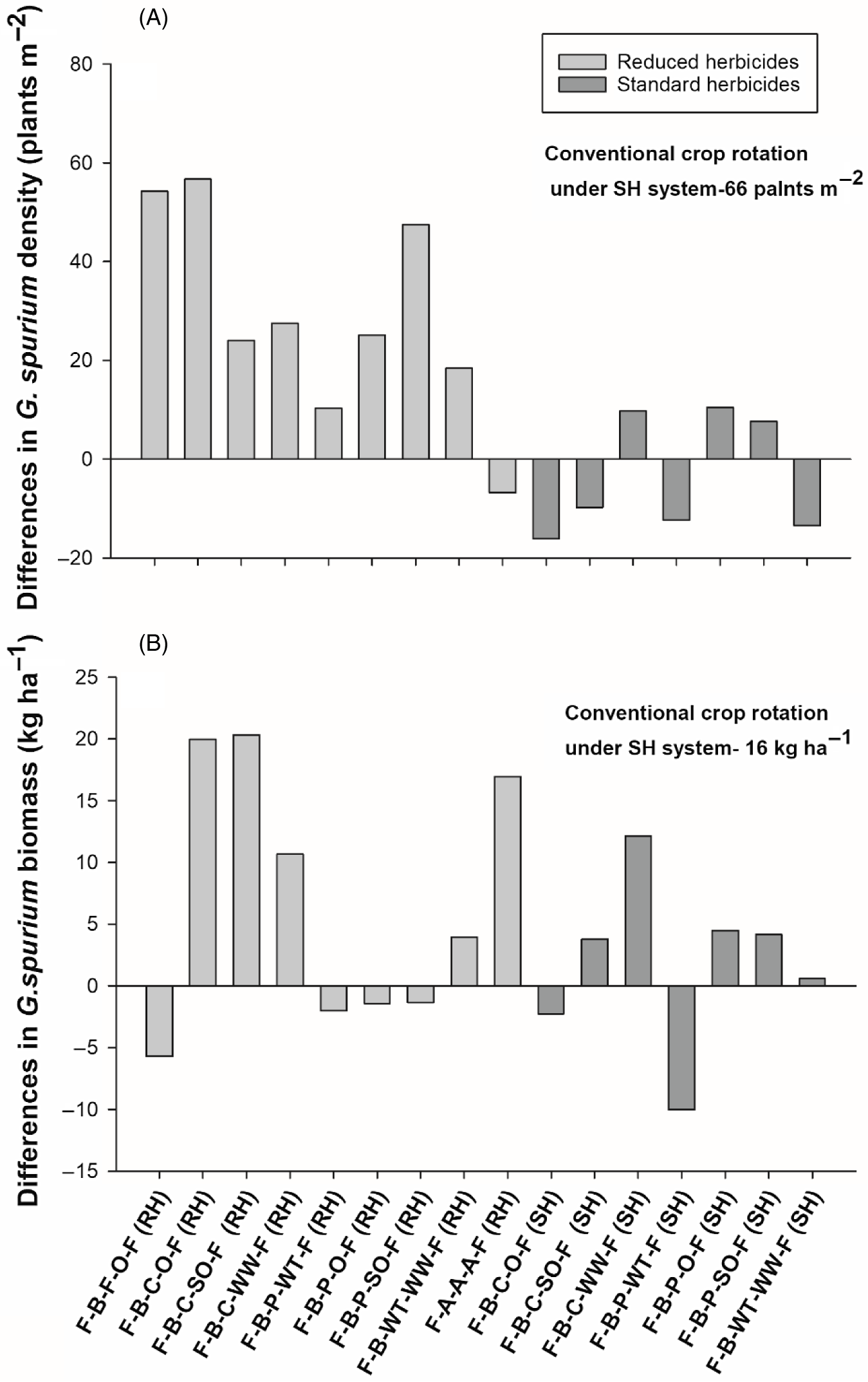
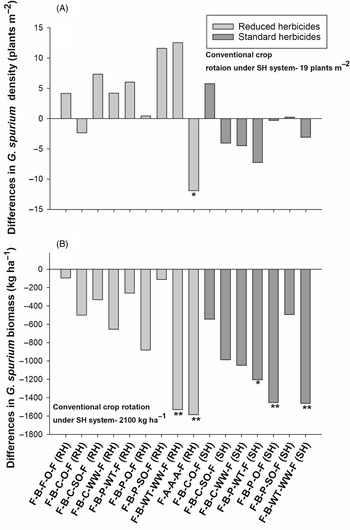
Figure 7. The effect of crop rotation on (A) Galium spurium density and (B) G. spurium biomass at Indian Head, SK, Canada. Bars represent mean differences between the conventional crop rotation F-B-F-O-F (SH) and other crop rotations. ST, spring triticale; SW, spring wheat; F, flax; A, alfalfa; B, barley; O, oat; WT, winter triticale; WW, winter wheat; SO, silage oat; P, pea; C, canola; RH, reduced herbicides; SH, standard herbicides.
Overall, in this study we observed improved weed control (G. spurium and A. fatua) by including 3 yr of alfalfa in a crop rotation centered around flax production. Meiss et al. (Reference Meiss, Le Lagadec, Munier-Jolain, Waldhardt and Petit2010) also noted better control of these two weed species in a perennial alfalfa rotation. While other studies demonstrated better A. fatua control with perennial alfalfa (Benaragama et al. Reference Benaragama, Leeson and Shirtliffe2019; Entz et al. Reference Entz, Bullied and Katepa-Mupondwa1995; Harker et al. Reference Harker, O’Donovan, Turkington, Blackshaw, Lupwayi and Smith2016), ours is the first study to examine its inclusion to minimize weed densities before a flax crop under both RH and SH systems. Gulden et al. (Reference Gulden, Lewis, Froese, Van Acker, Martens, Entz, Derksen and Bell2011) reported that after 10 yr in a rotation of flax–oat–alfalfa–alfalfa, the weed seedbank was reduced even when omitting herbicides in flax and oat. This study also shows that alfalfa is an essential crop in reducing the weed population over the long-term.
The improved weed control due to alfalfa could be due to a number of factors, including reduced soil disturbance, increased competition, reduced weed seedbank addition, or even enhanced weed seed predation (Chung and Miller Reference Chung and Miller1995; De Heij and Willenborg Reference De Heij and Willenborg2020; Meiss et al. Reference Meiss, Le Lagadec, Munier-Jolain, Waldhardt and Petit2010; Ominski et al. Reference Ominski, Entz and Kenkel1999). Reduced weed seed production due to intense competition and reduced weed seed shed due to early harvesting for hay production are traits common to alfalfa crops and could well be the mechanisms responsible for improved A. fatua control identified in this study. Yet surprisingly, similar results were not observed for the silage crop, indicating both the timing and number of forage harvesting events have a significant impact on future A. fatua populations. In fact, much of the impact of timing of forage harvest is a function of seed shed timing of weedy species. In contrast to A. fatua, seed retention of G. spurium is variable but generally greater than A. fatua (Burton et al. Reference Burton, Beckie, Willenborg, Shirtliffe, Schoenau and Johnson2016; Tidemann et al. Reference Tidemann, Hall, Harker, Beckie, Johnson and Stevenson2017). Therefore, reduced seed shed due to early harvesting may not be the primary mechanism of reduced G. spurium biomass and density in the rotation that contained alfalfa. Hence, having an annual silage crop (alternative to perennial crop) in the rotation may not be as beneficial for G. spurium compared with A. fatua. However, because G. spurium forms a tangling vine, cutting alfalfa for hay could have a supplementary impact on controlling this species. Moreover, the continuous ground cover provided by alfalfa may exert competition, thereby reducing seed production. It may also inhibit germination of G. spurium, thereby decreasing the G. spurium population over time. This is important, because G. spurium exhibits variation in emergence timing within and between years and often has multiple flushes (Defelice Reference Defelice2002; De Roo Reference De Roo2016; Royo-Esnal et al. Reference Royo-Esnal, Torra, Conesa and Forcella Recasens2010). These traits collectively make G. spurium challenging to control with the timing of in-crop herbicides, but the continuous cover provided by perennial crops is likely to improve long-term management of G. spurium in prairie crop rotations.
Winter cereal crops in rotation with summer crops break the life cycle of summer annual weeds by incorporating diverse crop planting and harvesting dates. Alternating between winter and summer crops has been found to reduce weed densities in other studies (Blackshaw Reference Blackshaw1994; Thomas and Frick Reference Thomas and Frick1993). Our study identified that having a single winter crop in the rotation did not contribute to the improved management of A. fatua or G. spurium. However, having two consecutive winter crops did show some promise for the control of both G. spurium and A. fatua at Carman, which was the only site where good overwintering occurred. These results are congruent with those of Anderson (Reference Anderson2005, Reference Anderson2008), who showed that weed control was more profound when two consecutive warm-season crops were followed by two successive winter crops. Our results, combined with those of Anderson (Reference Anderson2005, Reference Anderson2008), support the idea that the stacking of crop phases (Derksen et al. Reference Derksen, Anderson, Blackshaw and Maxwell2002) will be more beneficial than rotating crops every season (Garrison et al. Reference Garrison, Miller, Ryan, Roxburgh and Shea2014). Winter cereals can be beneficial in weed management due to their early establishment during spring and the ability to compete better than spring cereals (Beres et al. Reference Beres, Harker, Clayton, Bremer, Blackshaw and Graf2010; Entz and Fowler Reference Entz and Fowler1991). However, we used a 2X seeding rate for all winter cereal crops in this study, which might have made them more competitive. We did this because poor establishment due to winterkill is common in winter cereals (Beres et al. Reference Beres, Harker, Clayton, Bremer, Blackshaw and Graf2010; Lafond and Fowler Reference Lafond and Fowler1989), and it was likely the reason for the failure of these crops at Kernen and Indian Head. Among the three sites, only Carman and Indian Head had crop rotations with a single-season winter crop established. Yet at both of these sites, crop rotations with a single-season winter crop did not provide better A. fatua management than the conventional crop rotation.
Crop rotations containing different summer annual crops, including grain oat, spring wheat, and silage oat tended to show inconsistent A. fatua control compared with the conventional crop rotation. Perhaps more interesting is that we observed no effect on A. fatua management due to the use of silage crops in the rotation when compared with summer annual grain crops, winter crops, or alfalfa crops. This contrasts with the findings of Harker et al. (Reference Harker, Kirkland, Baron and Clayton2003), who showed substantial reductions in A. fatua density could be achieved by growing silage barley compared with barley grown for grain. The lack of impact that silage crops had on controlling A. fatua in the current study could be due to numerous factors. First, to minimize A. fatua seed return to the seedbank, silage harvesting would have to be carried out before A. fatua seed shed. Differences in accumulated growing degree days and higher growing season temperatures can cause A. fatua to mature early and shed seeds before they can be removed with the silaging operation (Shirtliffe et al. Reference Shirtliffe, Entz and Van Acker2000). Moreover, multiple cuts of alfalfa are taken each year, often for several years, whereas silage oat is only harvested only once annually, and that operation would need to be timed before A. fatua seed shatter in order to impact A. fatua populations. Second, the silage phase of the rotation occurred in the fourth year of the rotation, and thus its weed control benefit may not have been detected immediately in the following year, which was the fifth and final year of the study. Avena fatua is known to have dormancy characteristics (Beckie et al. Reference Beckie, Francis and Hall2012; Sharma et al. Reference Sharma, McBeath and Vanden Born1976) that may cause difficulties in the immediate detection of weed seed reduction by cultural strategies. It is nevertheless possible that the benefit of A. fatua seedbank depletion offered by silage crops (Harker et al. Reference Harker, O’Donovan, Turkington, Blackshaw, Lupwayi and Smith2016) could possibly be recognized in longer rotation studies. Other studies that have reported positive results of silage crops on A. fatua included early-cut silage treatments for three consecutive years (Harker et al. Reference Harker, Kirkland, Baron and Clayton2003) or in two alternate years (Harker et al. Reference Harker, O’Donovan, Turkington, Blackshaw, Lupwayi and Smith2016), which differed from the treatment regime in the current study. Our study included only a single silage crop, because rotations in which silage is cut early for 3 yr in a row or is grown in two alternate years of rotation are not common on the prairies and are unlikely to be adopted by grain growers.
None of the other crop diversification strategies (rotating canola, peas, and oat) were identified as comparatively effective in managing A. fatua or G. spurium. As summer annual crops, these crops did not add substantial functional diversity to the rotations, thus highlighting the importance of increasing functional diversity in crop rotations for reduced herbicide reliance in weed management.
Avena fatua and G. spurium are some of the most important weed species in the prairie cropping systems and are becoming increasingly problematic due to increased herbicide resistance. In this study, we identified that crop rotation with 3 yr of an alfalfa crop (flax–barley–alfalfa–alfalfa–alfalfa–flax) was the most consistent in managing both A. fatua and G. spurium with reduced herbicides over the long term. However, alfalfa may not be attractive for grain growers due to a reduction in cash flow during the 3 yr the alfalfa is grown (Smith et al. Reference Smith, Harker, O’Donovan, Turkington, Blackshaw, Lupwayi, Johnson, Pageau, Shirtliffe, Gulden and Hall2018), unless there is a demand for hay in the region. Nevertheless, our results show it can have a marked impact on weed populations before growing flax, a crop for which few herbicide options exist. The alternative crop rotations we tested, such as the inclusion of winter cereals, were found to be promising, particularly when two winter crops were included in the rotation. However, the addition of a silage crop in the rotation did not produce better weed management compared with the standard flax–barley–flax–oat–flax rotation with standard herbicides. The inability to detect better weed management benefits in rotation containing silage oat does not preclude using silage crops in rotations, however, as the effect of silage crops in reducing weed seedbanks may not have been realized within the study period.
Acknowledgments
We would like to sincerely thank all technical staff involved with this project for their help and expertise, including Gerry Stuber and Aaron Gerein at Kernen, Orla Willoughby at Indian Head, and Derek Lewis at Carman. This work was funded by the Growing Forward Canola/Flax Agri-Science Cluster. No conflicts of interest have been declared.





 Open access
Open access