



Introduction
While alcohol is widely consumed worldwide, socially accepted, and legally available in many cultures, its effects on individuals and society are heavily debated. On the one hand, low-volume alcohol consumption does not seem to affect mortality significantly compared to abstinence or occasional drinking (Stockwell et al., Reference Stockwell, Zhao, Panwar, Roemer, Naimi and Chikritzhs2016). On the other hand, alcohol consumption is robustly associated with the risk of over 200 diseases (Rehm et al., Reference Rehm, Baliunas, Borges, Graham, Irving, Kehoe, Parry, Patra, Popova, Poznyak, Roerecke, Room, Samokhvalov and Taylor2010; Roerecke and Rehm, Reference Roerecke and Rehm2012; World Health Organisation, 2014). Furthermore, 5.1% of the global burden of disease and injury, measured in disability-adjusted life years (DALYs), is attributable to alcohol consumption (World Health Organisation, 2014) with accidents and injuries causing 30% of the DALYs and various chronic diseases causing the remaining 70% of the DALYs (World Health Organisation, 2014). Most alcohol-related premature deaths are caused by the consumption of harmful levels of alcohol and alcohol dependence (AD) (Rehm et al., Reference Rehm, Shield, Gmel, Rehm and Frick2013).
Considering the potentially harmful effects of alcohol consumption, especially when consumed in large quantities (e.g. Stahre et al., Reference Stahre, Roeber, Kanny, Brewer and Zhang2014), it is important to monitor alcohol consumption behaviors. To assess problematic consumption of alcohol in the general population, current alcohol consumption is a useful metric as it allows for a comparison to the guidelines for low-risk alcohol consumption (e.g. ⩽14 alcoholic beverages per week, in most countries the threshold is lower for women than for men; CMOs, 2016, Kalinowski and Humphreys, Reference Kalinowski and Humphreys2016). It has also been proposed to replace alcohol use disorder diagnostics by simple alcohol consumption metrics with treatments of problematic or excessive alcohol consumption directed at the reduction of alcohol use (Nutt and Rehm, Reference Nutt and Rehm2014). The benefit of assessing levels of alcohol consumption by questionnaires is that it is cost-effective which allows the inclusion of larger study samples. Furthermore, fast and low-cost assessment allows for the inclusion of more people in treatment and may therefore narrow the current treatment gap for people with an alcohol use disorder (Kohn et al., Reference Kohn, Saxena, Levav and Saraceno2004; Tuithof et al., Reference Tuithof, ten Have, van den Brink, Vollebergh and de Graaf2016). The diagnostic value of alcohol consumption metrics is exemplified by the presence of considerable phenotypic correlations with, for example, AD (Kendler et al., Reference Kendler, Myers, Dick and Prescott2010). Two widely used measures to monitor alcohol consumption in the general population are ‘alcohol consumption frequency’ (i.e. the number of days on which alcohol is consumed per week) and ‘alcohol consumption quantity’ (i.e. the number of standard alcoholic drinks consumed per week).
Alcohol consumption frequency and alcohol consumption quantity have a high phenotypic correlation (based on UK Biobank data; Sudlow et al., Reference Sudlow, Gallacher, Allen, Beral, Burton, Danesh, Downey, Elliott, Green, Landray, Liu, Matthews, Ong, Pell, Silman, Young, Sprosen, Peakman and Collins2015), and have been used interchangeably in some epidemiological studies (Berggren and Sutton, Reference Berggren and Sutton1999, Heckley et al., Reference Heckley, Jarl and Gerdtham2017). However, various studies suggest that alcohol consumption quantity and alcohol consumption frequency have different and sometimes opposing effects on health. Differences in effect have been shown for cardiovascular diseases, cancers, and various mental health conditions. For example, high alcohol consumption quantity has been related to an elevated risk of all-cause mortality, cardiovascular disease, depression, and insomnia, to reduced cerebellar blood flow, and a higher risk of cancer in men. In contrast, high alcohol consumption frequency (restricted to low-to-moderate consumption quantity), has previously been related to a reduced risk of cardiovascular disease, type 2 diabetes, improved cerebellar blood flow, and an increased risk of cancer in women (Conigrave et al., Reference Conigrave, Hu, Camargo, Stampfer, Willett and Rimm2001; Stein and Friedmann, Reference Stein and Friedmann2005; Breslow and Graubard, Reference Breslow and Graubard2008; Christie et al., Reference Christie, Price, Edwards, Muldoon, Meltzer and Jennings2008; Breslow et al., Reference Breslow, Chen, Graubard and Mukamal2011; Ronksley et al., Reference Ronksley, Brien, Turner, Mukamal and Ghali2011; Piano, Reference Piano2017).
The opposing effects on the health of these alcohol consumption patterns may be explained by their differential association with specific life circumstances. For example, research suggests that people with a high socio-economic status (SES; e.g. assessed based on income and education) tend to drink more frequently and consume a larger total amount than people with a lower SES (Heckley et al., Reference Heckley, Jarl and Gerdtham2017), but consume on average less per drinking occasion. Furthermore, alcohol consumption is known to be related to a broad range of phenotypes, including substance use disorders (Grucza and Bierut, Reference Grucza and Bierut2006; Kendler et al., Reference Kendler, Myers, Dick and Prescott2010), other psychiatric disorders (Clark et al., Reference Clark, Lesnick and Hegedus1997; Kessler et al., Reference Kessler, Crum, Warner, Nelson, Schulenberg and Anthony1997; Khoury et al., Reference Khoury, Tang, Bradley, Cubells and Ressler2010), and personality/psychological traits (Hicks et al., Reference Hicks, Durbin, Blonigen, Iacono and McGue2012; Turiano et al., Reference Turiano, Whiteman, Hampson, Roberts and Mroczek2012; Hakulinen et al., Reference Hakulinen, Elovainio, Batty, Virtanen, Kivimaki and Jokela2015). The complex relation of the alcohol consumption with SES (Staff et al., Reference Staff, Patrick, Loken and Maggs2008; Sudlow et al., Reference Sudlow, Gallacher, Allen, Beral, Burton, Danesh, Downey, Elliott, Green, Landray, Liu, Matthews, Ong, Pell, Silman, Young, Sprosen, Peakman and Collins2015; Collins, Reference Collins2016; Heckley et al., Reference Heckley, Jarl and Gerdtham2017) is especially interesting, because SES itself also has a known relation with substance use disorders, psychiatric disorders, and personality/psychological traits (e.g. Hiscock et al., Reference Hiscock, Bauld, Amos, Fidler and Munafo2012; von Stumm and Plomin, Reference von Stumm and Plomin2015; Russell et al., Reference Russell, Ford, Williams and Russell2016). However, epidemiological studies that investigate associations of alcohol consumption with various other phenotypes do not provide insight into the underlying biological mechanisms which explain differential patterns of phenotypic associations for frequency and quantity of alcohol consumption.
Twin studies show that 50–60% of the phenotypic variation in alcohol consumption quantity (Swan et al., Reference Swan, Carmelli, Rosenman, Fabsitz and Christian1990), and alcohol use disorders (Mbarek et al., Reference Mbarek, Milaneschi, Fedko, Hottenga, de Moor, Jansen, Gelernter, Sherva, Willemsen, Boomsma, Penninx and Vink2015; Verhulst et al., Reference Verhulst, Neale and Kendler2015) are heritable. Furthermore, several studies have explored which genetic variants contribute to alcohol consumption quantity using Genome-Wide Association analysis (GWAS). The most recent and largest study of alcohol consumption to date has identified 14 significant loci (Clarke et al., Reference Clarke, Adams, Davies, Howard, Hall, Padmanabhan, Murray, Smith, Campbell, Hayward, Porteous, Deary and McIntosh2017). The aldehyde dehydrogenases (ALDH) and alcohol dehydrogenases (ADH) gene clusters have proven to be robust findings as they were consistently found by various alcohol consumption quantity GWAS (Schumann et al., Reference Schumann, Liu, O'Reilly, Gao, Song, Xu, Ruggeri, Amin, Jia, Preis, Segura Lepe, Akira, Barbieri, Baumeister, Cauchi, Clarke, Enroth, Fischer, Hallfors, Harris, Hieber, Hofer, Hottenga, Johansson, Joshi, Kaartinen, Laitinen, Lemaitre, Loukola, Luan, Lyytikainen, Mangino, Manichaikul, Mbarek, Milaneschi, Moayyeri, Mukamal, Nelson, Nettleton, Partinen, Rawal, Robino, Rose, Sala, Satoh, Schmidt, Schraut, Scott, Smith, Starr, Teumer, Trompet, Uitterlinden, Venturini, Vergnaud, Verweij, Vitart, Vuckovic, Wedenoja, Yengo, Yu, Zhang, Zhao, Boomsma, Chambers, Chasman, Daniela, de Geus, Deary, Eriksson, Esko, Eulenburg, Franco, Froguel, Gieger, Grabe, Gudnason, Gyllensten, Harris, Hartikainen, Heath, Hocking, Hofman, Huth, Jarvelin, Jukema, Kaprio, Kooner, Kutalik, Lahti, Langenberg, Lehtimaki, Liu, Madden, Martin, Morrison, Penninx, Pirastu, Psaty, Raitakari, Ridker, Rose, Rotter, Samani, Schmidt, Spector, Stott, Strachan, Tzoulaki, van der Harst, van Duijn, Marques-Vidal, Vollenweider, Wareham, Whitfield, Wilson, Wolffenbuttel, Bakalkin, Evangelou, Liu, Rice, Desrivieres, Kliewer, Mangelsdorf, Muller, Levy and Elliott2016; Clarke et al., Reference Clarke, Adams, Davies, Howard, Hall, Padmanabhan, Murray, Smith, Campbell, Hayward, Porteous, Deary and McIntosh2017; Jorgenson et al., Reference Jorgenson, Thai, Hoffmann, Sakoda, Kvale, Banda, Schaefer, Risch, Mertens, Weisner and Choquet2017). Moreover, the ADH cluster has also been found to play a role in AD (Gelernter et al., Reference Gelernter, Kranzler, Sherva, Almasy, Koesterer, Smith, Anton, Preuss, Ridinger, Rujescu, Wodarz, Zill, Zhao and Farrer2014a; Walters et al., Reference Walters, Polimanti, Johnson, McClintick, Adams, Adkins, Aliev, Bacanu, Batzler, Bertelsen, Biernacka, Bigdeli, Chen, Clarke, Chou, Degenhardt, Docherty, Edwards, Fontanillas, Foo, Fox, Frank, Giegling, Gordon, Hack, Hartmann, Hartz, Heilmann-Heimbach, Herms, Hodgkinson, Hoffmann, Hottenga, Kennedy, Alanne-Kinnunen, Konte, Lahti, Lahti-Pulkkinen, Lai, Ligthart, Loukola, Maher, Mbarek, McIntosh, McQueen, Meyers, Milaneschi, Palviainen, Pearson, Peterson, Ripatti, Ryu, Saccone, Salvatore, Sanchez-Roige, Schwandt, Sherva, Streit, Strohmaier, Thomas, Wang, Webb, Wedow, Wetherill, Wills, Boardman, Chen, Choi, Copeland, Culverhouse, Dahmen, Degenhardt, Domingue, Elson, Frye, Gabel, Hayward, Ising, Keyes, Kiefer, Kramer, Kuperman, Lucae, Lynskey, Maier, Mann, Mannisto, Muller-Myhsok, Murray, Nurnberger, Palotie, Preuss, Raikkonen, Reynolds, Ridinger, Scherbaum, Schuckit, Soyka, Treutlein, Witt, Wodarz, Zill, Adkins, Boden, Boomsma, Bierut, Brown, Bucholz, Cichon, Costello, De Wit, Diazgranados, Dick, Eriksson, Farrer, Foroud, Gillespie, Goate, Goldman, Grucza, Hancock, Harris, Heath, Hesselbrock, Hewitt, Hopfer, Horwood, Iacono, Johnson, Kaprio, Karpyak, Kendler, Kranzler, Krauter, Lichtenstein, Lind, McGue, MacKillop, Madden, Maes, Magnusson, Martin, Medland, Montgomery, Nelson, Nothen, Palmer, Pedersen, Penninx, Porjesz, Rice, Rietschel, Riley, Rose, Rujescu, Shen, Silberg, Stallings, Tarter, Vanyukov, Vrieze, Wall, Whitfield, Zhao, Neale, Gelernter, Edenberg, Agrawal and Team2018). Individually, single nucleotide polymorphisms (SNPs), identified by GWAS, explain a very small proportion of the variation in alcohol consumption quantity, as is the case for most complex traits (Manolio et al., Reference Manolio, Collins, Cox, Goldstein, Hindorff, Hunter, McCarthy, Ramos, Cardon, Chakravarti, Cho, Guttmacher, Kong, Kruglyak, Mardis, Rotimi, Slatkin, Valle, Whittemore, Boehnke, Clark, Eichler, Gibson, Haines, Mackay, McCarroll and Visscher2009; Clarke et al., Reference Clarke, Adams, Davies, Howard, Hall, Padmanabhan, Murray, Smith, Campbell, Hayward, Porteous, Deary and McIntosh2017). However, the SNP-based heritability of alcohol consumption quantity is estimated at 13–18% (Vrieze et al., Reference Vrieze, McGue, Miller, Hicks and Iacono2013; Clarke et al., Reference Clarke, Adams, Davies, Howard, Hall, Padmanabhan, Murray, Smith, Campbell, Hayward, Porteous, Deary and McIntosh2017), indicating that collectively GWAS SNPs explain a modest proportion of the variation in alcohol consumption quantity. Until now, no GWAS studies have been published on alcohol consumption frequency.
The reason that genetic variants that are genome-wide significantly associated with alcohol consumption only explain a modest proportion of the heritability is because alcohol consumption, like most other complex traits, has a polygenic genetic architecture. Therefore, the total heritability is distributed over thousands of variants of small effect (Visscher et al., Reference Visscher, Wray, Zhang, Sklar, McCarthy, Brown and Yang2017). A method that estimates genetic overlap by utilizing the information of all genetic variants in a GWAS, including the ones below the stringent significance threshold, is genetic correlation. Compared to epidemiological studies, which allow investigation of phenotypic correlations, genetic correlation analysis has the benefit of providing insight into the extent to which genetic risk factors are shared between traits. Previous studies which used this technique demonstrated genetic overlap between various psychiatric traits which were traditionally not seen as closely related (Anttila et al., Reference Anttila, Bulik-Sullivan, Finucane, Walters, Bras, Duncan, Escott-Price, Falcone, Gormley, Malik, Patsopoulos, Ripke, Wei, Yu, Lee, Turley, Grenier-Boley, Chouraki, Kamatani, Berr, Letenneur, Hannequin, Amouyel, Boland, Deleuze, Duron, Vardarajan, Reitz, Goate, Huentelman, Kamboh, Larson, Rogaeva, St George-Hyslop, Hakonarson, Kukull, Farrer, Barnes, Beach, Demirci, Head, Hulette, Jicha, Kauwe, Kaye, Leverenz, Levey, Lieberman, Pankratz, Poon, Quinn, Saykin, Schneider, Smith, Sonnen, Stern, Van Deerlin, Van Eldik, Harold, Russo, Rubinsztein, Bayer, Tsolaki, Proitsi, Fox, Hampel, Owen, Mead, Passmore, Morgan, Nothen, Rossor, Lupton, Hoffmann, Kornhuber, Lawlor, McQuillin, Al-Chalabi, Bis, Ruiz, Boada, Seshadri, Beiser, Rice, van der Lee, De Jager, Geschwind, Riemenschneider, Riedel-Heller, Rotter, Ransmayr, Hyman, Cruchaga, Alegret, Winsvold, Palta, Farh, Cuenca-Leon, Furlotte, Kurth, Ligthart, Terwindt, Freilinger, Ran, Gordon, Borck, Adams, Lehtimaki, Wedenoja, Buring, Schurks, Hrafnsdottir, Hottenga, Penninx, Artto, Kaunisto, Vepsalainen, Martin, Montgomery, Kurki, Hamalainen, Huang, Huang, Sandor, Webber, Muller-Myhsok, Schreiber, Salomaa, Loehrer, Gobel, Macaya, Pozo-Rosich, Hansen, Werge, Kaprio, Metspalu, Kubisch, Ferrari, Belin, van den Maagdenberg, Zwart, Boomsma, Eriksson, Olesen, Chasman, Nyholt, Avbersek, Baum, Berkovic, Bradfield, Buono, Catarino, Cossette, De Jonghe, Depondt, Dlugos, Ferraro, French, Hjalgrim, Jamnadas-Khoda, Kalviainen, Kunz, Lerche, Leu, Lindhout, Lo, Lowenstein, McCormack, Moller, Molloy, Ng, Oliver, Privitera, Radtke, Ruppert, Sander, Schachter, Schankin, Scheffer, Schoch, Sisodiya, Smith, Sperling, Striano, Surges, Thomas, Visscher, Whelan, Zara, Heinzen, Marson, Becker, Stroink, Zimprich, Gasser, Gibbs, Heutink, Martinez, Morris, Sharma, Ryten, Mok, Pulit, Bevan, Holliday, Attia, Battey, Boncoraglio, Thijs, Chen, Mitchell, Rothwell, Sharma, Sudlow, Vicente, Markus, Kourkoulis, Pera, Raffeld, Silliman, Perica, Thornton, Huckins, Rayner, Lewis, Gratacos, Rybakowski, Keski-Rahkonen, Raevuori, Hudson, Reichborn-Kjennerud, Monteleone, Karwautz, Mannik, Baker, O'Toole, Trace, Davis, Helder, Ehrlich, Herpertz-Dahlmann, Danner, van Elburg, Clementi, Forzan, Docampo, Lissowska, Hauser, Tortorella, Maj, Gonidakis, Tziouvas, Papezova, Yilmaz, Wagner, Cohen-Woods, Herms, Julia, Rabionet, Dick, Ripatti, Andreassen, Espeseth, Lundervold, Steen, Pinto, Scherer, Aschauer, Schosser, Alfredsson, Padyukov, Halmi, Mitchell, Strober, Bergen, Kaye, Szatkiewicz, Cormand, Ramos-Quiroga, Sanchez-Mora, Ribases, Casas, Hervas, Arranz, Haavik, Zayats, Johansson, Williams, Dempfle, Rothenberger, Kuntsi, Oades, Banaschewski, Franke, Buitelaar, Vasquez, Doyle, Reif, Lesch, Freitag, Rivero, Palmason, Romanos, Langley, Rietschel, Witt, Dalsgaard, Borglum, Waldman, Wilmot, Molly, Bau, Crosbie, Schachar, Loo, McGough, Grevet, Medland, Robinson, Weiss, Bacchelli, Bailey, Bal, Battaglia, Betancur, Bolton, Cantor, Celestino-Soper, Dawson, Rubeis, Duque, Green, Klauck, Leboyer, Levitt, Maestrini, Mane, Moreno-De-Luca, Parr, Regan, Reichenberg, Sandin, Vorstman, Wassink, Wijsman, Cook, Santangelo, Delorme, Roge, Magalhaes, Arking, Schulze, Thompson, Strohmaier, Matthews, Melle, Morris, Blackwood, McIntosh, Bergen, Schalling, Jamain, Maaser, Fischer, Reinbold, Fullerton, Guzman-Parra, Mayoral, Schofield, Cichon, Muhleisen, Degenhardt, Schumacher, Bauer, Mitchell, Gershon, Rice, Potash, Zandi, Craddock, Ferrier, Alda, Rouleau, Turecki, Ophoff, Pato, Anjorin, Stahl, Leber, Czerski, Cruceanu, Jones, Posthuma, Andlauer, Forstner, Streit, Baune, Air, Sinnamon, Wray, MacIntyre, Porteous, Homuth, Rivera, Grove, Middeldorp, Hickie, Pergadia, Mehta, Smit, Jansen, de Geus, Dunn, Li, Nauck, Schoevers, Beekman, Knowles, Viktorin, Arnold, Barr, Bedoya-Berrio, Bienvenu, Brentani, Burton, Camarena, Cappi, Cath, Cavallini, Cusi, Darrow, Denys, Derks, Dietrich, Fernandez, Figee, Freimer, Gerber, Grados, Greenberg, Hanna, Hartmann, Hirschtritt, Hoekstra, Huang, Huyser, Illmann, Jenike, Kuperman, Leventhal, Lochner, Lyon, Macciardi, Madruga-Garrido, Malaty, Maras, McGrath, Miguel, Mir, Nestadt, Nicolini, Okun, Pakstis, Paschou, Piacentini, Pittenger, Plessen, Ramensky, Ramos, Reus, Richter, Riddle, Robertson, Roessner, Rosario, Samuels, Sandor, Stein, Tsetsos, Van Nieuwerburgh, Weatherall, Wendland, Wolanczyk, Worbe, Zai, Goes, McLaughlin, Nestadt, Grabe, Depienne, Konkashbaev, Lanzagorta, Valencia-Duarte, Bramon, Buccola, Cahn, Cairns, Chong, Cohen, Crespo-Facorro, Crowley, Davidson, DeLisi, Dinan, Donohoe, Drapeau, Duan, Haan, Hougaard, Karachanak-Yankova, Khrunin, Klovins, Kucinskas, Keong, Limborska, Loughland, Lonnqvist, Maher, Mattheisen, McDonald, Murphy, Nenadic, van Os, Pantelis, Pato, Petryshen, Quested, Roussos, Sanders, Schall, Schwab, Sim, So, Stogmann, Subramaniam, Toncheva, Waddington, Walters, Weiser, Cheng, Cloninger, Curtis, Gejman, Henskens, Mattingsdal, Oh, Scott, Webb, Breen, Churchhouse, Bulik, Daly, Dichgans, Faraone, Guerreiro, Holmans, Kendler, Koeleman, Mathews, Price, Scharf, Sklar, Williams, Wood, Cotsapas, Palotie, Smoller, Sullivan, Rosand, Corvin, Neale and Consortium2018), and demonstrated genetic overlap between various substance use traits (Nivard et al., Reference Nivard, Verweij, Minica, Treur, Vink, Boomsma and Consortium2016). Furthermore, a study by Hill et al. (Reference Hill, Hagenaars, Marioni, Harris, Liewald, Davies, Okbay, McIntosh, Gale and Deary2016) demonstrated that heritability of SES is captured by GWAS, and showed substantial genetic correlations with various complex traits (Hill et al., Reference Hill, Hagenaars, Marioni, Harris, Liewald, Davies, Okbay, McIntosh, Gale and Deary2016). Genetic correlation analysis, therefore, also allows for the investigation of the complex relation between the quantity and frequency of alcohol consumption with SES and other complex traits, on a genetic level.
The current study aims to elucidate to what extent genetic factors are shared between quantity (in regular drinkers) and frequency of alcohol consumption. In addition, we investigate genetic correlations of alcohol consumption quantity and frequency with traits in four phenotypic categories: SES, substance use disorders, other psychiatric disorders and personality/psychological traits. These genetic correlations will provide a better insight into the etiology of different alcohol consumption behaviors and their relation to other phenotypic traits.
Methods
UK Biobank data
Data used for the alcohol consumption frequency and alcohol consumption quantity GWAS were obtained from UK Biobank (http://www.ukbiobank.ac.uk) (Sudlow et al., Reference Sudlow, Gallacher, Allen, Beral, Burton, Danesh, Downey, Elliott, Green, Landray, Liu, Matthews, Ong, Pell, Silman, Young, Sprosen, Peakman and Collins2015). The UK Biobank resource was established by the Wellcome Trust medical charity, Medical Research Council, Department of Health, Scottish Government and the Northwest Regional Development Agency. It is a major resource with the aim of improving the prevention, diagnosis and treatment of a wide range of health problems. UK Biobank recruited over 500 000 individuals aged 40–69 years between 2006 and 2010 from across the UK, and collected information on >2000 phenotypes (Sudlow et al., Reference Sudlow, Gallacher, Allen, Beral, Burton, Danesh, Downey, Elliott, Green, Landray, Liu, Matthews, Ong, Pell, Silman, Young, Sprosen, Peakman and Collins2015).
Information on alcohol consumption frequency was obtained through a self-report questionnaire (UK Biobank field ID: 1558; description: Alcohol consumption frequency), which contained seven ‘frequency’ categories, from ‘never’ to ‘daily or almost daily’, individuals also had the option to fill out: ‘prefer not to tell’. Alcohol consumption frequency was assessed in a total of 501 731 subjects. In those who indicated to drink at least once or twice a week (i.e. regular drinkers), also information on alcohol consumption quantity was assessed (N = 348 039). Occasional drinkers, defined as drinking less than once per week (Stockwell et al., Reference Stockwell, Zhao, Panwar, Roemer, Naimi and Chikritzhs2016), were excluded from our analyses, since the nature of our data prevented an accurate assessment of their weekly alcohol consumption quantity. In addition, occasional drinkers are known to underestimate their personal alcohol consumption (Stockwell et al., Reference Stockwell, Zhao and Macdonald2014; Stockwell et al., Reference Stockwell, Zhao, Panwar, Roemer, Naimi and Chikritzhs2016). The assessment of alcohol consumption quantity was made using the average weekly alcohol intake for five general alcohol beverage classes: 1568 (red wine) 1578 (champagne plus white wine), 1598 (spirits), 1558 (beer plus cider), and 1608 (fortified wine). The following item was used ‘In an average WEEK, how many glasses of -class of alcohol- would you drink?’ The total units of alcohol were calculated by multiplying the number of glasses with a factor depending on the class of alcohol, a procedure similar to the one used by Clarke et al. (Reference Clarke, Adams, Davies, Howard, Hall, Padmanabhan, Murray, Smith, Campbell, Hayward, Porteous, Deary and McIntosh2017). The factors we used for the multiplication were 1.67 (red wine and champagne/white wine), 2.3 (beer), 1 (spirit) and 2.25 (fortified wine). Individuals with an alcohol consumption quantity deviating >5 s.d. from their sex-specific mean were excluded from our analyses.
GWAS
SNPs with imputation quality <0.6 and MAF <0.001 were excluded from the association analysis. After genetic QC and exclusion of ethnic outliers, we included 438 308 individuals who were genetically identified to be of white British ancestry in the alcohol consumption frequency GWAS and 307 098 individuals in the alcohol consumption quantity GWAS (i.e. a subset of the alcohol consumption frequency sample). The detailed information regarding the UK biobank genotyping and QC for our analyses was described in Ong et al. (Reference Ong, An, Law, Whiteman, Neale, Gharahkhani and MacGregor2018).
Due to the large number of related individuals in the UK Biobank cohort, the GWAS for alcohol consumption frequency and alcohol consumption quantity were performed using BOLT-LMM, which is a linear mixed model framework that explicitly models the genetic relatedness within the sample (Loh et al., Reference Loh, Tucker, Bulik-Sullivan, Vilhjalmsson, Finucane, Salem, Chasman, Ridker, Neale, Berger, Patterson and Price2015). More specifically, GWAS analyses were performed using the BOLT-LMM v2.3 package, with age, sex, and the top ten ancestral principal components fitted as covariates in the model.
Summary statistics
Data from the other phenotypes used in the current study consisted of summary statistics from previously conducted GWAS meta-analyses, all of which were based on large samples, including UK biobank data (http://www.nealelab.is/blog/2017/7/19/rapid-gwas-of-thousands-of-phenotypes-for-337000-samples-in-the-uk-biobank). We included 41 other phenotypic traits: eight substance use disorder traits, 11 other psychiatric traits, 13 personality and psychological traits, and nine SES-related measures (Table 1). Detailed information about the samples included in this study is provided in Supplementary Table 1.
Table 1. Overview of the GWAS samples used in the current study
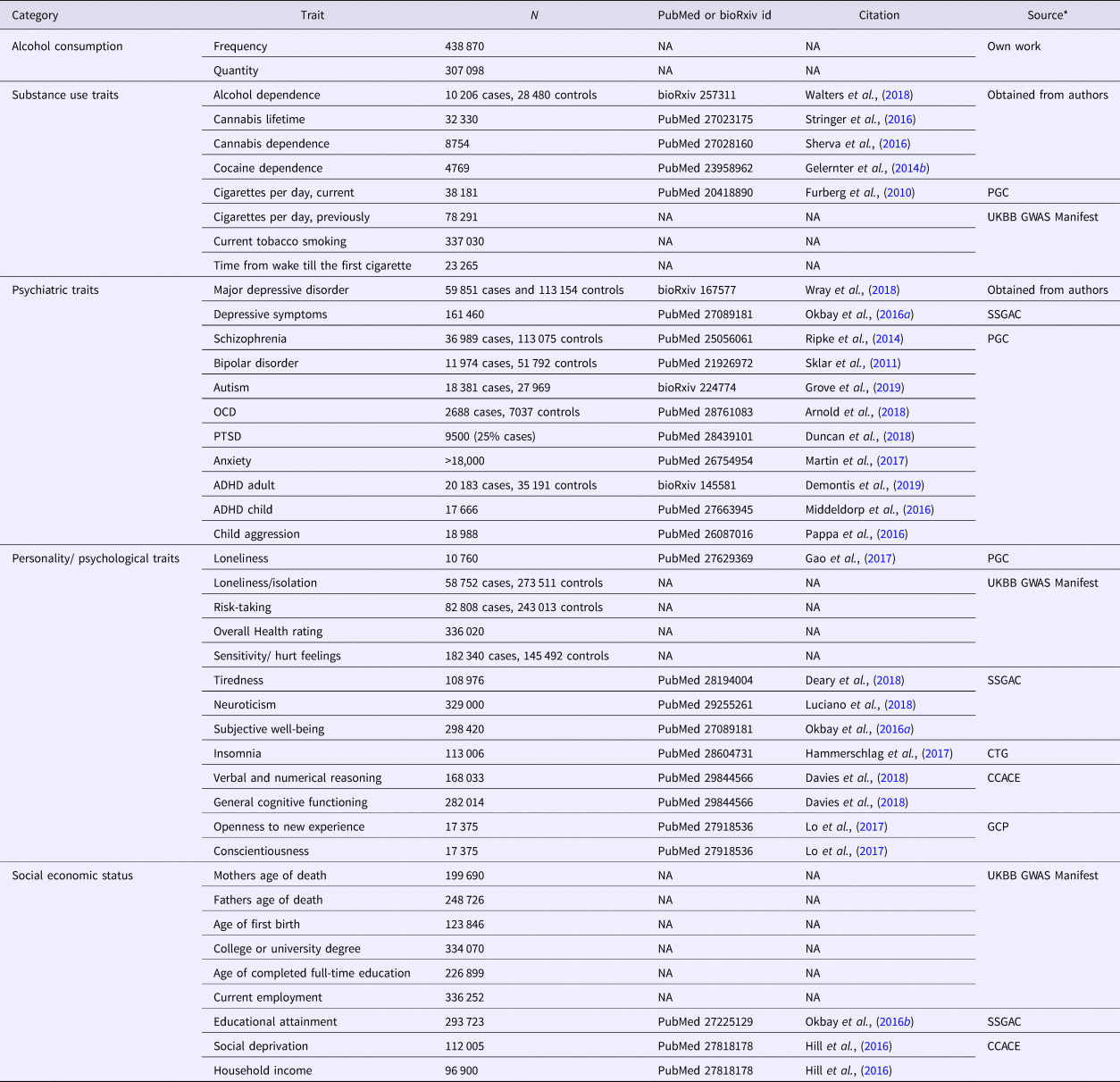
CCACE, Centre for Cognitive Ageing and Cognitive Epidemiology (http://www.ccace.ed.ac.uk/node/335); CTG, Complex Trait Genetics (https://ctg.cncr.nl/software/summary_statistics); GCP, Genetics of Personality Consortium (http://www.tweelingenregister.org/GPC/); PGC, Psychiatric Genetics Consortium (https://www.med.unc.edu/pgc/results-and-downloads); SSGAC, Social Science Genetic Association Consortium (https://www.thessgac.org/data); UKBB GWAS Manifest (https://docs.google.com/spreadsheets/d/1b3oGI2lUt57BcuHttWaZotQcI0-mBRPyZihz87Ms_No/edit#gid=1209628142)
Cross-trait linkage disequilibrium (LD) score regression
LD score regression is a method based on the assumption that an estimated SNP effect-size includes effects of all SNPs in LD (a measure of non-random association between alleles at different loci at the same chromosome in a given population) with that SNP. SNPs that tag (represent due to LD) many other SNPs will have a higher probability of tagging causal genetic variants compared to SNPs that tag few other SNPs. The LD-score is a measure of the amount of genetic variation tagged by a particular SNP within a specific population. Therefore, SNPs with a higher LD-score have, on average, stronger effect sizes than SNPs with lower LD-scores (Bulik-Sullivan et al., Reference Bulik-Sullivan, Loh, Finucane, Ripke, Yang, Patterson, Daly, Price, Neale and Grp2015b). Thus, if the effect size from the association analysis is regressed against the LD-score for each SNP, the slope of the regression line provides an estimate of the proportion of variance explained by all analyzed SNPs (Bulik-Sullivan et al., Reference Bulik-Sullivan, Loh, Finucane, Ripke, Yang, Patterson, Daly, Price, Neale and Grp2015b). An extension of this method, which allows for the estimation of genetic correlation, is cross-trait LD score regression (Bulik-Sullivan et al., Reference Bulik-Sullivan, Finucane, Anttila, Gusev, Day, Loh, Duncan, Perry, Patterson, Robinson, Daly, Price and Neale2015a). The genetic correlation is estimated using the slope from the regression of the product of z-scores from the GWAS on the LD-score. This estimate represents the genetic covariation between the two traits based on all polygenic effects captured by SNPs. Note that genetic correlation is a stronger condition than pleiotropy: a pleiotropic relation only entails that two traits are influenced by the same genetic variant, but for a genetic correlation the directions of effect must be consistently aligned, across the genome, either in the same direction (a positive genetic correlation) or in the opposite direction (a negative genetic correlation) (Bulik-Sullivan et al., Reference Bulik-Sullivan, Finucane, Anttila, Gusev, Day, Loh, Duncan, Perry, Patterson, Robinson, Daly, Price and Neale2015a). The cross-trait LD score regression analyses in the current study were performed using 1 215 002 SNPs that were present in the HapMap 3 reference panel with a 1000 genomes project EUR (European) MAF >5%, with the exclusion of the MHC region.
Significant genetic correlations were identified by applying a FDR 0.05 threshold over all tests. For visualization, we used the R-library ‘ggplot2’.
Results
Alcohol consumption frequency in all drinkers and alcohol consumption quantity in regular drinkers displayed strong polygenic signals (Supplementary Table 1). The genetic correlation between these alcohol consumption metrics was 0.52 (p = 1.31·10−131), while the phenotypic correlation was 0.62 (p < 0.001).
Despite the positive genetic correlation between the two alcohol consumption measures, external phenotypes often showed significant genetic correlations with both alcohol consumption measures in opposite directions. Traits that show significant correlations with both traits, but in opposite directions, are indicated in bold in Fig. 1. Since the GWAS of alcohol consumption quantity was performed in a subset of the sample of the GWAS of alcohol consumption frequency, we investigated the potential influence of this selection. We conducted a sensitivity analysis by performing a GWAS of alcohol consumption frequency in regular drinkers only (the same subset that was used for the alcohol consumption quantity analysis). The opposite pattern of genetic correlations remained largely similar, although for loneliness/isolation we observed a sign flip of the genetic correlation (Supplementary Fig. 2).

Fig. 1. Genetic overlap between alcohol frequency (left panels) and alcohol quantity (right panels) against four categories of traits; SES, substance use disorders, psychiatric disorders, and personality/psychological traits. Traits printed in bold show opposite directions of effect. The error bars represent 95% confidence intervals, an arrow towards one indicates confidence interval >1 or −1, asterisks indicate significant associations (FDR-adjusted p-value <0.05). Social deprivation scores were reversed so that higher social deprivation/Townsend indicates higher SES.
In the full analysis, frequency of alcohol consumption showed significant genetic correlations (FDR-adjusted p-value <0.05) with 31 of the 40 other phenotypic traits: nine SES traits, five substance use disorder traits, eight other psychiatric disorder traits, and nine psychological/personality-related traits. Genetic correlations with measures of SES were consistently positive, i.e. frequent drinking was genetically associated with higher levels of SES. Frequent drinking was genetically negatively associated with tobacco smoking, as indicated by four different smoking measures, i.e. frequent drinking was associated with lower levels of smoking. In contrast, a positive genetic correlation was found between frequency of alcohol consumption and the risk of lifetime cannabis use. We did not observe evidence for a significant genetic correlation between alcohol consumption frequency and AD. The pattern of genetic correlations with other psychiatric disorders was somewhat inconsistent. We observed that frequency of consumption was genetically associated with a reduced risk of ADHD, Major Depressive disorder (MDD), and depressive symptoms, while the genetic correlation with bipolar disorder was positive. A higher genetic predisposition for frequency of alcohol consumption was associated with reduced ratings of insomnia and self-reported tiredness, increased levels of verbal and numerical reasoning, general cognitive functioning, subjective well-being, overall health, loneliness, neuroticism, and an increased vulnerability to getting feelings hurt.
Among regular drinkers, the quantity of alcohol consumption showed significant genetic correlations (FDR-adjusted p-value <0.05) with 27 of the 40 phenotypic traits: eight SES traits, six substance use disorder traits, six psychiatric disorder traits, and seven psychological/ personality-related traits (see Fig. 1). The genetic correlations of alcohol consumption quantity and different measures of SES were consistently negative; high quantity of alcohol consumption was genetically associated with low SES. Alcohol consumption quantity was genetically positively associated with the number of cigarettes smoked per day, AD, and lifetime cannabis use. We observed that a higher genetic risk for increased alcohol consumption quantity was associated with an increased genetic risk for schizophrenia, bipolar disorder, depression, and ADHD. Furthermore, we observed a positive genetic correlation with insomnia, risk-taking, and tiredness ratings, whereas negative genetic correlations were found with subjective well-being, general cognitive function, loneliness isolation, and self-reported health.
Thus, alcohol consumption frequency in all subjects and alcohol consumption quantity in regular drinkers often showed the opposite direction of effect (sign of correlation coefficient) on all four categories of phenotypes (see Fig. 1 and Supplementary Fig. 1), as highlighted by the pattern of association with SES.
Discussion
The overarching aim of this study was to explore the genetic correlation between alcohol consumption frequency and alcohol consumption quantity in regular drinkers, and to examine the genetic correlations of these alcohol consumption measures against 41 other phenotypic traits. We included traits from four broad categories: SES, substance use disorders, other psychiatric disorders, and personality/personality traits, using GWAS summary statistics of studies comprising 4000~ 100 000 subjects. We identified a phenotypic and genetic correlation between alcohol consumption frequency and alcohol consumption quantity of 0.62 (p < 0.001) and 0.52 (p = 1.31·10−131). Although these are substantial correlations, it clearly demonstrates that these two metrics cannot be used interchangeably, confirming previous epidemiological evidence (Berggren and Sutton, Reference Breslow, Chen, Graubard and Mukamal1999; Heckley et al., Reference Heckley, Jarl and Gerdtham2017). Furthermore, alcohol consumption frequency and alcohol consumption quantity showed significant genetic correlations with many of the other examined phenotypic traits, but these genetic correlations often showed effects in opposing directions.
In the categories ‘substance use disorders’, ‘other psychiatric disorders’, and ‘personal/psychological traits’, 14 phenotypic traits showed opposite directions of effect to the alcohol measures central in this study. Alcohol consumption quantity (in regular drinkers) was genetically associated with psychopathology, whereas this was generally not the case for alcohol consumption frequency, suggesting that these alcohol consumption metrics indeed measure different aspects of drinking behavior with different genetic risk profiles. These opposite effects were most consistently found in the genetic correlations with SES-related measures. Epidemiological studies suggest that high SES is related to a higher frequency of alcohol consumption, whereas low SES is related to higher alcohol consumption quantity (Casswell et al., Reference Casswell, Pledger and Hooper2003; Huckle et al., Reference Huckle, You and Casswell2010; Giskes et al., Reference Giskes, Turrell, Bentley and Kavanagh2011). Our results show these opposing phenotypic associations are at least partly explained by opposite associations at the genetic level.
The observation that SES has a positive genetic correlation with alcohol consumption frequency and negative genetic correlation with alcohol consumption quantity may suggest that SES plays a mediating role in the pattern of genetic correlation we observe between these alcohol consumption measures and the other phenotypes. This hypothesis is supported by the observation that individuals with lower SES seem to bear a disproportionate burden of negative alcohol-related consequences (Collins, Reference Collins2016). Previously, genetic correlations have been reported between SES and various behavioral phenotypes (Hill et al., Reference Hill, Hagenaars, Marioni, Harris, Liewald, Davies, Okbay, McIntosh, Gale and Deary2016), including smoking, various psychiatric disorders, and psychology/personality-related traits. Exploration of the patterns observed in the current study with those found by Hill et al., reveals noteworthy similarities. Six of the seven phenotypes that showed significant correlation with SES in the study by Hill et al. (i.e. educational attainment, intelligence, schizophrenia, smoking, MDD, and bipolar disorder) were correlated with alcohol frequency and alcohol quantity in opposite directions in the current study, where high SES and high frequency of alcohol consumption had consistent signs. The only exception was formed by neuroticism, which in contrast to the other results in the comparison, had an opposing direction between high SES and high alcohol consumption frequency. This observation further supports our findings that variability in alcohol consumption frequency and quantity is partly explained through the influence of SES but in opposite directions.
If SES mediates many of the genetic correlations between alcohol consumption traits and other traits, we expect that these traits show opposite effects in their correlation between alcohol consumption frequency and quantity as well. Indeed, the various smoking parameters showed opposite patterns genetic correlations between frequency and quantity of alcohol consumption. Increased alcohol consumption quantity was genetically associated with increased levels of smoking, whereas the increased frequency of alcohol consumption was genetically associated with reduced levels of smoking. The latter seems somewhat counterintuitive, as previous studies have shown a positive phenotypic correlation between smoking and frequency of drinking (Jiang and Ling, Reference Jiang and Ling2013; Lo et al., Reference Lo, Oeltmann, Odhiambo, Beynon, Pevzner, Cain, Laserson and Phillips-Howard2013). Furthermore, those with low SES drink less frequently than those with high SES. Since low SES increases the susceptibility to smoking (Hiscock et al., Reference Hiscock, Bauld, Amos, Fidler and Munafo2012), it seems plausible that the negative genetic association between frequency of alcohol consumption and smoking is mediated by SES. Cannabis lifetime use showed a positive genetic correlation with both alcohol consumption metrics, which seems to be supported by the phenotypic/epidemiological literature (Subbaraman and Kerr, Reference Subbaraman and Kerr2015). Previous research indicates that adolescents from families of higher SES are more likely to experiment with cannabis, which is also suggested by a recent genetic correlation study (Legleye et al., Reference Legleye, Beck, Khlat, Peretti-Watel and Chau2012; Pasman et al., Reference Pasman, Verweij, Gerring, Stringer, Sanchez-Roige, Treur, Abdellaoui, Nivard, Baselmans, Ong, Ip, van der Zee, Bartels, Day, Fontanillas, Elson, de Wit, Davis, MacKillop, Derringer, Branje, Hartman, Heath, van Lier, Madden, Magi, Meeus, Montgomery, Oldehinkel, Pausova, Ramos-Quiroga, Paus, Ribases, Kaprio, Boks, Bell, Spector, Gelernter, Boomsma, Martin, MacGregor, Perry, Palmer, Posthuma, Munafo, Gillespie, Derks and Vink2018), possibly mediating this relation. The positive genetic correlation of alcohol consumption quantity with cannabis lifetime, conflicts with our SES hypothesis, and might be explained through other factors such as risk-taking behavior (de Haan et al., Reference de Haan, Egberts and Heerdink2015; Pasman et al., Reference Pasman, Verweij, Gerring, Stringer, Sanchez-Roige, Treur, Abdellaoui, Nivard, Baselmans, Ong, Ip, van der Zee, Bartels, Day, Fontanillas, Elson, de Wit, Davis, MacKillop, Derringer, Branje, Hartman, Heath, van Lier, Madden, Magi, Meeus, Montgomery, Oldehinkel, Pausova, Ramos-Quiroga, Paus, Ribases, Kaprio, Boks, Bell, Spector, Gelernter, Boomsma, Martin, MacGregor, Perry, Palmer, Posthuma, Munafo, Gillespie, Derks and Vink2018).
With regard to other psychiatric disorders, we found that alcohol consumption frequency and alcohol consumption quantity were correlated with ADHD, depressive symptoms and MDD in opposite directions. Interestingly, the negative genetic association between alcohol consumption frequency and risk of both ADHD and depression is not supported by recent (endo)phenotypic studies, which found no evidence of association of alcohol consumption frequency with ADHD and a positive association with MDD (Weafer et al., Reference Weafer, Milich and Fillmore2011; Edwards et al., Reference Edwards, Heron, Dick, Hickman, Lewis, MacLeod and Kendler2014). However, previous research has indicated a phenotypic association of low SES with the presence of ADHD and MDD (Gavin et al., Reference Gavin, Walton, Chae, Alegria, Jackson and Takeuchi2010; Russell et al., Reference Russell, Ford, Williams and Russell2016). This may imply that genetic variants associated with low SES make individuals more susceptible to develop ADHD and MDD.
The benefit of investigating genetic correlations from GWAS over phenotypic correlations obtained by conventional epidemiological studies are numerous. Genetic correlations provide information on the genetic similarity of phenotypic traits. Therefore, this method allows investigation of whether traits which are in high phenotypic correlation with each other are also genetically similar, e.g., frequency and quantity of alcohol consumption. A high genetic correlation might suggest shared causal genes and shared biological pathways between the traits, while a low genetic correlation would suggest that a high phenotypic correlation is caused by independent biological or environmentally determined mechanisms. This information can provide additional insights into biology underlying co-morbidity and disease risk compared to epidemiological studies. Furthermore, due to the ability of genetic correlation to investigate the genetic similarity between traits, this method can aid in finding alternative phenotypes, which are genetically similar to an outcome of interest, but are less expensive to assess and thus more suited for large scale studies. For example, based on our results it could be concluded that alcohol consumption quantity but not alcohol consumption frequency would be a valid proxy for AD, since AD showed a positive genetic correlation with quantity but not with frequency of alcohol consumption. This finding is in line with results from bivariate twin modeling which has shown a shared heritability of 0.63 (Whitfield et al., Reference Whitfield, Zhu, Madden, Neale, Heath and Martin2004) between AD and quantity of alcohol consumption.
Taken together, the current study suggests a possible mediation of SES in the (genetic) correlation between alcohol consumption measures and other phenotypic traits. However, genetic correlations between SES and substance use disorders, other psychiatric disorders, and personality/psychology-related phenotypes are complex in itself and can be explained in at least three ways. The first explanation is that two phenotypes are genetically correlated due to the influence of genetic variants with pleiotropic effects of the same net direction (e.g. alcohol consumption frequency ß shared genetic effects (G) à educational attainment). The second form of genetic correlation emerges if a certain phenotype is influenced by genetic variants, and that phenotype has a direct causal relationship with another phenotype (e.g. G à educational attainment à alcohol consumption frequency) (Solovieff et al., Reference Solovieff, Cotsapas, Lee, Purcell and Smoller2013). The third form of genetic correlation can occur through gene–environment interaction (e.g. G à specific environment à alcohol consumption frequency and educational attainment). Since our study was designed to only evaluate the correlation structure between these variables, we are unable to definitively untangle different modes of pleiotropy.
We have shown significant genetic correlations between SES and measures of alcohol consumption. SES is a complex construct that is influenced by other traits such as intelligence and personality traits (Hill et al., Reference Hill, Hagenaars, Marioni, Harris, Liewald, Davies, Okbay, McIntosh, Gale and Deary2016). This raises the possibility that the alcohol consumption measures central in the current study are being mediated by phenotypic traits which influence the SES of the individual. Therefore, genetic risk variants which directly influence phenotypic traits which affect SES, might be picked up by a GWAS investigating alcohol consumption. The correct interpretation of such findings is challenging for researchers, as significant loci are normally interpreted within the perspective of plausible biological pathways of the trait under investigation. The current study, and other genetic correlation studies, can help to widen the perspective of researchers outside the scope of biological mechanisms causally affecting a single phenotype of interest. Deciphering the meaning of future GWAS findings in light of the vast interdependency of phenotypes can aid our understanding of the complex genetic architecture of trait and disease and their underlying causal mechanisms.
Our results may have been influenced by selection bias. The response rate in the UK Biobank sample is <5%, and this subset may be an inaccurate representation of the UK population. Previous studies have indicated that the UK Biobank sample is healthier than the general population, suggesting that selection or selection bias may have had some impact on our findings regarding SES, education, and mental health traits (Knudsen et al., Reference Knudsen, Hotopf, Skogen, Overland and Mykletun2010; Fry et al., Reference Fry, Littlejohns, Sudlow, Doherty, Adamska, Sprosen, Collins and Allen2017; Davis et al., Reference Davis, Coleman, Adams, Allen, Breen, Cullen, Dickens, Fox, Graham, Holliday, Howard, John, Lee, McCabe, McIntosh, Pearsall, Smith, Sudlow, Ward, Zammit and Hotopf2018). In addition, the selection of regular drinkers could potentially make our study sensitive to collider bias. Collider bias occurs when two variables independently influence a third variable, and that third variable is conditioned upon. Collider bias can lead to biased estimates of associations (Munafo et al., Reference Munafo, Tilling, Taylor, Evans and Smith2018). Before and after restricting both alcohol consumption measures the general pattern of the opposite genetic correlations remained. The exception to this was loneliness/isolation, where we observed a sign flip of the genetic correlation after restricting alcohol consumption frequency to regular drinkers. However, selecting regular drinkers for the GWAS of alcohol consumption is not likely to have caused collider bias, since regular drinking is not ‘caused’ by frequency of alcohol consumption, namely, regular drinking is a dichotomized version of frequency of alcohol consumption. Likewise, due to its arithmetic relation with frequency of alcohol consumption (see methods), the quantity of alcohol consumption is not fully explained by a causal influence of alcohol frequency. Nevertheless, we cannot fully exclude the possibility of a collider being present, nor would we want to exclude this possibility. Previous alcohol consumption GWAS have used similar selection strategies (e.g. Schumann et al., Reference Schumann, Liu, O'Reilly, Gao, Song, Xu, Ruggeri, Amin, Jia, Preis, Segura Lepe, Akira, Barbieri, Baumeister, Cauchi, Clarke, Enroth, Fischer, Hallfors, Harris, Hieber, Hofer, Hottenga, Johansson, Joshi, Kaartinen, Laitinen, Lemaitre, Loukola, Luan, Lyytikainen, Mangino, Manichaikul, Mbarek, Milaneschi, Moayyeri, Mukamal, Nelson, Nettleton, Partinen, Rawal, Robino, Rose, Sala, Satoh, Schmidt, Schraut, Scott, Smith, Starr, Teumer, Trompet, Uitterlinden, Venturini, Vergnaud, Verweij, Vitart, Vuckovic, Wedenoja, Yengo, Yu, Zhang, Zhao, Boomsma, Chambers, Chasman, Daniela, de Geus, Deary, Eriksson, Esko, Eulenburg, Franco, Froguel, Gieger, Grabe, Gudnason, Gyllensten, Harris, Hartikainen, Heath, Hocking, Hofman, Huth, Jarvelin, Jukema, Kaprio, Kooner, Kutalik, Lahti, Langenberg, Lehtimaki, Liu, Madden, Martin, Morrison, Penninx, Pirastu, Psaty, Raitakari, Ridker, Rose, Rotter, Samani, Schmidt, Spector, Stott, Strachan, Tzoulaki, van der Harst, van Duijn, Marques-Vidal, Vollenweider, Wareham, Whitfield, Wilson, Wolffenbuttel, Bakalkin, Evangelou, Liu, Rice, Desrivieres, Kliewer, Mangelsdorf, Muller, Levy and Elliott2016) and may therefore also be sensitive to selection or collider bias. It should therefore be emphasized that our results are only representative for the population of regular drinkers. Due to the complex pattern of associations between alcohol consumption quantity/frequency with external variables (e.g. SES) observed in our study, it is clear that statements such as ‘one glass of red wine is good for your health’ are a simplification of the true picture.
The findings and conclusions of this study should be interpreted in view of some key limitations. An important limitation is that our estimates of genetic correlations are based on LD score regression using only common SNPs. In family studies, the information on all genetic variants is captured, not just common SNPs. Therefore, family studies estimate the total genetic correlation. If effects are differently correlated among common variants than among rare variants the total genetic correlation can deviate from the common SNP-based genetic correlation (Bulik-Sullivan et al., Reference Bulik-Sullivan, Finucane, Anttila, Gusev, Day, Loh, Duncan, Perry, Patterson, Robinson, Daly, Price and Neale2015a). Furthermore, while alcohol consumption frequency was assessed in a general population sample (regular drinkers, occasional drinkers and teetotalers), alcohol consumption quantity was assessed only in participants who drink regularly. This led to a significantly smaller sample for alcohol consumption quantity, and a reduction in power to find significant genetic correlations relative to frequency of alcohol consumption. Moreover, this study assessed current alcohol intake, which might have led to misclassification of some subjects in regards to the genetic risk factors they carry for alcohol consumption, since alcohol intake is not stable over time (Kerr et al., Reference Kerr, Fillmore and Bostrom2002). Furthermore, despite the comparatively large sample sizes of the alcohol and non-alcohol consumption GWAS in the current study, some of our analyses may still be underpowered to reliably detect genetic correlations. In addition, this study showed merely genetic correlations, it is important to realize that correlations do not equal causation. To fully evaluate causality, Mendelian Randomization (MR) studies ought to be conducted (Lawlor et al., Reference Lawlor, Harbord, Sterne, Timpson and Smith2008), although at present the genetic instruments for most phenotypes investigated in these analyses are not sufficient to allow well-powered MR analyses. The individual genetic correlations should therefore be interpreted carefully with these limitations in mind. Furthermore, the possible mediating role of SES on the alcohol consumption measures might obscure their true genetic correlation to non-SES traits, which has not been tested in the present study.
Conclusion
We have shown that although frequency and quantity of alcohol consumption showed substantial genetic overlap, they consistently show opposite patterns of genetic correlations with SES-related phenotypes. Our findings provide novel insights into the genetic architecture shared between alcohol consumption and phenotypes of four broad categories (SES, substance use disorders, other psychiatric disorders and psychology/personality traits) and hints SES as a potential mediator of the relationship. The latter indicates that future studies should carefully consider the potential influence of SES on the shared genetic etiology between alcohol and adverse conditions.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0033291719000357
Author ORCIDs
Andries T. Marees, 0000-0002-9781-7492; Dirk J. A. Smit, 0000-0002-9639-0992; Jue-Sheng Ong, 0000-0002-6062-710X; Stuart MacGregor, 0000-0001-6731-8142; Jiyuan An, 0000-0002-8183-8248; Damiaan Denys, 0000-0002-3191-3844; Florence Vorspan, 0000-0001-5168-3727; Wim van den Brink, 0000-0001-8301-0121; Eske M. Derks, 0000-0002-6292-6883
Acknowledgements
This work was conducted using the UK Biobank Resource (application number 25331). We thank Scott Wood and John Pearson from QIMR Berghofer for IT support. The Substance Use Disorders Working Group of the Psychiatric Genomics Consortium (PGC-SUD) is supported by funds from NIDA and NIMH to MH109532 and, previously, with analyst support from NIAAA to U01AA008401 (COGA). We gratefully acknowledge our contributing studies and the participants in those studies without whom this effort would not be possible.
This work is supported by a project grant (1123248) from the Australian National Health and Medical Research Council (NHMRC). ATM and EMD are supported by the Foundation Volksbond Rotterdam, FV is supported by the Investissement d'Avenir program managed by the ANR under reference ANR-11-IDEX-0004-02. JSO received scholarship support from the University of Queensland and QIMR Berghofer Medical Research Institute
Conflict of interest
Dr Vorspan received over the last 5 years the following support/funding: Congress registration and travel: Lundbeck Pharmaceuticals (2015). European or French Ministry of Research competitive research grants: Labex BioPsy (ANR, Grant/Award Number: ANR-11-IDEX-0004-02; Investissement d'Avenir programme): 2013, 2014, 2016; ANR Samenta 2013; ERA-net Neuron 2013, 2017. Fondation pour la Recherche en Alcoologie (FRA), hébergée par la fondation de France: 2016, 2018. The other authors declare no conflict of interest with regard to this study.





 Open access
Open access