



1. Introduction
The discrete equation

is a popular modeling framework for analysing the dynamical behaviour of a single species. In (1.1), $x_n\geq 0$ is the population density in the $n$
is the population density in the $n$ -th generation and $g(x_n)\geq 0$
-th generation and $g(x_n)\geq 0$ represents the density-dependent growth rate (or fitness function) from generation to generation. A common assumption in population dynamics is that $g$
represents the density-dependent growth rate (or fitness function) from generation to generation. A common assumption in population dynamics is that $g$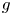 is decreasing. This means that the growth rate is mainly determined by negative density-dependent mechanisms such as intra-specific competition [Reference Fishman5] or cannibalism. However, cooperative predation, resource defense, increased availability of mates and conspecific enhancement of reproduction are other biological mechanisms producing non-monotone growth rates, see [Reference Kang and Yakubu20]. For example, alders, big leaf maples, poplars and some pine trees thrive at low densities and decrease at high ones due to overcrowding effects and other ecosystem constraints [Reference Franke and Yakubu6, Reference Franke and Yakubu8, Reference Franke and Yakubu10, Reference Selgrade and Namkoong33].
is decreasing. This means that the growth rate is mainly determined by negative density-dependent mechanisms such as intra-specific competition [Reference Fishman5] or cannibalism. However, cooperative predation, resource defense, increased availability of mates and conspecific enhancement of reproduction are other biological mechanisms producing non-monotone growth rates, see [Reference Kang and Yakubu20]. For example, alders, big leaf maples, poplars and some pine trees thrive at low densities and decrease at high ones due to overcrowding effects and other ecosystem constraints [Reference Franke and Yakubu6, Reference Franke and Yakubu8, Reference Franke and Yakubu10, Reference Selgrade and Namkoong33].
In describing the interactions of $k$ species, a natural extension of equation (1.1) is
species, a natural extension of equation (1.1) is

The growth rate $H_i$ is typically of the form
is typically of the form

with $g_i:[0,\,+\infty )\longrightarrow [0,\,+\infty )$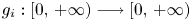 a continuous function. In particular, if $a_{ij}>0$
a continuous function. In particular, if $a_{ij}>0$ and $g_{i}$
and $g_{i}$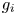 is decreasing for all $i,\,j=1,\,\ldots,\,n$
is decreasing for all $i,\,j=1,\,\ldots,\,n$ , system (1.2) describes the evolution of $k$
, system (1.2) describes the evolution of $k$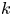 competing species. See [Reference Franke and Yakubu7, Reference Franke and Yakubu9, Reference Franke and Yakubu10, Reference Gyllenberg, Jiang, Niu and Yan13, Reference May23] and the references therein for a detailed discussion on these models.
competing species. See [Reference Franke and Yakubu7, Reference Franke and Yakubu9, Reference Franke and Yakubu10, Reference Gyllenberg, Jiang, Niu and Yan13, Reference May23] and the references therein for a detailed discussion on these models.
Understading the dynamical behaviour of (1.2) is of critical importance from an applied point of view. There are several approaches for analysing this issue for competitive systems. For example, the convexity arguments given by Kon [Reference Kon22], the split Liapunov function method given by Baigent and Hou [Reference Baigent and Hou2, Reference Hou and Baigent18] or the theory of carrying simplex [Reference Ruiz-Herrera31]. Recently, Hou [Reference Hou16, Reference Hou17] has provided a criterion of global attraction based on the relative position of the nullclines reminiscent to the classical results for the Lotka–Volterra system

see [Reference Ahmad and Lazer1] and the references therein. In contrast with the monotone case, the literature on non-monotone systems is relatively scarce. It is worth noting that system (1.2) can exhibit chaotic dynamics.
In this paper we describe the global dynamical picture of model (1.2) when the functions $g_{i}$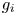 are not necessarily decreasing. In particular, our criteria could be perceived as an extension of Hou's results to non-monotone systems. The method of proof is completely different from those papers mentioned above. First, we divide the phase space in suitable positively invariant regions and then we apply the theory of translation arcs [Reference Campos, Ortega and Tineo3, Reference Graff and Ruiz-Herrera12, Reference Ortega27] in a subtle manner. As emphasized in § 3, our conclusions are rather sharp.
are not necessarily decreasing. In particular, our criteria could be perceived as an extension of Hou's results to non-monotone systems. The method of proof is completely different from those papers mentioned above. First, we divide the phase space in suitable positively invariant regions and then we apply the theory of translation arcs [Reference Campos, Ortega and Tineo3, Reference Graff and Ruiz-Herrera12, Reference Ortega27] in a subtle manner. As emphasized in § 3, our conclusions are rather sharp.
The organization of the paper is as follows. In § 2, we introduce some notation and definitions. In § 3, we give the main results of the paper. Applications to population models for pioneer-climax species [Reference Franke and Yakubu7, Reference Franke and Yakubu9, Reference Franke and Yakubu10] and species with weak Allee effects [Reference Kang and Yakubu20] are discussed. To show the versatility of our results with different interactions, we discuss the dynamical behaviour of several classical predator–prey systems. We conclude the paper with a discussion on our findings.
2. Mathematical framework
The Euclidean disk with centre at $p=(p_{1},\,p_{2})\in \mathbb {R}^{2}$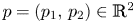 and radius $R>0$
and radius $R>0$ is denoted by
is denoted by

with $\|(x,\,y)\|=\sqrt {x^{2}+y^{2}}$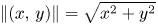 . A subset of the plane homeomorphic to a (closed) Euclidean disk is called a topological disk. A map $h:V\subset \mathbb {R}^{2}\longrightarrow V$
. A subset of the plane homeomorphic to a (closed) Euclidean disk is called a topological disk. A map $h:V\subset \mathbb {R}^{2}\longrightarrow V$ which is continuous and injective is called an embedding. Notice that if $h(V)=V$
which is continuous and injective is called an embedding. Notice that if $h(V)=V$ , then $h$
, then $h$ is a homeomorphism. In this section, we work, without further mention, with embeddings defined on topological disks. We recall that $h:D\longrightarrow D$
is a homeomorphism. In this section, we work, without further mention, with embeddings defined on topological disks. We recall that $h:D\longrightarrow D$ is an orientation preserving embedding if it has degree one, that is,
is an orientation preserving embedding if it has degree one, that is,

where $h(p_{0})=q_{0}$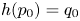 with $p_{0}\in Int D$
with $p_{0}\in Int D$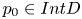 and $U\subset D$
and $U\subset D$ is any open neighbourhood of $p_{0}$
is any open neighbourhood of $p_{0}$ . The reader can consult the appendix in [Reference Ortega27] for a detailed discussion on topological degree and index. When $h: D\longrightarrow D$
. The reader can consult the appendix in [Reference Ortega27] for a detailed discussion on topological degree and index. When $h: D\longrightarrow D$ is an embedding of class $\mathcal {C}^{1}$
is an embedding of class $\mathcal {C}^{1}$ , the sign of the determinant of the Jacobian matrix of $h$
, the sign of the determinant of the Jacobian matrix of $h$ can determine whether $h$
can determine whether $h$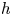 preserves the orientation. Specifically, if
preserves the orientation. Specifically, if

for all $x\in D$ , then $h$
, then $h$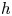 is an orientation preserving embedding. Now we give a practical criterion to deduce when a map is an orientation preserving embedding. We have employed the notation $\partial D$
is an orientation preserving embedding. Now we give a practical criterion to deduce when a map is an orientation preserving embedding. We have employed the notation $\partial D$ to refer to the boundary of $D$
to refer to the boundary of $D$ in $\mathbb {R}^{2}$
in $\mathbb {R}^{2}$ .
.
Proposition 2.1 Let $h:D\longrightarrow D$ be a map of class $\mathcal {C}^{1}$
be a map of class $\mathcal {C}^{1}$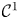 with $D\subset \mathbb {R}^{2}$
with $D\subset \mathbb {R}^{2}$ a topological disk. Assume the following conditions:
a topological disk. Assume the following conditions:
(i) $h(\partial D)\subset D$
 .
.(ii) $\det Dh(x)>0$
for all $x\in D$.(iii) There is a point $q\in D$
with a unique pre-image, that is, $h^{-1}(\{q\})=\{p\}$.
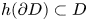

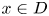
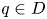
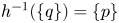
Then, $h(D)\subset D$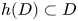 and
and

is an orientation preserving embedding.
Proof. Using that $h^{-1}(\{q\})=\{p\}$ and that $h$
and that $h$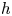 is locally injective (by (ii)), we deduce that
is locally injective (by (ii)), we deduce that

is a homeomorphism (see lemma 2.3.4 in [Reference Chow and Hale4]). We know that $h(D)$ is a topological disk with boundary contained in the topological disk $D$
is a topological disk with boundary contained in the topological disk $D$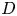 . This implies that $h(D)\subset D$
. This implies that $h(D)\subset D$ (see lemma 6 in [Reference Ortega27, p. 39]). Hence,
(see lemma 6 in [Reference Ortega27, p. 39]). Hence,

is an embedding. Notice that $h$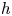 is an orientation preserving embedding because
is an orientation preserving embedding because

for all $x\in D$ .
.
Given $h:D\longrightarrow D$ an embedding, the $\omega$
an embedding, the $\omega$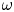 -limit set of a point $p\in D$
-limit set of a point $p\in D$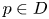 is defined as
is defined as

We say that $h$ has trivial dynamics if for all $x\in D$
has trivial dynamics if for all $x\in D$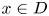 ,
,

where $Fix(h)$ denotes the fixed point set of $h$
denotes the fixed point set of $h$ . It is worth noting that if $h$
. It is worth noting that if $h$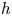 is dissipative and $\omega (h,\,x)\subset Fix(h)$
is dissipative and $\omega (h,\,x)\subset Fix(h)$ for all $x\in D$
for all $x\in D$ , then $\omega (h,\,x)$
, then $\omega (h,\,x)$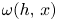 is a connected set (see proposition 2 in [Reference Ortega27, p. 42]). In particular, if $h$
is a connected set (see proposition 2 in [Reference Ortega27, p. 42]). In particular, if $h$ has a finite number of fixed points, then for all $x\in D$
has a finite number of fixed points, then for all $x\in D$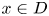 , there is $q\in Fix(h)$
, there is $q\in Fix(h)$ so that $\omega (h,\,x)=\{q\}$
so that $\omega (h,\,x)=\{q\}$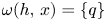 . We stress that an embedding $h:D\longrightarrow D$
. We stress that an embedding $h:D\longrightarrow D$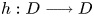 defined on a topological disk is always dissipative.
defined on a topological disk is always dissipative.
Next we recall two results based on the theory of translation arcs.
Theorem 2.2 (Corollary 2.1 in [Reference Ruiz-Herrera30], [Reference Campos, Ortega and Tineo3])
Let $h: D\longrightarrow D$ be an orientation preserving embedding defined on the topological disk $D$
be an orientation preserving embedding defined on the topological disk $D$ . If $Fix(h)\subset \partial D,$
. If $Fix(h)\subset \partial D,$ then $h$
then $h$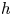 has trivial dynamics.
has trivial dynamics.
Theorem 2.3 (Theorem 2.1 in [Reference Niu and Ruiz-Herrera26], [Reference Ortega27])
Let $h: D\longrightarrow D$ be an orientation preserving embedding defined on the topological disk $D$
be an orientation preserving embedding defined on the topological disk $D$ . If $Fix(h)\cap Int D=\{q\}$
. If $Fix(h)\cap Int D=\{q\}$ with $index(h,\,q)=-1$
with $index(h,\,q)=-1$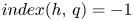 then $h$
then $h$ has trivial dynamics.
has trivial dynamics.
In the previous theorem, $index(h,\,q)$ denotes the usual topological index of $h$
denotes the usual topological index of $h$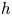 at $q$
at $q$ . We mention that theorem 2.1 in [Reference Niu and Ruiz-Herrera26] deals with homeomorphisms. However, the same proof works for embeddings.
. We mention that theorem 2.1 in [Reference Niu and Ruiz-Herrera26] deals with homeomorphisms. However, the same proof works for embeddings.
3. Geometric criteria of trivial dynamics
In this section we consider the system

with $\alpha,\,\beta >0$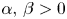 and $g_{i}:[0,\,+\infty )\longrightarrow (0,\,+\infty )$
and $g_{i}:[0,\,+\infty )\longrightarrow (0,\,+\infty )$ a function of class $\mathcal {C}^{1}$
a function of class $\mathcal {C}^{1}$ for $i=1,\,2$
for $i=1,\,2$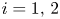 . Denote by
. Denote by

Notice that $G([0,\,+\infty )^{2})\subset [0,\,+\infty )^{2}$ . A common assumption for the single-species model (1.1), already stated by Ricker [Reference Ricker28] and by Moran [Reference Moran25], is that there is a positive equilibrium $p$
. A common assumption for the single-species model (1.1), already stated by Ricker [Reference Ricker28] and by Moran [Reference Moran25], is that there is a positive equilibrium $p$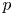 (the carrying capacity) so that
(the carrying capacity) so that
(C) $g(x)>1$
for all $x\in (0,\,p)$ and $g(x)<1$ for all $x>p$.
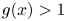

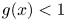

The previous condition can be adapted in model (3.1) as follows:
(P) For $i=1,\,2$
, there exists $r_{i}>0$ so that $g_{i}(x)>1$ if $x\in (0,\,r_{i})$ and $g_{i}(x)<1$ if $x>r_{i}$.
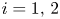



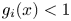

Biologically, condition (P) means that the density of population of species $i$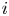 increases (resp. decreases) in the next generation when the weighted total biomass of both species is below (resp. above) a threshold.
increases (resp. decreases) in the next generation when the weighted total biomass of both species is below (resp. above) a threshold.
In this section, we will make the assumption
(S) $\det DG(x,\,y)>0$
for all $(x,\,y)\in \mathcal {R}$, where $\mathcal {R}=[0,\, A_{1}]\times [0,\,A_{2}]$ with
\[ A_{1}=\max\left\{r_{1},\frac{r_{2}}{\beta}\right\},\quad A_{2}=\max\left\{r_{2},\frac{r_{1}}{\alpha}\right\}. \]
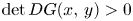
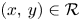
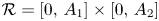

Condition (S) implies that $G$ is one-to-one on $\mathcal {R}$
is one-to-one on $\mathcal {R}$ . The biological meaning of this is simple. If we take two different initial data, the densities of population are different each other in any future generation. To avoid technical problems, we also suppose:
. The biological meaning of this is simple. If we take two different initial data, the densities of population are different each other in any future generation. To avoid technical problems, we also suppose:
(H) $g_{i}'(r_{i})\not =0$
for $i=1,\,2$.


Define

and

The system has, at most, four equilibria namely $(0,\,0)$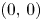 , $(r_{1},\,0)$
, $(r_{1},\,0)$ , $(0,\,r_{2})$
, $(0,\,r_{2})$ and
and

Obviously, the last equilibrium is located at the intersection of $L_{1}$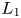 and $L_{2}$
and $L_{2}$ . In our analysis, we exclude the case $L_{1}=L_{2}$
. In our analysis, we exclude the case $L_{1}=L_{2}$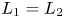 ($\alpha =\beta =1$
($\alpha =\beta =1$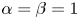 and $r_{1}=r_{2}$
and $r_{1}=r_{2}$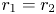 ).
).
We prove that the relative position of $L_{1}$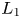 and $L_{2}$
and $L_{2}$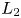 completely determines the dynamical behaviour of (3.1). This is a well-known result when the system is monotone. Our contribution will be to show that it is also true for non-monotone systems. We stress that if $(x,\,y)\in (0,\,+\infty )^{2}\backslash \mathcal {R}$
completely determines the dynamical behaviour of (3.1). This is a well-known result when the system is monotone. Our contribution will be to show that it is also true for non-monotone systems. We stress that if $(x,\,y)\in (0,\,+\infty )^{2}\backslash \mathcal {R}$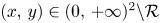 , then
, then

Proposition 3.1 Assume that (P) and (S) are satisfied. Then, $G(\mathcal {R})\subset \mathcal {R}$ and
and

is an orientation preserving embedding.
Proof. First we notice that $G^{-1}(\{0\})=\{0\}$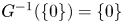 . Next we prove that $G(\partial \mathcal {R})\subset \mathcal {R}$
. Next we prove that $G(\partial \mathcal {R})\subset \mathcal {R}$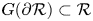 . Take a point of the form $(A_{1},\,y)$
. Take a point of the form $(A_{1},\,y)$ with $y\in [0,\, A_{2}]$
with $y\in [0,\, A_{2}]$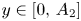 . We observe that $A_{1}+\alpha y\geq r_{1}$
. We observe that $A_{1}+\alpha y\geq r_{1}$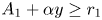 and $y+\beta A_{1}\geq r_{2}$
and $y+\beta A_{1}\geq r_{2}$ . Then, by condition (P),
. Then, by condition (P),

Now, it is clear that

Analogously, we can prove that

We also observe that condition (S) ensures that $G_{1}(x,\,0)=xg_{1}(x)$ and $G_{2}(0,\,y)=yg_{2}(y)$
and $G_{2}(0,\,y)=yg_{2}(y)$ are strictly increasing (they are locally injective and have two fixed points). Thus, $G([0,\,A_{1}]\times \{0\})\subset [0,\,A_{1}]\times \{0\}$
are strictly increasing (they are locally injective and have two fixed points). Thus, $G([0,\,A_{1}]\times \{0\})\subset [0,\,A_{1}]\times \{0\}$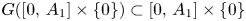 and $G(\{0\}\times [0,\,A_{2}])\subset \{0\}\times [0,\,A_{2}]$
and $G(\{0\}\times [0,\,A_{2}])\subset \{0\}\times [0,\,A_{2}]$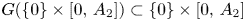 . Collecting the above information, we deduce that
. Collecting the above information, we deduce that

The conclusion now follows from proposition 2.1 immediately.
Lemma 3.2 Let $\{(x_{n},\,y_{n})\}$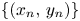 be a sequence of (3.1) with initial condition $(x_{0},\,y_{0})\not \in \mathcal {R}$
be a sequence of (3.1) with initial condition $(x_{0},\,y_{0})\not \in \mathcal {R}$ . Assume that (P) and (S) hold. Then, one of the following properties is satisfied:
. Assume that (P) and (S) hold. Then, one of the following properties is satisfied:
(i) $\{(x_{n},\,y_{n})\}\longrightarrow q$
with $q$ an equilibrium of (3.1).(ii) There exists $n_{0}\in \mathbb {N}$
so that $(x_{n},\,y_{n})\in \mathcal {R}$ for all $n\geq n_{0}$.
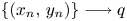
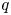


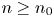
Proof. Assume that $(x_{0},\,y_{0})\in (0,\,+\infty )^{2}$ . If $(x_{n},\,y_{n})\not \in \mathcal {R}$
. If $(x_{n},\,y_{n})\not \in \mathcal {R}$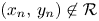 for all $n\in \mathbb {N}$
for all $n\in \mathbb {N}$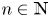 , condition (P) ensures that $\{x_{n}\}$
, condition (P) ensures that $\{x_{n}\}$ and $\{y_{n}\}$
and $\{y_{n}\}$ are strictly decreasing, see (3.3). Then, (i) holds. If there is $n_{0}\in \mathbb {N}$
are strictly decreasing, see (3.3). Then, (i) holds. If there is $n_{0}\in \mathbb {N}$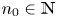 so that $(x_{n_{0}},\,y_{n_{0}})\in \mathcal {R}$
so that $(x_{n_{0}},\,y_{n_{0}})\in \mathcal {R}$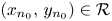 , we deduce that $(x_{n},\,y_{n})\in \mathcal {R}$
, we deduce that $(x_{n},\,y_{n})\in \mathcal {R}$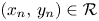 for all $n\geq n_{0}$
for all $n\geq n_{0}$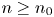 . Notice that $G(\mathcal {R})\subset \mathcal {R}$
. Notice that $G(\mathcal {R})\subset \mathcal {R}$ by the previous proposition. The case $x_{0}=0$
by the previous proposition. The case $x_{0}=0$ (resp. $y_{0}=0$
(resp. $y_{0}=0$ ) is treated in an analogous manner. Observe that in such a case $x_{n}=0$
) is treated in an analogous manner. Observe that in such a case $x_{n}=0$ (resp. $y_{n}=0$
(resp. $y_{n}=0$ ) for all $n\in \mathbb {N}$
) for all $n\in \mathbb {N}$ and $y_{n}$
and $y_{n}$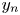 (resp. $x_{n}$
(resp. $x_{n}$ ) is decreasing provided $y_{n}>A_{2}$
) is decreasing provided $y_{n}>A_{2}$ (resp. $x_{n}>A_{1}$
(resp. $x_{n}>A_{1}$ ).
).
Remark 3.3 Lemma 3.2 says that it is enough for analysing the dynamical behaviour of (3.1) in $\mathcal {R}$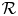 . We repeatedly use this fact in the subsequent results.
. We repeatedly use this fact in the subsequent results.
The following result describes the behaviour of (3.1) on the axes.
Lemma 3.4 Assume that (P) and (S) hold. Then, $r_{1}$ and $r_{2}$
and $r_{2}$ are globally asymptotically stable for
are globally asymptotically stable for

and

respectively in $(0,\,+\infty )$ .
.
Proof. We study the equilibrium $r_{1}$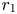 . The other case is analogous and we omit the details. By remark 3.3, we can restrict our attention on $[0,\,A_{1}]$
. The other case is analogous and we omit the details. By remark 3.3, we can restrict our attention on $[0,\,A_{1}]$ . As mentioned in the proof of proposition 3.1, $\varphi (x)=xg_{1}(x)$
. As mentioned in the proof of proposition 3.1, $\varphi (x)=xg_{1}(x)$ is strictly increasing in $[0,\,A_{1}]$
is strictly increasing in $[0,\,A_{1}]$ . We remark that the unique fixed points of $\varphi$
. We remark that the unique fixed points of $\varphi$ are $0$
are $0$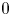 and $r_{1}$
and $r_{1}$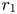 , and the fact $\varphi (A_{1})\leq A_{1}$
, and the fact $\varphi (A_{1})\leq A_{1}$ . By condition (P), $g_1(x)>1$
. By condition (P), $g_1(x)>1$ for all $x\in (0,\,r_{1})$
for all $x\in (0,\,r_{1})$ . Hence, $\varphi (x)>x$
. Hence, $\varphi (x)>x$ for all $x\in (0,\,r_{1})$
for all $x\in (0,\,r_{1})$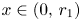 . Now it is clear that $0$
. Now it is clear that $0$ is unstable. Thus, $r_{1}$
is unstable. Thus, $r_{1}$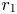 is a global attractor for $\varphi$
is a global attractor for $\varphi$ in $(0,\,A_{1}]$
in $(0,\,A_{1}]$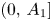 .
.
Now we are in a position to give the main result of this section.
Theorem 3.5 Assume (P), (H) and (S) hold. Then the dynamical behaviour of (3.1) is as follows:
(i) $(r_{1},\,0)$
is a global attractor in $(0,\,+\infty )^{2}$ provided $\frac {r_{2}}{\beta }\leq r_{1}$ and $\frac {r_{1}}{\alpha }>r_{2}$.(ii) $(0,\,r_{2})$
is a global attractor in $(0,\,+\infty )^{2}$ provided $\frac {r_{2}}{\beta }> r_{1}$ and $\frac {r_{1}}{\alpha }\leq r_{2}$.(iii) $(x^{*},\,y^{*})$
is a global attractor in $(0,\,+\infty )^{2}$ provided $\frac {r_{2}}{\beta }>r_{1}$ and $\frac {r_{1}}{\alpha }>r_{2}$.(iv) There is trivial dynamics in (3.1) with $(r_{1},\,0)$
and $(0,\,r_{2})$ as local attractors in $(0,\,+\infty )^{2}$ provided $\frac {r_{2}}{\beta }< r_{1}$ and $\frac {r_{1}}{\alpha }< r_{2}$.


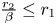



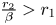
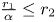

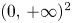




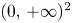


Proof. First we realize that the origin is always a local repeller in $(0,\,+\infty )^{2}$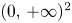 . Indeed, by condition (P), there is a neighbourhood $U$
. Indeed, by condition (P), there is a neighbourhood $U$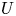 of the origin in $\mathcal {R}$
of the origin in $\mathcal {R}$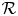 so that $g_{1}(x+\alpha y)>1$
so that $g_{1}(x+\alpha y)>1$ and $g_{2}(\beta x+y)>1$
and $g_{2}(\beta x+y)>1$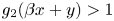 for all $(x,\,y)\in U$
for all $(x,\,y)\in U$ with $x\not =0$
with $x\not =0$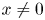 and $y\not =0$
and $y\not =0$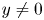 . This implies that $G_{1}(x,\,y)>x$
. This implies that $G_{1}(x,\,y)>x$ and $G_{2}(x,\,y)>y$
and $G_{2}(x,\,y)>y$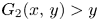 for all $(x,\,y)\in U$
for all $(x,\,y)\in U$ with $x\not =0$
with $x\not =0$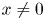 and $y\not =0$
and $y\not =0$ . Since $G((0,\,+\infty )^{2})\subset (0,\,+\infty )^{2}$
. Since $G((0,\,+\infty )^{2})\subset (0,\,+\infty )^{2}$ , it is clear that the origin is a local repeller in $(0,\,+\infty )^{2}$
, it is clear that the origin is a local repeller in $(0,\,+\infty )^{2}$ . We also stress that by remark 3.3, it is enough to study the dynamical behaviour in $\mathcal {R}$
. We also stress that by remark 3.3, it is enough to study the dynamical behaviour in $\mathcal {R}$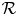 . Now we are ready to prove the theorem.
. Now we are ready to prove the theorem.
(i) In this case, $L_{1}$ and $L_{2}$
and $L_{2}$ do not intersect in $(0,\,+\infty )^{2}$
do not intersect in $(0,\,+\infty )^{2}$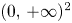 . Then, $G|_{\mathcal {R}}$
. Then, $G|_{\mathcal {R}}$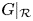 has three fixed points, namely, $(0,\,0)$
has three fixed points, namely, $(0,\,0)$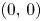 , $(r_{1},\,0)$
, $(r_{1},\,0)$ and $(0,\,r_{2})$
and $(0,\,r_{2})$ . Since $Fix(G|_{\mathcal {R}})\subset \partial \mathcal {R}$
. Since $Fix(G|_{\mathcal {R}})\subset \partial \mathcal {R}$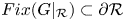 , theorem 2.2 and proposition 3.1 imply that for each $(x_{0},\,y_{0})\in Int\mathcal {R}$
, theorem 2.2 and proposition 3.1 imply that for each $(x_{0},\,y_{0})\in Int\mathcal {R}$ , there exists $q\in Fix(G|_{\mathcal {R}})$
, there exists $q\in Fix(G|_{\mathcal {R}})$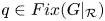 so that
so that

Next we prove that $q\not =(0,\,0)$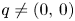 and $q\not =(0,\,r_{2})$
and $q\not =(0,\,r_{2})$ . To see this, we check that both fixed points are local repellers in $(0,\,+\infty )^{2}$
. To see this, we check that both fixed points are local repellers in $(0,\,+\infty )^{2}$ . At the beginning, we have already mentioned this fact for the origin. On the other hand, the eigenvalues of the linearized system at $(0,\,r_{2})$
. At the beginning, we have already mentioned this fact for the origin. On the other hand, the eigenvalues of the linearized system at $(0,\,r_{2})$ are
are

where the associated eigenvectors are $(0,\,1)$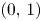 and $(w_{1},\,w_{2})$
and $(w_{1},\,w_{2})$ with $w_{1}\not =0$
with $w_{1}\not =0$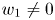 respectively. Since $r_{1}>\alpha r_{2}$
respectively. Since $r_{1}>\alpha r_{2}$ and (P), we conclude that $g_{1}(\alpha r_{2})>1$
and (P), we conclude that $g_{1}(\alpha r_{2})>1$ . On the other hand, we have already seen in the proof of proposition 3.1 that $\phi (y)=yg_{2}(y)$
. On the other hand, we have already seen in the proof of proposition 3.1 that $\phi (y)=yg_{2}(y)$ is an increasing function. Moreover, $r_{2}>0$
is an increasing function. Moreover, $r_{2}>0$ is a global attractor of $\phi$
is a global attractor of $\phi$ by lemma 3.4. Using these two facts together with (H), we conclude that $\phi '(r_{2})=1+r_{2}g_{2}'(r_{2})\in [0,\,1)$
by lemma 3.4. Using these two facts together with (H), we conclude that $\phi '(r_{2})=1+r_{2}g_{2}'(r_{2})\in [0,\,1)$ . Observe that by (S), $\phi '(r_{2})\not =0$
. Observe that by (S), $\phi '(r_{2})\not =0$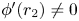 . Now, it is clear that $(0,\,r_{2})$
. Now, it is clear that $(0,\,r_{2})$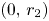 is a hyperbolic saddle point. In particular, it is a local repeller in $(0,\,+\infty )^{2}$
is a hyperbolic saddle point. In particular, it is a local repeller in $(0,\,+\infty )^{2}$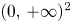 .
.
The proof of (ii) is analogous and we omit the details.
(iii) In this case, $G|_{\mathcal {R}}$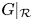 has four fixed points, namely, $(0,\,0)$
has four fixed points, namely, $(0,\,0)$ , $(r_{1},\,0)$
, $(r_{1},\,0)$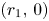 , $(0,\,r_{2})$
, $(0,\,r_{2})$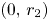 and $(x^{*},\,y^{*})$
and $(x^{*},\,y^{*})$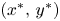 . Define $\mathcal {A}=\mathcal {L}_{1}^{-}\cup \mathcal {L}_{2}^{-}$
. Define $\mathcal {A}=\mathcal {L}_{1}^{-}\cup \mathcal {L}_{2}^{-}$ . The boundary of $\mathcal {A}$
. The boundary of $\mathcal {A}$ is made of four segments: $\{0\}\times [0,\,\frac {r_{1}}{\alpha }]$
is made of four segments: $\{0\}\times [0,\,\frac {r_{1}}{\alpha }]$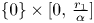 , $[0,\,\frac {r_{2}}{\beta }]\times \{0\}$
, $[0,\,\frac {r_{2}}{\beta }]\times \{0\}$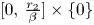 , $l_{1}$
, $l_{1}$ and $l_{2}$
and $l_{2}$ where $l_{1}$
where $l_{1}$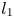 is the segment on $L_{1}$
is the segment on $L_{1}$ with ends at $(0,\,\frac {r_{1}}{\alpha })$
with ends at $(0,\,\frac {r_{1}}{\alpha })$ and $(x^{*},\,y^{*})$
and $(x^{*},\,y^{*})$ and $l_{2}$
and $l_{2}$ is the segment on $L_{2}$
is the segment on $L_{2}$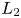 with ends at $(\frac {r_{2}}{\beta },\,0)$
with ends at $(\frac {r_{2}}{\beta },\,0)$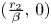 and $(x^{*},\,y^{*})$
and $(x^{*},\,y^{*})$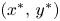 . Since $\frac {r_{2}}{\beta }>r_{1}$
. Since $\frac {r_{2}}{\beta }>r_{1}$ and $\frac {r_{1}}{\alpha }>r_{2}$
and $\frac {r_{1}}{\alpha }>r_{2}$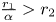 , we have that
, we have that

and

Then, given $(x,\,y)\in l_{1}$ different from the ends $(x^{*},\,y^{*})$
different from the ends $(x^{*},\,y^{*})$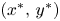 and $(0,\,\frac {r_{1}}{\alpha })$
and $(0,\,\frac {r_{1}}{\alpha })$ , we have that $G_{1}(x,\,y)=x$
, we have that $G_{1}(x,\,y)=x$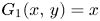 because $(x,\,y)\in l_{1}\subset L_{1}$
because $(x,\,y)\in l_{1}\subset L_{1}$ and $G_{2}(x,\,y)< y$
and $G_{2}(x,\,y)< y$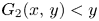 by (3.5). Analogously,
by (3.5). Analogously,

for all $(x,\,y)\in l_{2}\backslash \{(x^{*},\,y^{*}),\,(\frac {r_{2}}{\beta },\,0)\}$ . These expressions guarantee that
. These expressions guarantee that

Recall that the intervals $[0,\,A_{1}]\times \{0\}$ and $\{0\}\times [0,\,A_{2}]$
and $\{0\}\times [0,\,A_{2}]$ are positively invariant under $G$
are positively invariant under $G$ . Now, arguing as in the proof of proposition 2.1, we have that $G(\mathcal {A})\subset \mathcal {A}$
. Now, arguing as in the proof of proposition 2.1, we have that $G(\mathcal {A})\subset \mathcal {A}$ and
and

is an orientation preserving embedding. A critical fact is that

Thus, if we apply theorem 2.2 to $G|_{\mathcal {A}}$ , we conclude that for each $p\in \mathcal {A}$
, we conclude that for each $p\in \mathcal {A}$ , there exists $q\in Fix(G|_{\mathcal {A}})$
, there exists $q\in Fix(G|_{\mathcal {A}})$ so that
so that

On the other hand, we notice that $\frac {r_{2}}{\beta }>r_{1},\, \frac {r_{1}}{\alpha }>r_{2}$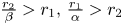 and (P) imply that
and (P) imply that

Repeating the argument of the proof of (i), we can prove $(r_{1},\,0)$ and $(0,\,r_{2})$
and $(0,\,r_{2})$ are local saddle points. Recall that the origin is always a local repeller in $(0,\,+\infty )^{2}$
are local saddle points. Recall that the origin is always a local repeller in $(0,\,+\infty )^{2}$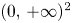 . Consequently, for all $p\in \mathcal {A}\cap (0,\,+\infty )^{2}$
. Consequently, for all $p\in \mathcal {A}\cap (0,\,+\infty )^{2}$ ,
,

Finally, we consider a sequence $\{(x_{n},\,y_{n})\}$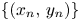 obtained from (3.1) so that $(x_{0},\,y_{0})\in \mathcal {R}\backslash \mathcal {A}$
obtained from (3.1) so that $(x_{0},\,y_{0})\in \mathcal {R}\backslash \mathcal {A}$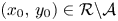 with $x_{0}>0$
with $x_{0}>0$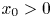 and $y_{0}>0$
and $y_{0}>0$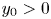 . By the same argument as that in lemma 3.2, one of the following facts holds:
. By the same argument as that in lemma 3.2, one of the following facts holds:
h1 $(x_{n},\,y_{n})$
tends to a fixed point of $G$.h2 $(x_{n},\,y_{n})\in \mathcal {A}$
for all $n\geq n_{0}$ with $n_{0}$ large enough.
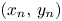



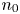
If h1 holds, then the fixed point has to be $(x^{*},\,y^{*})$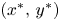 because the other fixed points are local repellers in $(0,\,+\infty )^{2}$
because the other fixed points are local repellers in $(0,\,+\infty )^{2}$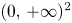 . If h2 holds, then we apply the above argument. The proof of (iii) is now completed.
. If h2 holds, then we apply the above argument. The proof of (iii) is now completed.
(iv) In this case, $G|_{\mathcal {R}}$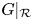 has four fixed points, namely, $(0,\,0)$
has four fixed points, namely, $(0,\,0)$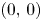 , $(r_{1},\,0)$
, $(r_{1},\,0)$ , $(0,\,r_{2})$
, $(0,\,r_{2})$ and $(x^{*},\,y^{*})$
and $(x^{*},\,y^{*})$ . By theorem 5.1 in [Reference Ruiz-Herrera32], we deduce that
. By theorem 5.1 in [Reference Ruiz-Herrera32], we deduce that

Obviously, $index (G|_{\mathcal {R}},\,(x^{*},\,y^{*}))=index (G,\,(x^{*},\,y^{*}))$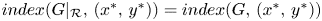 . Then, applying theorem 2.3 to $G|_{\mathcal {R}}$
. Then, applying theorem 2.3 to $G|_{\mathcal {R}}$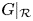 , we conclude that $G|_{\mathcal {R}}$
, we conclude that $G|_{\mathcal {R}}$ has trivial dynamics. Finally, linearizing the system and arguing as in (i), we conclude that $(r_{1},\,0)$
has trivial dynamics. Finally, linearizing the system and arguing as in (i), we conclude that $(r_{1},\,0)$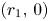 and $(0,\,r_{2})$
and $(0,\,r_{2})$ are local attractors.
are local attractors.
Example 3.6 Models with weak Allee effect.
The predation by a generalist predator with a saturating functional response is a common mechanism associated with the presence of weak Allee effects [Reference Kang and Yakubu20]. A natural extension in the plane of the single species models with this Allee effect is

where $\alpha,\,\beta >0,$

with $r>a>0$ and $b>0$
and $b>0$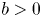 , see lemma 1.1 in [Reference Kang and Yakubu20].
, see lemma 1.1 in [Reference Kang and Yakubu20].
Although $g$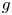 is not always monotone (see Fig. 1), we can apply theorem 3.5. Some particular choices of parameters are $r=0.5$
is not always monotone (see Fig. 1), we can apply theorem 3.5. Some particular choices of parameters are $r=0.5$ , $a=0.45$
, $a=0.45$ and $b=3$
and $b=3$ together with $\alpha =0.5$
together with $\alpha =0.5$ and $\beta =0.6$
and $\beta =0.6$ (i); $\alpha =1.1$
(i); $\alpha =1.1$ and $\beta =0.6$
and $\beta =0.6$ (iii); $\alpha =1.1$
(iii); $\alpha =1.1$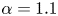 and $\beta =1.4$
and $\beta =1.4$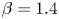 (iv). In general, condition (S) is difficult to verify in (3.7) because $\det D G(x,\,y)$
(iv). In general, condition (S) is difficult to verify in (3.7) because $\det D G(x,\,y)$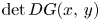 has a very complex expression. Notice that $det DG(x,\,y)$
has a very complex expression. Notice that $det DG(x,\,y)$ can be negative for some points $(x,\,y)\in \mathcal {R}$
can be negative for some points $(x,\,y)\in \mathcal {R}$ and for some choice of the parameters. Actually, system (3.7) can exhibit chaotic dynamics.
and for some choice of the parameters. Actually, system (3.7) can exhibit chaotic dynamics.

FIGURE 1. Representation of the function $g$ in system (3.7) with $r=0.5$
in system (3.7) with $r=0.5$ , $a=0.45$
, $a=0.45$ and $b=3$
and $b=3$ .
.
Example 3.7 A nonmonotone version of May–Oster model [Reference May and Oster24].
Consider the system

with $g(x)=\exp (x(r-x))$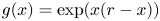 . After tedious computation, we can check that
. After tedious computation, we can check that

for all $(x,\,y)\in \mathcal {R}$ in the following range of parameters:
in the following range of parameters:
Case 1: $r=0.1$
, $\alpha \in (0.5,\,1)$ and $\beta \in (3,\,4.2)$.Case 2: $r=0.1$
, $\alpha \in (0.5,\,1)$ and $\beta \in (0.2,\,0.5)$.Case 3: $r=0.1$
, $\alpha \in (1,\,6)$ and $\beta \in (1,\,6)$.



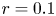

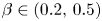



As a direct application of theorem 3.5, we obtain that $(0.1,\,0)$ is a global attractor in case 1, there is an interior fixed point $(x^{*},\,y^{*})$
is a global attractor in case 1, there is an interior fixed point $(x^{*},\,y^{*})$ that is a global attractor in case 2 and there is trivial dynamics with $(0.1,\,0)$
that is a global attractor in case 2 and there is trivial dynamics with $(0.1,\,0)$ and $(0,\,0.1)$
and $(0,\,0.1)$ as local attractors in case 3. As mentioned in the title of the example, (3.8) is a variant of the classical model discussed in [Reference May and Oster24]. We mention that this type of growth rates also appear in the evolution of climax species, see [Reference Franke and Yakubu7, Reference Franke and Yakubu9, Reference Franke and Yakubu10].
as local attractors in case 3. As mentioned in the title of the example, (3.8) is a variant of the classical model discussed in [Reference May and Oster24]. We mention that this type of growth rates also appear in the evolution of climax species, see [Reference Franke and Yakubu7, Reference Franke and Yakubu9, Reference Franke and Yakubu10].
Example 3.8 The conclusions in theorem 3.5 might not be true when the condition (S) does not hold.
Consider

with $g_1(x)=\frac {r}{x+a}$ and $g_2(x)=\exp (p-qx)$
and $g_2(x)=\exp (p-qx)$ , where $\alpha =\beta =a=1$
, where $\alpha =\beta =a=1$ , $r=21$
, $r=21$ , $p=2.5$
, $p=2.5$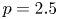 , $q=0.1$
, $q=0.1$ . In this case, we have $\mathcal {R}=[0,\,25]\times [0,\,25]$
. In this case, we have $\mathcal {R}=[0,\,25]\times [0,\,25]$ . One can check that
. One can check that

so condition (S) does not hold for model (3.9) because $(0,\,20)\in \mathcal {R}$ . As shown in [Reference Franke and Yakubu7], there is an asymptotically stable 2-cycle for this model so that the two species can coexist. This indicates that the conclusions in theorem 3.5 is not true for the model (3.9) by noticing that $\frac {r_{2}}{\beta }> r_{1}$
. As shown in [Reference Franke and Yakubu7], there is an asymptotically stable 2-cycle for this model so that the two species can coexist. This indicates that the conclusions in theorem 3.5 is not true for the model (3.9) by noticing that $\frac {r_{2}}{\beta }> r_{1}$ and $\frac {r_{1}}{\alpha }\leq r_{2}$
and $\frac {r_{1}}{\alpha }\leq r_{2}$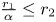 .
.
4. Extinction in planar systems beyond (3.1)
Predator–prey models are prototypes of Kolmogorov systems in which the results of § 3 are not directly applicable. However, some ideas developed in theorem 3.5 also work in these models. This shows the versatility of the mathematical framework given in § 2.
Consider the system

with $f_{1},\, f_{2}:[0,\,+\infty )^{2}\longrightarrow (0,\,+\infty )$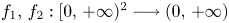 functions of class $\mathcal {C}^{1}$
functions of class $\mathcal {C}^{1}$ . We denote by
. We denote by

the map associated with (4.1). To model the predator–prey iteraction, we assume the following conditions ($x$ predator and $y$
predator and $y$ prey):
prey):
(PP1) $\frac {\partial F_{1}}{\partial x}(x,\,y)>0$
and $\frac {\partial F_{2}}{\partial y}(x,\,y)>0$ for all $(x,\,y)\in [0,\,+\infty )^{2}$.(PP2) $\frac {\partial f_{1}(x,\,y)}{\partial y}>0$
and $\frac {\partial f_{1}(x,\,y)}{\partial x}< 0$ for all $(x,\,y)\in [0,\,+\infty )^{2}$.(PP3) $\frac {\partial f_{2}(x,\,y)}{\partial x}<0$
and $\frac {\partial f_{2}(x,\,y)}{\partial y}< 0$ for all $(x,\,y)\in [0,\,+\infty )^{2}$.(LG) There are two fixed points $(p^{*},\,0)$
and $(0,\,q^{*})$ with $p^{*},\,q^{*}>0$ which are globally asymptotically stable on the $x$-axis and $y$-axis respectively.(PP4) There is a constant $\widetilde {K}>0$
so that $x f_{1}(x,\,q^{*}+1)< x$ for all $x>\widetilde {K}$.
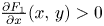
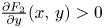

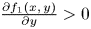


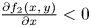


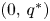

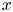

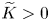
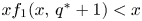

Condition (PP1) means that the intra-specific competition is contest (see [Reference Hassell15]). (PP2) indicates that the growth rate of the predator is the result of the conjunction of two biological facts: the intra-specific competition and the consumption of the prey. (PP3) has an analogous meaning for the prey. (LG) says that each species in isolation has logistic growth rate with carrying capacities $p^{*}$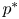 and $q^{*}$
and $q^{*}$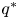 respectively. (PP4) says that the predator density decreases in the next generation when it is above a suitable threshold.
respectively. (PP4) says that the predator density decreases in the next generation when it is above a suitable threshold.
We say that system (4.1) is permanent if there are two constants $\varepsilon,\, M>0$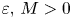 so that
so that

for all sequence $\{(x_{n},\,y_{n})\}$ of (4.1) with initial condition $(x_{0},\,y_{0})\in (0,\,+\infty )^{2}$
of (4.1) with initial condition $(x_{0},\,y_{0})\in (0,\,+\infty )^{2}$ . Informally speaking, the notion of permanence excludes the extinction of some species in the system. To apply the classical theory of permanent systems we need to recall the notion of absorbing set. We say that a positively invariant set $\mathcal {R}\subset [0,\,+\infty )^{2}$
. Informally speaking, the notion of permanence excludes the extinction of some species in the system. To apply the classical theory of permanent systems we need to recall the notion of absorbing set. We say that a positively invariant set $\mathcal {R}\subset [0,\,+\infty )^{2}$ is an absorbing set for (4.1) if for all $(x_{0},\,y_{0})\in [0,\,+\infty )^{2}$
is an absorbing set for (4.1) if for all $(x_{0},\,y_{0})\in [0,\,+\infty )^{2}$ , there is $n_{0}\in \mathbb {N}$
, there is $n_{0}\in \mathbb {N}$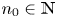 so that $F^{n}(x_{0},\,y_{0})\in \mathcal {R}$
so that $F^{n}(x_{0},\,y_{0})\in \mathcal {R}$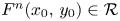 for all $n\geq n_{0}$
for all $n\geq n_{0}$ .
.
The following lemma is an immediate consequence of lemma 2.1 in [Reference Hutson and Moran19] by considering the average Liapunov function $V(x,\,y)=x^{\mu _1}y^{\mu _2}$ . See also theorem 3.1 in [Reference Ruiz-Herrera29].
. See also theorem 3.1 in [Reference Ruiz-Herrera29].
Lemma 4.1 Assume that the map $F$ associated with (4.1) has an absorbing set $\mathcal {R}:=[0,\,K_1]\times [0,\,K_2]$
associated with (4.1) has an absorbing set $\mathcal {R}:=[0,\,K_1]\times [0,\,K_2]$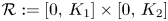 with $K_1,\,K_2>0$
with $K_1,\,K_2>0$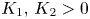 . If there are real numbers $\mu _1,\,\mu _2>0$
. If there are real numbers $\mu _1,\,\mu _2>0$ such that
such that

for each $q$ in $\Omega (\partial \mathcal {R}):=\displaystyle \bigcup _{p\in \partial \mathcal {R}}\omega (F,\,p),$
in $\Omega (\partial \mathcal {R}):=\displaystyle \bigcup _{p\in \partial \mathcal {R}}\omega (F,\,p),$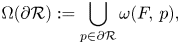 then (4.1) is permanent.
then (4.1) is permanent.
Next we give the main result of this section.
Theorem 4.2 Suppose that (PP1)–(PP4) and (LG) hold. Then every sequence of (4.1) is bounded. Moreover, we have the following conclusion:
(i) If $f_{2}(p^{*},\,0)>1,$
(4.1) is permanent.(ii) If $f_{2}(p^{*},\,0)\leq 1,$
$(p^{*},\,0)$ is a global attractor in $(0,\,+\infty )^{2}$.



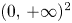
Proof. First, let us prove that $\mathcal {R}=[0,\,\widetilde {K}]\times [0,\,q^{*}+1]$ is an absorbing set for (4.1). Take $\{(x_{n},\,y_{n})\}$
is an absorbing set for (4.1). Take $\{(x_{n},\,y_{n})\}$ a sequence of (4.1). Define $\phi (y)=F_{2}(0,\,y)$
a sequence of (4.1). Define $\phi (y)=F_{2}(0,\,y)$ . It follows from (PP1) and (PP3) that $\phi (y)$
. It follows from (PP1) and (PP3) that $\phi (y)$ is strictly increasing and $\phi (y)\geq F_{2}(x,\,y)$
is strictly increasing and $\phi (y)\geq F_{2}(x,\,y)$ for all $(x,\,y)\in [0,\,+\infty )^{2}$
for all $(x,\,y)\in [0,\,+\infty )^{2}$ . From these properties we can deduce that
. From these properties we can deduce that

for all $n\in \mathbb {N}$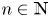 . Indeed,
. Indeed,

Repeating this argument, we obtain that

Using that $\phi$ is increasing and the previous step,
is increasing and the previous step,

We complete the argument after a simple induction. Since $q^{*}>0$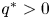 is a global attractor of
is a global attractor of

in $(0,\,+\infty )$ by (LG) it is clear that there exists $n_{1}\in \mathbb {N}$
by (LG) it is clear that there exists $n_{1}\in \mathbb {N}$ so that $y_{n}\in [0,\,q^{*}+1]$
so that $y_{n}\in [0,\,q^{*}+1]$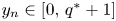 for all $n\geq n_{1}$
for all $n\geq n_{1}$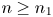 by (4.3). Next we define $\varphi (x)=F_{1}(x,\,q^{*}+1)$
by (4.3). Next we define $\varphi (x)=F_{1}(x,\,q^{*}+1)$ . By (PP1), this function is strictly increasing. Using (PP4), we have that for all $z_{0}\in [0,\,+\infty )$
. By (PP1), this function is strictly increasing. Using (PP4), we have that for all $z_{0}\in [0,\,+\infty )$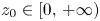 , the sequence $\{z_{n}\}$
, the sequence $\{z_{n}\}$ obtained from
obtained from

satisfies that $z_{n}\longrightarrow q$ with $q<\widetilde {K}$
with $q<\widetilde {K}$ . We also know by (PP2) that $F_{1}(x_{n},\,y_{n})\leq F_{1}(x_{n},\,q^{*}+1)=\varphi (x_{n})$
. We also know by (PP2) that $F_{1}(x_{n},\,y_{n})\leq F_{1}(x_{n},\,q^{*}+1)=\varphi (x_{n})$ for all $n\geq n_{1}$
for all $n\geq n_{1}$ . Arguing as above, we can find $n_{2}\geq n_{1}$
. Arguing as above, we can find $n_{2}\geq n_{1}$ so that
so that

for all $n\geq n_{2}$ . Now, it is clear that $[0,\,\widetilde {K}]\times [0,\,q^{*}+1]$
. Now, it is clear that $[0,\,\widetilde {K}]\times [0,\,q^{*}+1]$ is an absorbing set for (4.1). Next we properly prove the theorem.
is an absorbing set for (4.1). Next we properly prove the theorem.
(i) It follows from (LG) that

Note that $f_1(p^{*},\,0)=1$ and $f_2(0,\,q^{*})=1$
and $f_2(0,\,q^{*})=1$ because $(p^{*},\,0)$
because $(p^{*},\,0)$ and $(0,\,q^{*})$
and $(0,\,q^{*})$ are fixed points. Then, (PP2) and (PP3) imply that $f_i(0,\,0)>1$
are fixed points. Then, (PP2) and (PP3) imply that $f_i(0,\,0)>1$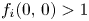 , $i=1,\,2$
, $i=1,\,2$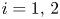 , and $f_1(0,\,q^{*})>1$
, and $f_1(0,\,q^{*})>1$ . We know by assumptions that $f_{2}(p^{*},\,0)>1$
. We know by assumptions that $f_{2}(p^{*},\,0)>1$ . At this moment, it is clear there are $\mu _1,\,\mu _2>0$
. At this moment, it is clear there are $\mu _1,\,\mu _2>0$ such that (4.2) holds for each $q\in \{(0,\,0),\,(p^{*},\,0),\,(0,\,q^{*})\}$
such that (4.2) holds for each $q\in \{(0,\,0),\,(p^{*},\,0),\,(0,\,q^{*})\}$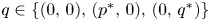 . Hence (4.1) is permanent by lemma 4.1. (ii) It suffices to prove that every sequence of (4.1) with initial condition in $\mathcal {R}$
. Hence (4.1) is permanent by lemma 4.1. (ii) It suffices to prove that every sequence of (4.1) with initial condition in $\mathcal {R}$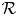 converges to $(p^{*},\,0)$
converges to $(p^{*},\,0)$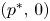 . First we prove that (4.1) does not admit fixed points in $(0,\,+\infty )^{2}$
. First we prove that (4.1) does not admit fixed points in $(0,\,+\infty )^{2}$ . Assume, by contradiction, that $(p,\,q)\in (0,\,+\infty )^{2}$
. Assume, by contradiction, that $(p,\,q)\in (0,\,+\infty )^{2}$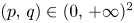 is a fixed point of $F$
is a fixed point of $F$ . Then,
. Then,

By (PP2), we obtain that

Consequently, $p>p^{*}$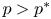 . On the other hand, by (PP3), we deduce that
. On the other hand, by (PP3), we deduce that

a contradiction. After simple computations, we can prove that

for all $(x,\,y)\in [0,\,+\infty )^{2}$ . Moreover, it is clear that $F^{-1}(\{0\})=\{0\}$
. Moreover, it is clear that $F^{-1}(\{0\})=\{0\}$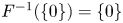 . Then, by proposition 2.1, we have that
. Then, by proposition 2.1, we have that

is an orientation preserving embedding. We notice that

Moreover, $f_{1}(0,\,0)>1$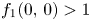 , $f_{2}(0,\,0)>1$
, $f_{2}(0,\,0)>1$ and $f_{1}(0,\,q^{*})>1$
and $f_{1}(0,\,q^{*})>1$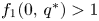 , (see the proof of (i)). Linearizing the system, we easily obtain that $(0,\,0)$
, (see the proof of (i)). Linearizing the system, we easily obtain that $(0,\,0)$ is a local repeller. Regarding $(0,\,q^{*})$
is a local repeller. Regarding $(0,\,q^{*})$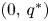 , we have that $\{f_{1}(0,\,q^{*}),\,1+q^{*} \frac {\partial f_{2}}{\partial y}(0,\,q^{*})\}$
, we have that $\{f_{1}(0,\,q^{*}),\,1+q^{*} \frac {\partial f_{2}}{\partial y}(0,\,q^{*})\}$ are the eigenvalues of the linearized system. By (PP1) and (PP3), $0<\frac {\partial F_{2}}{\partial y}(0,\,q^{*})=1+q^{*} \frac {\partial f_{2}}{\partial y}(0,\,q^{*})<1$
are the eigenvalues of the linearized system. By (PP1) and (PP3), $0<\frac {\partial F_{2}}{\partial y}(0,\,q^{*})=1+q^{*} \frac {\partial f_{2}}{\partial y}(0,\,q^{*})<1$ . In other words, $(0,\,q^{*})$
. In other words, $(0,\,q^{*})$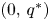 is a saddle point so that the repelling direction is $(w_{1},\,w_{2})$
is a saddle point so that the repelling direction is $(w_{1},\,w_{2})$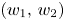 with $w_{1}>0$
with $w_{1}>0$ . The conclusion follows from theorem 2.2.
. The conclusion follows from theorem 2.2.
There are many classical models that satisfy conditions (PP1)–(PP4) and (LG). For example,

where $r>1+h_{1}(0)$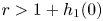 , $s>1+h_{2}(0)$
, $s>1+h_{2}(0)$ and the functions $h_{1},\,h_{2}:[0,\,+\infty )\longrightarrow (0,\,+\infty )$
and the functions $h_{1},\,h_{2}:[0,\,+\infty )\longrightarrow (0,\,+\infty )$ are of class $\mathcal {C}^{1}$
are of class $\mathcal {C}^{1}$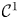 with $h_{1}'(x)<0$
with $h_{1}'(x)<0$ and $h_{2}'(x)>0$
and $h_{2}'(x)>0$ for all $x\in (0,\,+\infty )$
for all $x\in (0,\,+\infty )$ .
.
5. Discussion
It is well known that the relative position of the nullclines determines the dynamical behaviour of the classical Lotka–Volterra model

In this paper we have proved that the same results remain true in a broad family of discrete systems, namely

In our analysis, we have imposed mainly the following:
(P) For $i=1,\,2$
, there exists $r_{i}>0$ so that $g_{i}(x)>1$ if $x\in (0,\,r_{i})$ and $g_{i}(x)<1$ if $x>r_{i}$.(S) $\det DG(x,\,y)>0$
for all $(x,\,y)\in \mathcal {R}$ with $\mathcal {R}=[0,\, A_{1}]\times [0,\,A_{2}]$ with
\[ A_{1}=\max\left\{r_{1},\frac{r_{2}}{\beta}\right\},\quad A_{2}=\max\left\{r_{2},\frac{r_{1}}{\alpha}\right\} \]and $G(x,\,y)=(x g_{1}(x+\alpha y),\,y g_{2}(y+\beta x))$.
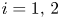


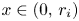

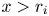


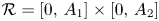


The role of these conditions is critical. Notice that (P) is a necessary condition to maintain the dynamical behaviour of (5.1) in (5.2). On the other hand, if we drop (S), as discussed in § 3, new phenomena emerge in (5.2) in comparison with (5.1). For example, the presence of 2-cycles or chaotic dynamics. It is worth stressing that condition (P) encompasses functions that are not monotone. Particularly, we can describe the dynamical behaviour of species subject to Allee effects. Other marked examples are the population models for pioneer-climax species that appears when $g_{1}$ is decreasing and $g_{2}$
is decreasing and $g_{2}$ is one-humped, see [Reference Harper14, Reference Kim and Marlin21, Reference Selgrade and Namkoong33, Reference Sumner34]. For a long time, the dominant topic in these models was mainly the exclusion of the pioneer species, ignoring other dynamical patterns. In contrast with this point of view, theorem 3.5 describes all the possible dynamical patterns. In particular, we study the exclusion for the climax species, which has also been analysed recently by Gilbertson and Kot [Reference Gilbertson and Kot11].
is one-humped, see [Reference Harper14, Reference Kim and Marlin21, Reference Selgrade and Namkoong33, Reference Sumner34]. For a long time, the dominant topic in these models was mainly the exclusion of the pioneer species, ignoring other dynamical patterns. In contrast with this point of view, theorem 3.5 describes all the possible dynamical patterns. In particular, we study the exclusion for the climax species, which has also been analysed recently by Gilbertson and Kot [Reference Gilbertson and Kot11].
There are many models in population dynamics that are not of the form (5.2), i.e., the growth rates are not a scalar function composed by a linear combination of the densities of the species. Nevertheless, many results are valid when the map associated with the model satisfies condition (S). This is the case of most predator–prey systems with a generalist predator. For these models, we have proved that the absence of coexistence states leads to the exclusion of the prey. A direct consequence of this is that the presence of any oscillatory behaviour in $(0,\,+\infty )^{2}$ implies the existence of a coexistence state.
implies the existence of a coexistence state.
Acknowledgments
The authors would like to thank the editor and the anonymous referees for their feedback and comments, which improved the quality of the manuscript. The work of L. Niu was supported by the National Natural Science Foundation of China (NSFC) under Grant No. 12001096 and the Fundamental Research Funds for the Central Universities (No. 2232021G-13).






 Open access
Open access