



Introduction
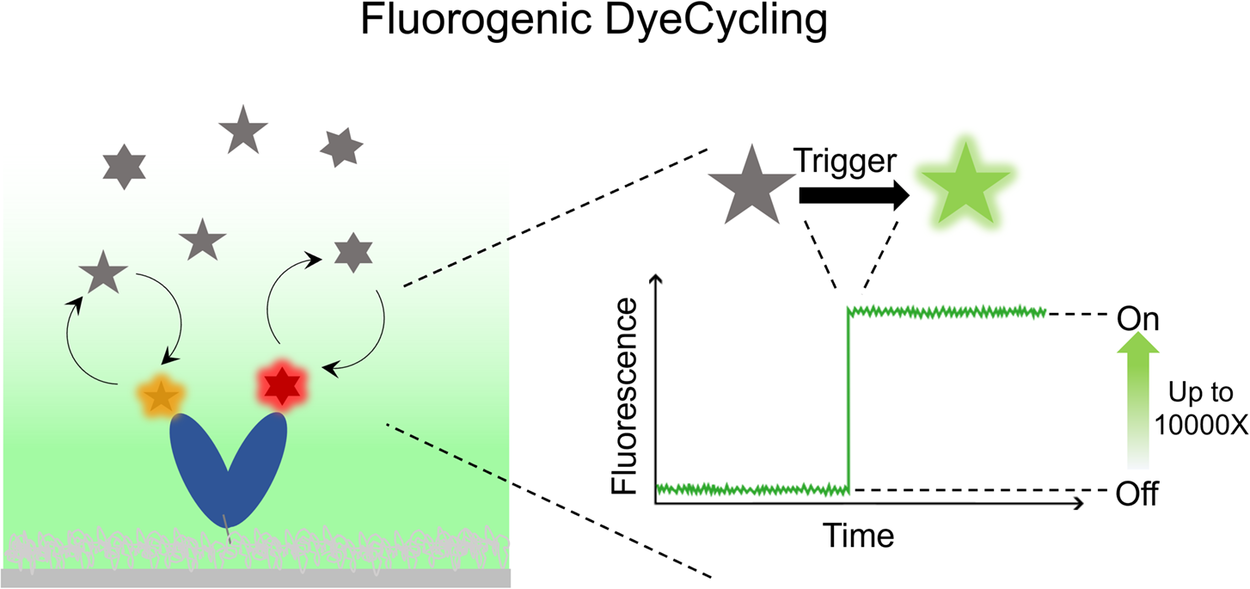
Fluorogenicity is the ability to transform the state of a fluorophore from a ‘dark’ to a ‘bright’ state, usually caused by a trigger event, such as target binding or a laser pulse, which changes the fluorophore structure and/or its local environment (Figure 1). A critical number is the fluorogenic ratio, i.e., the ratio of the fluorescence of the high-emission to low-emission forms, which can reach up to four orders of magnitude (Kozma and Kele, Reference Kozma and Kele2019). In a range of sensing and imaging works, fluorogenicity has been a valuable tool to enhance measurement contrast by suppressing non-specific fluorescence background and the noise resulting from it. For example, Liu et al. detect drug-induced proteome stress using a protein aggregation-specific fluorogenic probe, an improvement over prior cytotoxic assays (Liu et al., Reference Liu, Fares, Dunham, Gao, Miao, Jiang, Bollinger, Boal and Zhang2017). Aggregation Induced emission (AIE) has been used to screen the effectiveness of antibiotics against bacterial growth, laying the foundation for personalized medicine (Zhao et al., Reference Zhao, Chen, Chen, Deng, Gui, Leung, Hong, Lam and Tang2015). Molecular beacons (MBs) have been widely used since their introduction in 1996 (Tyagi and Kramer, Reference Tyagi and Kramer1996), for example in monitoring PCR (Wang and Yang, Reference Wang and Yang2013), as pathogen diagnostic tools (Abravaya et al., Reference Abravaya, Huff, Marshall, Merchant, Mullen, Scheider and Robinson2003), and recently in super-resolution microscopy via DNA-PAINT (Kim and Li, Reference Kim and Li2023). Fluorogenic RNA aptamers allow for precise detection and quantification of RNA production (Lu et al., Reference Lu, Kong and Unrau2023). In microscopy applications, fluorogenicity allows the observation of target-bound probes over freely diffusing ones at high contrast, thus enabling wash-free imaging (Werther et al., Reference Werther, Yserentant, Braun, Grußmayer, Navikas, Yu, Zhang, Ziegler, Mayer, Gralak, Busch, Chi, Rominger, Radenovic, Liu, Lemke, Buckup and Wombacher2021).

Figure 1. Schematic fluorogenic behaviour: a trigger event transforms the fluorophore from a dark to a brightly-emitting state. Fluorogenic ratios (bright to dark) of 10,000:1 can be achieved (Kozma and Kele, Reference Kozma and Kele2019).
Single-molecule Förster resonance energy transfer (smFRET) is used widely for studying biomolecular dynamics in the range of 2–8 nm (Ha et al., Reference Ha, Fei, Schmid, Lee, Gonzalez, Paul and Yeou2024). In its conventional implementation, a donor and acceptor fluorophore pair is site-specifically coupled to the biomolecule of interest. The energy transfer efficiency (E) from the donor to the acceptor results from the interaction of the donor and acceptor transition dipoles and scales with the inter-fluorophore distance (R) as E = (1 + (R/R 0)6)−1, where the pair-specific constant R 0 is the Förster radius at which E = 0.5. Experimentally, E can be obtained from the individual donor and acceptor fluorescence intensities as E = I A/(I D + I A), subject to experiment-specific corrections (Hellenkamp et al., Reference Hellenkamp, Schmid, Doroshenko, Opanasyuk, Kühnemuth, Rezaei Adariani, Ambrose, Aznauryan, Barth, Birkedal, Bowen, Chen, Cordes, Eilert, Fijen, Gebhardt, Götz and Hugel2018). Thanks to its strong nanoscale distance dependence, smFRET is sometimes called ‘a molecular ruler’. Indeed, it can reveal conformational changes of biomolecules in real-time, providing spatio-temporal information that is missed by structural biology techniques like cryo-electron microscopy or X-ray diffraction. Moreover, owing to its single-molecule resolution, smFRET is capable of resolving steady-state dynamics in- and out-of-equilibrium as well as static sample heterogeneity (Lerner et al., Reference Lerner, Barth, Hendrix, Ambrose, Birkedal, Blanchard, Börner, Chung, Cordes, Craggs, Deniz, Diao, Fei, Gonzalez, Gopich, Ha, Hanke and Boudker2021). In the experiment, single-molecule resolution is achieved with microscope designs that minimize background fluorescence and associated noise by exciting only a small sample volume, which is realised most commonly using confocal or total internal reflection (TIR) fluorescence microscopes (Ha et al., Reference Ha, Fei, Schmid, Lee, Gonzalez, Paul and Yeou2024). To monitor functional dynamics of a given protein over seconds or minutes, the biomolecule of interest is often tethered to a passivated surface, where hundreds to thousands of single molecules can be recorded in parallel using wide field detection by s-CMOS or EM-CCD cameras in TIR microscopes. Over the past three decades, smFRET has rapidly evolved from a niche technique to a widely popular approach to answering fundamental questions in biology (Ha et al., Reference Ha, Fei, Schmid, Lee, Gonzalez, Paul and Yeou2024).
Still, surface-tethered smFRET experiments are fundamentally limited by irreversible photobleaching of the covalently labelled fluorophores (Figure 2A). Each organic fluorophore molecule, used as FRET donor or acceptor, has a finite photon budget before its photoinduced degradation or photobleaching (Zheng and Lavis, Reference Zheng and Lavis2017). This fact forces the experimenter to choose between obtaining a high signal-to-noise ratio (SNR) with a high excitation laser power leading to faster bleaching or a longer observation time with a low laser power and thus low SNR. Currently, the achieved temporal bandwidth in smFRET literature (spanning from the shortest to the longest measurable time interval) spans only 2–3 orders of magnitude in time (Vermeer and Schmid, Reference Vermeer and Schmid2022). Despite efforts in developing more photostable self-healing fluorophores (Altman et al., Reference Altman, Terry, Zhou, Zheng, Geggier, Kolster, Zhao, Javitch, Warren and Blanchard2012; Isselstein et al., Reference Isselstein, Zhang, Glembockyte, Brix, Cosa, Tinnefeld and Cordes2020; Pati et al., Reference Pati, El, Jockusch, Zhou, Altman, Fitzgerald, Asher, Terry, Borgia, Holsey, Batchelder, Abeywickrama, Huddle, Rufa, Javitch, henrik and Blanchard2020; Zheng et al., Reference Zheng, Juette, Jockusch, Wasserman, Zhou, Altman and Blanchard2014) and the development of photo-stabilizers, such as oxygen scavengers (Benesch and Benesch, Reference Benesch and Benesch1953; Aitken et al., Reference Aitken, Marshall and Puglisi2008; Swoboda et al., Reference Swoboda, Henig, Cheng, Brugger, Haltrich, Plumeré and Schlierf2012) and triplet state quenchers (Rasnik et al., Reference Rasnik, McKinney and Ha2006; Roy et al., Reference Roy, Hohng and Ha2008; Vogelsang et al., Reference Vogelsang, Kasper, Steinhauer, Person, Heilemann, Sauer and Tinnefeld2008), eventual photobleaching leads to an unwanted, early end of the smFRET experiment. Several biomolecular processes occur on timescales beyond the current observation limit of 2–3 orders of magnitude in time (Vermeer and Schmid, Reference Vermeer and Schmid2022). For example, the bacterial RNA polymerase makes processing pauses of tens to hundreds of seconds (Janissen et al., Reference Janissen, Eslami-Mossallam, Artsimovitch, Depken and Dekker2022) before continuing RNA polymerization, which are timescales that are out of reach for conventional smFRET experiments with reasonable time resolution of milliseconds. Also, ribosomal translation observed in smFRET experiments (with one covalent donor and multiple acceptors in a row) was limited to the detection of only 12 amino acid transfers (Tsai et al., Reference Tsai, Puglisi and Uemura2016), while the median human protein consists of 375 amino acids (Brocchieri and Karlin, Reference Brocchieri and Karlin2005), converting to >1 h translation time in such room temperature experiments. As a result, co-translational effects and many other biological processes remain far beyond the accessible timescales of today’s smFRET experiments. In particular, rare but decisive events, such as bursting and pausing, are hard or impossible to study by conventional smFRET and may even be discarded as ‘odd’ outlier events, while magnetic tweezers (Janissen et al., Reference Janissen, Eslami-Mossallam, Artsimovitch, Depken and Dekker2022), nanopores (Nova et al., Reference Nova, Craig, Mazumder, Laszlo, Derrington, Noakes, Brinkerhoff, Yang, Vahedian-Movahed, Li, Zhang, Bowman, Huang, Mount, Ebright and Gundlach2024), and fluorescent product detection (English et al., Reference English, Min, Van Oijen, Kang, Luo, Sun, Cherayil, Kou and Xie2006) demonstrate their prevalence in protein function. Also, the observations of long-lived dynamic disorder and large fluctuations in enzyme catalytic rates (van Oijen et al., Reference van Oijen, Blainey, Crampton, Richardson, Ellenberger and Xie2003) emphasize the importance of a per-molecule analysis that DyeCycling aims to provide.

Figure 2. Conventional smFRET vs DyeCycling. (A) Conventional smFRET illustrated with the covalently attached donor (yellow) and acceptor (red) fluorophores bound to the biomolecule under study. Schematic FRET time traces (blue) illustrate the short observation time of conventional smFRET due to irreversible photobleaching of the donor or the acceptor fluorophore. DA: FRET-sensitized acceptor emission after donor excitation; DD: donor emission after donor excitation; AA: acceptor emission after acceptor excitation; (B) Cartoon of DyeCycling with reversibly binding donor and acceptor fluorophores in solution. The schematic time traces illustrate that DyeCycling is not limited by photobleaching of individual fluorophores and can theoretically continue for hours. The grey shaded areas represent pauses with incomplete FRET pairs (donor and/or acceptor missing). The typical sampling rate of 100 ms is assumed here.
Moreover, current analysis approaches that combine data of many molecules to extract global kinetic models (the best option currently, some are listed in Table 1 in Lerner et al., Reference Lerner, Barth, Hendrix, Ambrose, Birkedal, Blanchard, Börner, Chung, Cordes, Craggs, Deniz, Diao, Fei, Gonzalez, Gopich, Ha, Hanke and Boudker2021) rely on the ergodic assumption, while Thirumalai and co-workers showed effective ergodicity breaking already for simple DNA-based Holliday junctions (Hyeon et al., Reference Hyeon, Lee, Yoon, Hohng and Thirumalai2012). Their findings hint at even more such complexity in protein systems. Hence, pooling many single-molecule traces together for a global analysis remains a non-ideal workaround, as long as the ergodic assumption is not verified for the studied dataset. Lastly, also from an energy perspective, the currently achieved temporal bandwidth of smFRET (2–3 orders of magnitude in time) is suboptimal. This bandwidth limits the maximum equilibrium constant (K) and hence Gibbs free energy that can be probed with conventional smFRET. For example, it translates to approximately 1/3–1/2 of the energy provided by ATP hydrolysis (ca. 30 kJ/mol depending on buffer conditions) (Rosing and Slater, Reference Rosing and Slater1971), as obtained by dG = − RTln(K) with T = 300 K, equilibrium constant K = k 1/k 2 = 100 to 1000 , where k 1 and k 2 are the fastest and slowest measurable processes within the bandwidth, respectively. Notably, this general thermodynamic statement holds for conventional smFRET experiments with various time resolutions.
DyeCycling can overcome these issues (Figure 2B): using reversible binding and dissociation of the fluorophores (or “dyes”), it decouples the observation of a given biomolecule from the photobleaching of just one fluorophore (Vermeer and Schmid, Reference Vermeer and Schmid2022) – a trick that has previously revolutionized super-resolution imaging through Point Accumulation for Imaging in Nanoscale Topography (PAINT) (Sharonov and Hochstrasser, Reference Sharonov and Hochstrasser2006; Jungmann et al., Reference Jungmann, Steinhauer, Scheible, Kuzyk, Tinnefeld and Simmel2010). DyeCycling, however, is not an imaging technique: instead of images with high resolution in space, DyeCycling resolves (bio-)molecular dynamics in time. More precisely, it broadens the temporal bandwidth of smFRET experiments, such that previously inaccessible biomolecular processes can be studied, e.g., the fast processing of RNA-polymerase interspaced by long pausing (Janissen et al., Reference Janissen, Eslami-Mossallam, Artsimovitch, Depken and Dekker2022). While the concept of DyeCycling was initially demonstrated using reversible DNA hybridization of fluorescently labelled cycler oligos to ssDNA docking strands coupled to the biomolecule of interest (similar to Renewable Emission via Fluorogenic and Repeated ssDNA Hybridisation, REFRESH-FRET (Kümmerlin et al., Reference Kümmerlin, Mazumder and Kapanidis2023)), technically every reversible coupling chemistry with suitable binding kinetics can be used. Two points are thereby key for the DyeCycling experiment: First, relatively high concentrations (ca. 1 μM) of free-floating fluorescent cyclers are needed for fast binding rates (ca. 1/s), in order to achieve nearly 100% cycler-pair-bound coverage and minimize unbound ‘blind gaps’. Second, as a result, good background suppression is crucial to keep the noise level low and achieve smFRET trajectories with good SNR. The upper limit of tolerable fluorophore concentrations in solution for typical TIR setups is ≤100 nM. Zero-mode waveguides (ZMWs) present a viable physical solution (Levene et al., Reference Levene, Korlach, Turner, Foquet, Craighead and Webb2002; Crouch et al., Reference Crouch, Han and Bohn2018) for background suppression in DyeCycling experiments, as will be published elsewhere. This is because they offer particularly small excitation volumes in the zeptoliter range (Yang et al., Reference Yang, Klughammer, Barth, Tanenbaum and Dekker2023). However, ZMWs require nanofabrication know-how and equipment, while reliable commercial solutions are costly or even lacking.
Here, we explore fluorogenicity as an alternative chemical solution for background suppression in DyeCycling experiments (Figure 3A). Since fluorogenic cyclers would only become bright upon specific target binding, causing minimal background fluorescence when unbound, fluorogenic DyeCycling would facilitate high-quality smFRET recordings using common, cheap microscopy slides or coverslips. In this perspective paper, we discuss various fluorogenic systems (Figure 3B) with a focus on their potential for DyeCycling-based smFRET experiments, the requirements of which are summarized in Box 1. More general fluorogenicity reviews covering more physico-chemical mechanisms exist already elsewhere (Kozma and Kele, Reference Kozma and Kele2019; Li et al., Reference Li, Tebo and Gautier2017b; Lu et al., Reference Lu, Kong and Unrau2023).

Figure 3. (A) Schematic representation of fluorogenic DyeCycling, where the donor and acceptor fluorophores are dark in solution but become fluorescent upon binding to the molecule of interest. Donor and acceptor are continuously replaced with the respective fluorophores in solution. (B) Several fluorogenic mechanisms exist to turn a dark fluorogen into a brightly fluorescent one. We categorize them into: removal of a quencher, modulation of the fluorogen’s core structure, modulation of its local environment, or binding to a biological ligand. Alternatively, apparent fluorogenicity results from energy transfer to another bright molecule.
Box 1: Requirements for fluorogenic DyeCycling probes
-
✓ A strong fluorogenic ratio of ≥10 is desired to efficiently suppress the background fluorescence and noise of unbound cyclers in solution (Kozma and Kele, Reference Kozma and Kele2019).
-
✓ Optimal photophysics: high photostability, minimal blinking, ideally a high extinction coefficient ε > 50,000 M−1 cm−1 (Ha et al., Reference Ha, Fei, Schmid, Lee, Gonzalez, Paul and Yeou2024) and ideally a quantum yield QY > 0.6, and minimal cross-talk between donor and acceptor fluorophores are important criteria for fluorogenic systems compliant with DyeCycling.
-
✓ FRET characteristics include further a sufficiently large overlap integral between donor emission and acceptor excitation spectra and no interference of the fluorogenic systems of the donor and acceptor with FRET.
-
✓ Small size and short linkers: precise inter-fluorophore distance determination by smFRET ideally requires small point probes with short linkers (i.e., small fluorophore-accessible volumes), yet ample orientational freedom. Large probes can sterically affect biomolecular dynamics, hence, small organic probes (~1 nm) are generally preferred.
-
✓ Water-solubility of the fluorogenic probe is required for DyeCycling in aqueous buffers. Coupling auxiliary hydrophilic groups (even DNA) to the fluorogenic species can improve solubility.
-
✓ Suitable binding kinetics: dissociation rates faster than the bleach rate and ideally diffusion-limited binding rates enable minimal blind gaps and maximal observation time in DyeCycling experiments.
-
✓ Site-specific binding of the donor as well as the acceptor probe to the biomolecule of interest at the desired position is necessary and can be achieved through a range of bioconjugate techniques.
Altogether, the requirements for fluorogenic DyeCycling are more stringent than for (super-resolution) imaging applications.
Fluorogenic mechanisms and their applications
Removal of a quencher
Fluorophore-quencher pairs are popular fluorogenic systems, where the fluorophore is dark when a quencher molecule is nearby, and the fluorescence is restored by removal of this quencher. Quenching can occur via several distance-dependent mechanisms (Crisalli and Kool, Reference Crisalli and Kool2011): through energy transfer (similar to FRET) if the quencher is within 2–10 nm of the fluorophore, or through shorter-range quenching if they are <2 nm apart. These short-range mechanisms include static contact quenching (e.g., by π–π stacking), dynamic collisional quenching in the excited state, photoinduced electron transfer (PET) driven by redox chemistry, and Dexter electron transfer via spectral overlap between the electron donor and acceptor (Goldberg et al., Reference Goldberg, Batjargal, Chen and Petersson2013).
A molecular beacon (MB) is a single-stranded DNA (ssDNA) probe with a fluorophore and a quencher at its two ends, which forms a hairpin structure causing close end-to-end proximity and thus fluorescence quenching. Binding to a complementary ssDNA target disrupts the hairpin, thereby separating quencher and fluorophore in space, which de-quenches the fluorescence. For super-resolution (PAINT) imaging, an MB-PAINT was designed with a target sequence (9 nt) partially embedded in the stem and partially exposed in the loop (Figure 4A) to accelerate target binding over conventional MB (Kim and Li, Reference Kim and Li2023). Nevertheless, MB-PAINT binding remained ca. 70-fold slower compared to unhindered hybridization, due to the competition with the intramolecular hairpin – a disadvantage for potential use in DyeCycling. Alternatively, longer (15 nt) ssDNA with fluorophore and quencher (Cy3B/BHQ2 and ATTO 643/IBFQ) at opposite ends were used for fluorogenic DNA-PAINT where mismatches were introduced to facilitate reversible binding (Chung et al., Reference Chung, Zhang, Kidd, Zhang, Williams, Rollins, Yang, Lin, Baddeley and Bewersdorf2022). This approach was also used in REFRESH-FRET (Kümmerlin et al., Reference Kümmerlin, Mazumder and Kapanidis2023), using the fluorophore-quencher pair Cy3B-BHQ2 or using contact quenching of two ATTO 647N molecules. While the latter tolerates shorter ssDNA strands (i.e., favourable faster dissociation), the two bright ATTO 647N molecules, once de-quenched, complicate the smFRET readout (increased noise, Homo-FRET, etc.).

Figure 4. Examples of fluorogenic systems. (A) The molecular beacon (MB) and its application for MB-PAINT. Upon binding to the docking strand, the fluorophore and quencher separate in space, leading to increased fluorophore emission. Image from Kim and Li. Reference Kim and Li2023. Reprint permission obtained from John Wiley & Sons – Books. (B) Reversible binding of ligands to HaloTag7. The chemical structures of the ligands are shown on the bottom. Image from Kompa et al. (Reference Kompa, Bruins, Glogger, Wilhelm, Frei, Tarnawski, D’Este, Heilemann, Hiblot and Johnsson2023). Reprint under OpenAccess with CC BY 4.0. (C) The fluorophore-binding aptamer (Rhodamine Binding Aptamer for Super-resolution Imaging Techniques – RhoBAST) with a contact-quenched fluorophore–quencher (F–Q) conjugate. The RhoBAST RNA sequence and the structure of the F–Q (tetramethylrhodamine (TMR) – dinitroaniline (DN)) are shown. Image from Sunbul et al. (Reference Sunbul, Lackner, Martin, Englert, Hacene, Grün, Nienhaus, Nienhaus and Jäschke2021). Reprint permission obtained from Springer Nature. (D) Fluorogenicity by energy transfer via DRET. (i) No donor emission is observed in ssDNA (dark). The structure of the dark donor is shown on the right. (ii) Weak donor emission in dsDNA with quantum yield (QY) ~ 0.05. (iii) The acceptor (e.g., Cy5, ATTO 647N) is not excited by the donor excitation wavelength. (iv) Efficient DRET occurs in dsDNA with donor and acceptor on complementary strands.
In general, for energy transfer-based quenchers, the fluorophore-quencher separation required for efficient dequenching (ca. 2*R 0 with R 0 = 3.5–7.5 nm (Le Reste et al., Reference Le Reste, Hohlbein, Gryte and Kapanidis2012)) imposes a lower limit for the duplex length (7–15 nm or 21–45 nt), and thus an upper limit for the dissociation rate, which can only be increased to some extent by base-pair mismatches. Hence, short-range quenching mechanisms appear promising in this regard: e.g., contact quenching using dinitroaniline, trinitroaniline, or carbazole (Sunbul and Jäschke, Reference Sunbul and Jäschke2013), or photo-induced electron transfer to tryptophan or guanine (Doose et al., Reference Doose, Neuweiler and Sauer2009).
Tetrazine groups – well-known from bio-orthogonal click chemistry (Deng et al., Reference Deng, Shen, Yu, Li, Zou, Gong, Zheng, Sun, Liu and Wu2024) – were elegantly used as quenchers in fluorogenic probes. Here tetrazine-coupled fluorophores de-quench upon reaction with a strained alkene substrate such as trans-cyclooct-2-ene (TCO) via the irreversible Inverse Electron Demand Diels – Alder (IEDDA) reaction, causing strong fluorogenicity (up to 39-fold (Beliu et al., Reference Beliu, Kurz, Kuhlemann, Behringer-Pliess, Meub, Wolf, Seibel, Shi, Schnermann, Grimm, Lavis, Doose and Sauer2019)). Quenching by tetrazine may involve resonant energy transfer (in case of spectral overlap of fluorophore emission and tetrazine absorption, ca. 520–540 nm (Pinto-Pacheco et al., Reference Pinto-Pacheco, Carbery, Khan, Turner and Buccella2020), PET from the excited fluorophore to the tetrazine (Beliu et al., Reference Beliu, Kurz, Kuhlemann, Behringer-Pliess, Meub, Wolf, Seibel, Shi, Schnermann, Grimm, Lavis, Doose and Sauer2019), as well as through-bond energy transfer (TBET, i.e., energy transfer from the fluorophore to tetrazine via a conjugated linker). Based on the latter, Loredo et al. developed a class of TBET-based photo-activatable probes for super-resolution imaging, based on coumarin, rhodamine, and BODIPY. In these systems, tetrazine undergoes photolysis, releasing nitriles and molecular nitrogen, yielding up to a 178-fold increase in BODIPY fluorescence, however only after 20 min of irradiation at 254 nm with 1400 μWcm−2 (Loredo et al., Reference Loredo, Tang, Wang, Wu, Peng and Xiao2020). Beyond the long photo-activation time and potentially damaging UV radiation, the irreversible bond formed is unsuitable for use in DyeCycling. The challenge would lie in making this strong fluorogenic probe a reversible binder that responds to longer-wavelength radiation.
Changes in the core structure of the fluorophore
The emission of an organic fluorophore can be modulated by changing its conjugated system (sometimes called its “core”), e.g., by a chemical reaction upon target binding or by a light pulse. The dynamic equilibrium of rhodamines between a non-fluorescent spirocyclic form and a fluorescent zwitterionic form was harnessed to create fluorogenic Silicon Rhodamines (SiRs). The equilibrium shifts to the fluorescent, zwitterionic form upon binding to the target, a HaloTag in this case (Si et al., Reference Si, Li, Bao, Zhang and Wang2023). Interestingly, besides the well-known covalent HaloTag ligands, Kompa et al. developed a series of exchangeable ligands that enable non-covalent, reversible labelling of the HaloTag (K D ≅ 10−8 M) with a fluorogenic ratio of 10 (Figure 4B; Kompa et al., Reference Kompa, Bruins, Glogger, Wilhelm, Frei, Tarnawski, D’Este, Heilemann, Hiblot and Johnsson2023). Also, fluorogenic SNAP-tag ligands were developed by exploiting the intramolecular cyclization of derivatives of commonly used cyanines (Cy3, Cy5, Cy7) conjugated with nucleophilic side chains. While a good fluorogenic ratio of up to 124 was achieved (Martin and Rivera-Fuentes, Reference Martin and Rivera-Fuentes2024), we are unaware of reversible SNAP-tag ligands of this kind, which precludes their application in DyeCycling. Altogether, while initially developed for imaging applications, reversible self-labelling is potentially interesting for DyeCycling, too. However, the large size of the current protein tags (HaloTag7: 34 kDa, SNAP-tag: 19.4 kDa) would significantly compromise site-specific positioning as well as the sub-nanometer resolution of smFRET (Ha et al., Reference Ha, Fei, Schmid, Lee, Gonzalez, Paul and Yeou2024; Sustarsic and Kapanidis, Reference Sustarsic and Kapanidis2015).
Light-induced fluorogenicity occurs in photoactivatable fluorophores, whose core structure can be photochemically converted from a non- or weakly emissive state into a bright emissive state. Typical implementations involve a caging group (e.g., 2-nitrobenzyl derivatives) that is removed by UV irradiation (Banala et al., Reference Banala, Maurel, Manley and Johnsson2012). Reversible photoactivation was further realized using rhodamine spiroamides that exist in a non-fluorescent ‘closed’ isomer and a highly emissive ‘open’ xanthylium isomer (λem ~580 nm). UV absorption (366 nm) by the closed isomer triggers conversion to the fluorescent open isomer via conjugation in the xanthene ring. The open isomer has a lifetime of 20–100 ms in polar solvents (Fölling et al., Reference Fölling, Belov, Kunetsky, Medda, Schönle, Egner, Eggeling, Bossi and Hell2007), until thermal conversion to the dark isomer or photobleaching takes place. Besides this very short bright life time, their utility is further limited by aggregation, non-specific adhesion, and low water solubility. Additionally, prompted by concerns about UV-induced damage to biomolecules, efforts were made to shift the photoswitching to a visible-range wavelength of 405 nm (Lee et al., Reference Lee, Rai, Williams, Twieg and Moerner2014). However, for DyeCycling, light-induced dissociation from the target molecule would be more useful than light-induced activation of all fluorophores in solution, causing unwanted background fluorescence.
Changes in the local environment of the fluorophore
Aggregation-caused quenching (ACQ) occurs through contact quenching, when certain fluorophores come into close proximity, causing strong π–π interactions through which the excited state decays via non-radiative photophysical pathways (Zhao and Chen, Reference Zhao and Chen2021). ACQ can arise due to the hydrophobic interactions in polar solvents, such as water, while the aggregates disassemble in apolar environments, generating a fluorogenic response. Intercalating fluorophores, such as amyloid-binding Thioflavin T (Hanczyc et al., Reference Hanczyc, Rajchel-Mieldzioć, Feng and Fita2021) and DNA-binding YOYO-1, work in similar ways: e.g., the dimeric YOYO-1 is non-fluorescent in water but opens upon DNA intercalation (Klymchenko, Reference Klymchenko2017), causing an over 1000-fold fluorescence increase. Conversely, aggregation-induced emission (AIE) refers to the opposite effect, where a fluorophore, quenched in solution, shows fluorescence enhancement upon aggregation. In this case, non-radiative relaxation in solution can occur through rotational and vibrational motions that get hindered upon aggregation, thus giving rise to aggregation-induced fluorogenic behaviour. For example, the tetraphenylethene is non-emissive when dissolved but becomes emissive upon aggregation, which limits the rotational freedom of its phenyl rotors and diphenylmethylene units. Such AIEgens are used in biosensing and fluorescence microscopy, where they were found to be superior to conventional organic fluorophores and quantum dots (Mei et al., Reference Mei, Leung, Kwok, Lam and Tang2015). However, the multiple closely-spaced fluorophores in ACQ and AIE are incompatible with precise single-molecule readouts, including smFRET and DyeCycling.
Binding to a protein or aptamer
Fluorogen-activating proteins (FAPs) are genetically encoded protein tags that bind specific fluorogens non-covalently, which stabilizes the emissive conformation of the fluorogen (e.g., thiazole orange, malachite green) (Li et al., Reference Li, Tebo and Gautier2017b). The resulting bright fluorogen-FAP complex was used, for example, in live cell STED imaging (Fitzpatrick et al., Reference Fitzpatrick, Yan, Sieber, Dyba, Schwarz, Szent-Gyorgyi, Woolford, Berget, Waggoner and Bruchez2009). Similarly, the Fluorescence-Activating and absorption-Shifting Tag (FAST) binds its fluorogen specifically and reversibly, through which fluorescence can be turned on or off by buffer change (±fluorogen), e.g., using microfluidics. Demonstrated FAST ligands include the green emitting 4-hydroxy-3-methylbenzylidene rhodanine (HMBR, λ abs = 481 nm, λ em = 540 nm, K D of 1.3 × 10−7 M, fluorogenic ratio of up to 550), as well as yellow and red emitting ligands (Li et al., Reference Li, Plamont, Sladitschek, Rodrigues, Aujard, Neveu, Le Saux, Jullien and Gautier2017a; Plamont et al., Reference Plamont, Billon-Denis, Maurin, Gauron, Pimenta, Specht, Shi, Quérard, Pan, Rossignol, Morellet, Volovitch, Lescop, Chen, Triller, Vriz, Le Saux and Gautier2016). A possible FRET donor is 4-hydroxy-3,5-dimethoxybenzylidene rhodanine having λ abs = 518 nm, λ em = 600 nm, K D of 9.7 × 10−7 M, and fluorogenic ratio = 220. While this is promising for DyeCycling, questions remain about its photostability (Kompa et al., Reference Kompa, Bruins, Glogger, Wilhelm, Frei, Tarnawski, D’Este, Heilemann, Hiblot and Johnsson2023). Also, the relatively big size of the FAST protein tag (14 kDa) and the limitation to amino- or carboxy-terminal labelling positions are not ideal for DyeCycling.
Alternatively, fluorogenic aptamers – here specifically RNA aptamers – bind and increase the fluorescence of their ligand. A large variety of fluorogenic RNA aptamers exist, with emission across the visible range and applications ranging from RNA sensors and transcriptional reporter arrays to super-resolution imaging of RNA (Lu et al., Reference Lu, Kong and Unrau2023). These aptamer systems function through varied fluorogenic mechanisms: intramolecular charge transfer, contact quenching, or spirocyclization. Interestingly, in some aptamers, the fluorophore molecule binds reversibly and thus can be replaced. For example, the RhoBAST (Rhodamine Binding Aptamer for Super-resolution Imaging Techniques) aptamer has the fluorophore-quencher pair of tetramethylrhodamine (TMR) and dinitroaniline (DN) as the fluorogenic ligand. TMR binding to the aptamer stops contact quenching by DN, leading to a fluorogenic ratio of 26 (Figure 4C). The K D of 1.5 ± 0.1 × 10−8 M of the RhoBAST system was found suitable for super-resolution imaging of a target RNA (Sunbul et al., Reference Sunbul, Lackner, Martin, Englert, Hacene, Grün, Nienhaus, Nienhaus and Jäschke2021). While this sounds promising for use in DyeCycling, large aptamers (55 nt for RhoBAST) have similar size limitations to the protein tags above, and the additional negative charge could impair protein activity. However, if there were smaller versions in the future (possibly from peptide nucleic acid, PNA), such minimal fluorogenic aptamers could be a promising option for DyeCycling.
Fluorogenicity via (dark) resonance energy transfer
Intermolecular dark resonant energy transfer (DRET) involves a quenched ‘dark’ donor and a resonant acceptor with corresponding spectral overlap, and it occurs like FRET via transition dipole coupling of the donor and acceptor. Barnoin et al. (Reference Barnoin, Shaya, Richert, Le, Vincent, Guérineau, Mély, Michel and Burger2021) used DialkylaminoFluoreneKetotriazolyl (DFK) as a nucleobase substitute in a DNA oligonucleotide, which served as the DRET donor (Figure 4D). Coupled to ssDNA (QY ~ 0.5%) or duplex DNA (QY ~ 5.5%), little DFK emission is observed, but in the vicinity of a suitable acceptor (e.g., Cy5, ATTO 647N), DRET occurs and the emission of the acceptor is observed. While the low donor QY naturally causes minimal donor background, which is promising for DyeCycling experiments, it also presents a drawback in comparison to FRET, where the mutual anticorrelation of donor and acceptor signals serves as a convenient internal control for data quality. DRET lacks this complementarity, as DRET-sensitised acceptor fluorescence is the only readout. This means, for the DyeCycling experiment, that low-DRET resulting from bona fide biomolecular conformations must be distinguishable from the unbound-acceptor or unbound-donor baseline to prevent their confusion and misinterpretation. This can likely be solved by choosing sufficiently close fluorophore positions, resulting in medium to high DRET. On the other hand, the high Stokes shift of DFK (~200 nm) allows one to select an acceptor causing minimal emission upon donor excitation. Also, given its apparent fluorogenic ratio of ~40 and the ability to tune the DNA binding kinetics, DRET appears as an interesting option for DyeCycling.
Discussion of fluorogenic strategies for DyeCycling
Fluorogenic probes for DyeCycling need to fulfil several requirements, as summarized in Box 1. Several of the techniques discussed in the previous sections fall short of reversibility, such as tetrazine-mediated irreversible IEDDA reactions. For protein tag-derived fluorogenic systems, reversible options exist (e.g., reversible HaloTag ligands (Kompa et al., Reference Kompa, Bruins, Glogger, Wilhelm, Frei, Tarnawski, D’Este, Heilemann, Hiblot and Johnsson2023), FAST protein (Plamont et al., Reference Plamont, Billon-Denis, Maurin, Gauron, Pimenta, Specht, Shi, Quérard, Pan, Rossignol, Morellet, Volovitch, Lescop, Chen, Triller, Vriz, Le Saux and Gautier2016)) but they are limited to end-labelling positions, and their large size (>14 kDa) makes them impractical and insensitive for nanometer-distance changes of biomolecular systems. Similarly, aggregation-caused quenching and aggregation-induced emission are impractical for single-molecule FRET as they involve multiple molecules of the same fluorophore in close proximity.
More promising fluorogenic strategies for DyeCycling are compared in Table 1. They include simple fluorophore-quencher pairs, such as DNA-based molecular beacons with quenching efficiency up to 98% (Marras et al., Reference Marras, Kramer and Tyagi2002). Mutually compatible fluorophore-quencher pairs can be selected for use as donor and acceptor, each with individual specificity for a docking strand attached to the biomolecule of interest. Care must be taken to ensure appropriate kinetics, as intra-molecular interactions may reduce the binding rate, which was previously improved by smart sequence engineering (Kim and Li, Reference Kim and Li2023). The dissociation rate, on the other hand, scales inversely with the duplex length, which allows one to tune it, e.g., by mismatch insertion (Chung et al., Reference Chung, Zhang, Kidd, Zhang, Williams, Rollins, Yang, Lin, Baddeley and Bewersdorf2022). Since quenching by resonant energy transfer can occur up to 10 nm, effective de-quenching would require relatively long duplexes, causing slow dissociation rates. Thus, short-range (1–2 nm) quenching mechanisms, such as contact, collisional, PET, or Dexter electron transfer-based quenching, appear more favourable, as they are compatible with shorter duplexes with suitable binding kinetics. RNA aptamers with reversibly binding fluorogenic ligands could make useful DyeCycling probes with tuneable binding kinetics, in particular, if small versions (possibly based on peptide nucleic acid) could be derived (Sunbul et al., Reference Sunbul, Lackner, Martin, Englert, Hacene, Grün, Nienhaus, Nienhaus and Jäschke2021). Intermolecular DRET (Barnoin et al., Reference Barnoin, Shaya, Richert, Le, Vincent, Guérineau, Mély, Michel and Burger2021), although not conventionally fluorogenic, still presents interesting characteristics for DyeCycling: DNA-based tuneable reversibility, water solubility, and high ‘apparent’ fluorogenicity of the acceptor. Hence, if the SNR of the DRET-sensitized acceptor fluorescence is sufficiently high, DRET appears as an interesting option for DyeCycling. Lastly, we note that if oligo-based cyclers are used to study RNA/DNA-containing systems, care should be taken not to introduce off-target binding.
Table 1. Comparing the potential of fluorogenic systems for DyeCycling: green: suitable, red: not optimal; D: donor, A: acceptor; Michael: Michael reaction of cysteine and maleimide; uaa: unnatural amino acid labelling. † Estimates of the contour length between fluorophore and biomolecule are specified.

Outlook
DyeCycling has the potential to offer better informed smFRET revealing biomolecular mechanisms that are currently beyond the accessible time range (e.g., polymerase pausing, co-translational effects, etc.). Fluorogenic DyeCycling would bring these benefits to regular glass (or quartz) slides, bypassing the need for nanophotonics for background suppression. The challenge in implementing fluorogenic DyeCycling lies in finding experimental conditions that meet several requirements simultaneously (Box 1). Donor and acceptor binding should be mutually orthogonal, have suitable binding kinetics (ca. 1/s binding rate, 0.1/s dissociation rate), be compatible with FRET (high overlap integral), and have excellent fluorogenic and photophysical properties (high fluorogenic ratio, brightness, and photostability). Given that the requirements of DyeCycling exceed those of typical imaging applications, it is encouraging that several existing fluorogenic systems appear compliant with fluorogenic DyeCycling. Various DNA, RNA, or PNA-based probes offer much design flexibility in terms of structure, binding kinetics, and diverse commercially available functionalization. Classical fluorophore-quencher pairs can be implemented intra-molecularly based on several short-range quenching mechanisms, which are compatible with short oligos offering suitably fast dissociation rates. Additional strategies worth investigating include small versions of fluorogenic aptamers as well as DRET combined with single-colour detection. Such fluorogenic approaches would benefit not only smFRET via DyeCycling, REFRESH-FRET (Kümmerlin et al., Reference Kümmerlin, Mazumder and Kapanidis2023) and FRET X (Filius et al., Reference Filius, Kim, Severins and Joo2021), but also super-resolution microscopy techniques (Chung et al., Reference Chung, Zhang, Kidd, Zhang, Williams, Rollins, Yang, Lin, Baddeley and Bewersdorf2022). Moreover, there is no fundamental reason limiting the development of new small-molecular and fluorogenic DyeCycling probes, which could be an interesting application of reversible covalent binders or host-guest chemistries, for example, in combination with non-canonical amino acids incorporated via genetic code expansion. Such potential future fluorogenic systems, along with the already existing options, make it very likely that fluorogenic DyeCycling can be implemented and become useful as a next-generation smFRET technique in vitro – and potentially even in cells.
Open peer review
To view the open peer review materials for this article, please visit http://doi.org/10.1017/qrd.2024.11.
Acknowledgements
We thank Pablo Rivera-Fuentes, Oliver S. Wenger, Björn Pfund, Alexandria Deliz Liang and Benjamin Vermeer for helpful discussions.
Author contribution
SS conceived the paper and SG wrote the first draft. Both discussed and edited the paper.
Financial support
This work was supported by the Swiss National Science Foundation as part of the NCCR Molecular Systems Engineering (51NF40–205608) and by the University of Basel.
Competing interest declaration
The authors declare no conflict of interest.






 Open access
Open access
Comments
Dear Editor,
Following the kind invitation by Profs. Fredrik Westerlund and Felix Ritort (cf. correspondence in Dec 2023
and May 2024), we are delighted to send you our perspective article entitled “The potential of
fluorogenicity for single molecule FRET and DyeCycling” as a contribution to the collection of articles
dedicated to “Single Molecule Challenges in the 21st Century".
Our article targets the biggest challenge in single-molecule FRET which – in its traditional form – is
fundamentally limited by photo-bleaching. To solve the problem, we recently presented the concept of
DyeCycling (https://doi.org/10.1007/s12274-022-4420-5) which uses reversibly bound fluorophores that
effectively circumvent the photo-bleaching limitation, and we now realized such DyeCycling experiments
with the help of nanophotonic devices (in preparation).
In the submitted article for QRB Discovery, we take the next logical step and present possible solutions to
democratize DyeCycling and generalize it for use in the most common FRET setups, i.e., using standard
coverslips instead of specialized nanophotonics. For that purpose, fluorogenicity offers unique advantages
and we systematically review and discuss existing fluorogenic systems, as well as their individual
strengths and weaknesses in the context of DyeCycling. Ultimately, we conclude on the most promising
fluorogenic systems for DyeCycling, and we sketch possible ways forward in the outlook section. We
believe that our article makes a valuable contribution to solving the most pressing challenge in singlemolecule FRET experiments of the 21st century.
Seite 2/3
Universität Basel
Departement Chemie
Physikalische Chemie
Mattenstrasse 22, BPR1096
4058 Basel, Schweiz
www.chemie.unibas.ch
Sonja Schmid
[email protected]
schmid.chemie.unibas.ch
For your convenience, we attach a list of suggested referees with highly relevant expertise regarding
single-molecule fluorescence techniques, and we will gladly arrange the required reprint permissions. We
hope you share our enthusiasm about the work and we are looking forward to your response.
Kind regards
Prof Sonja Schmid