No CrossRef data available.
Published online by Cambridge University Press: 29 November 2023
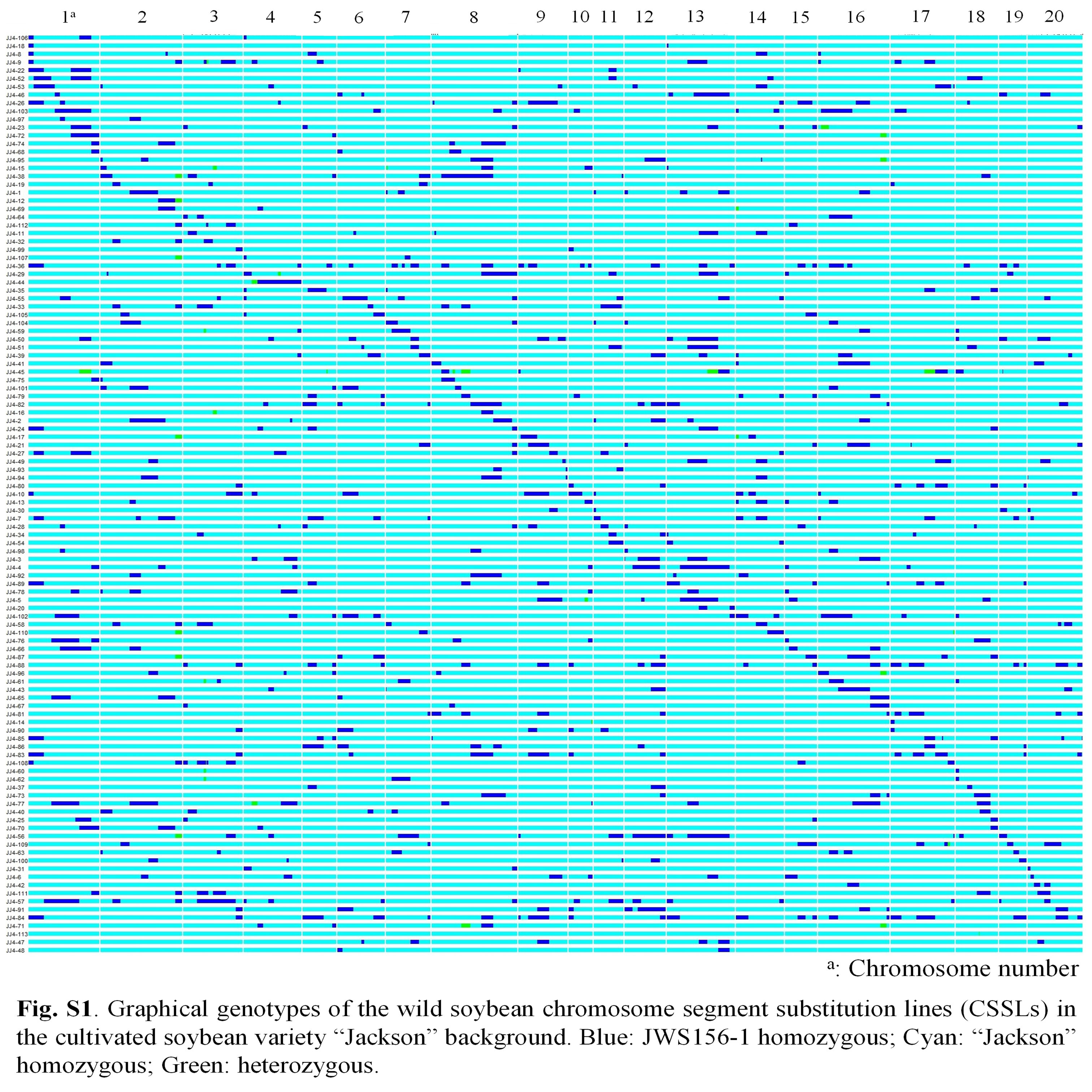
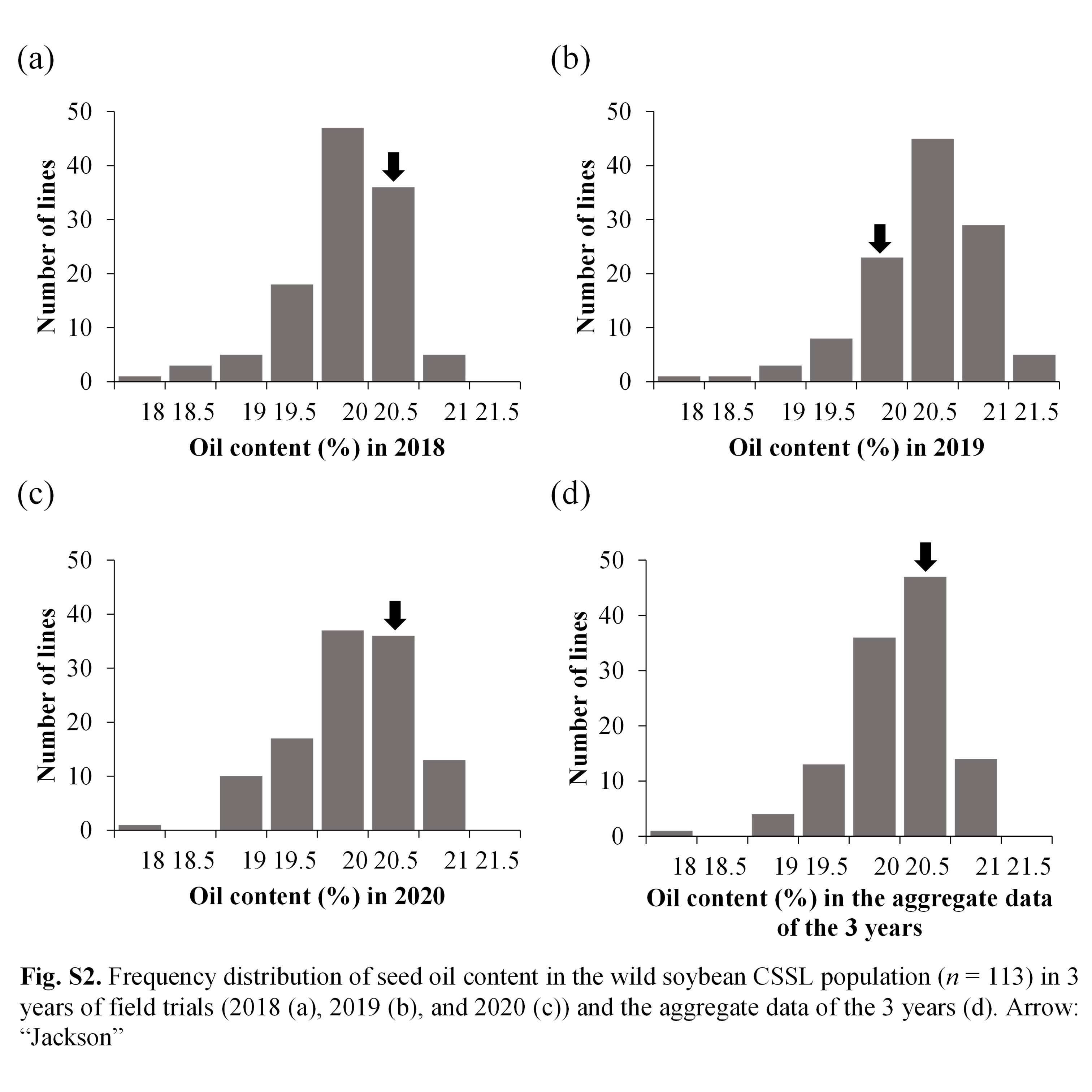
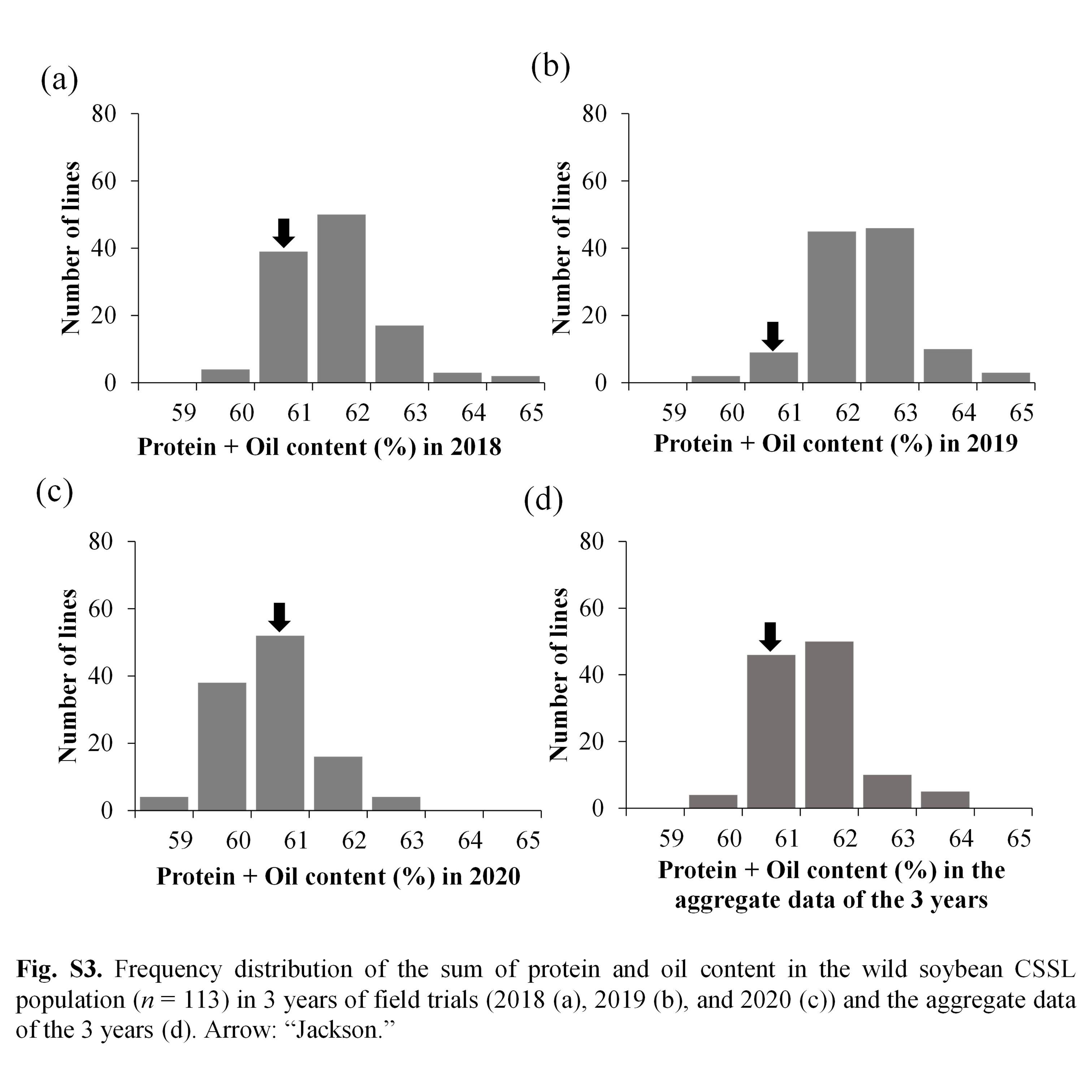
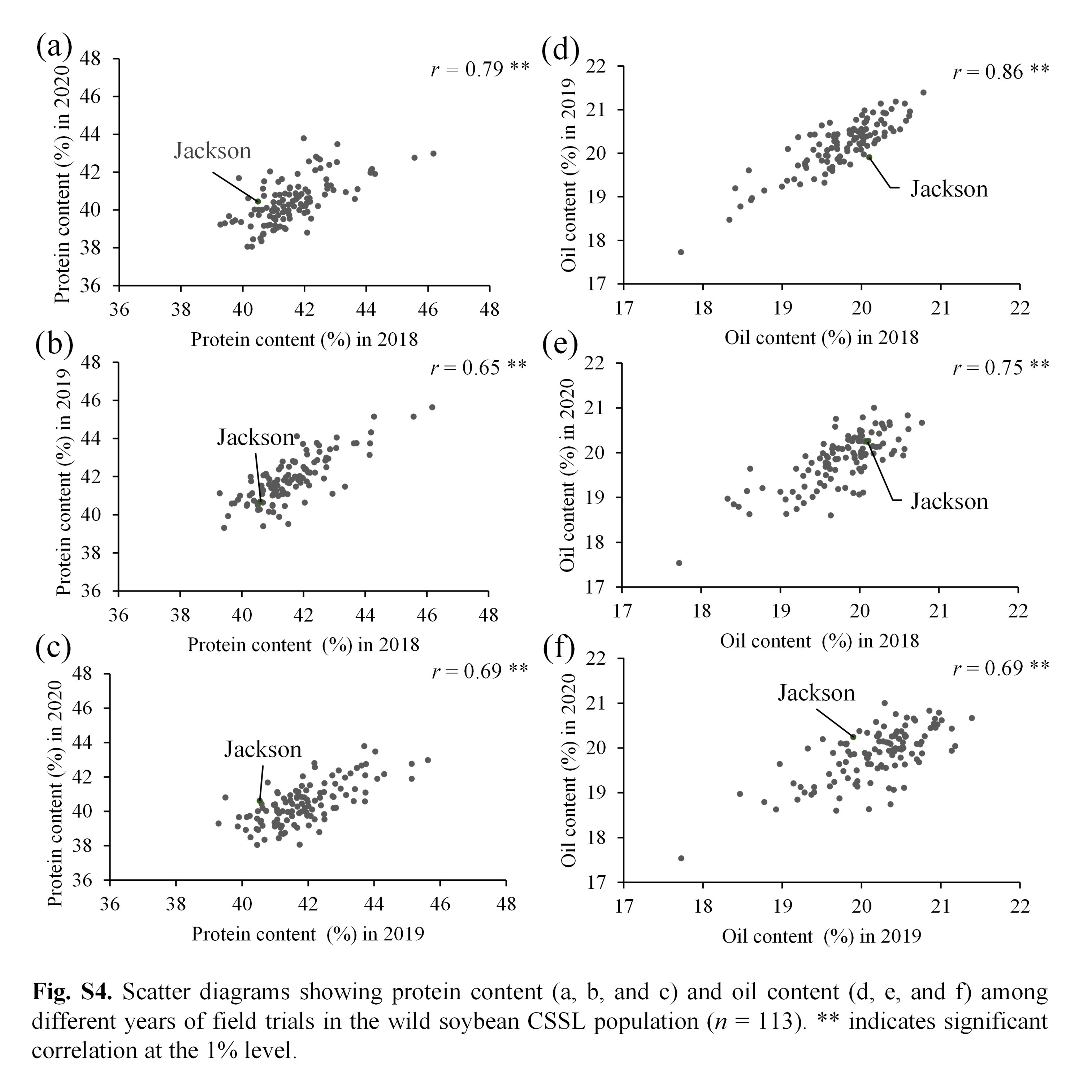
Soybean is one of the chief crops producing protein and oil for human consumption. Wild soybean, the ancestor of cultivated soybean, possesses high seed protein content; therefore, it is a valuable genetic resource that could enhance protein content in the cultivated varieties. To identify the genes responsible for increasing protein content in wild soybean, a population comprising 113 BC4F6 chromosome segment substitution lines (CSSL) was developed from a cross between soybean cultivar ‘Jackson’ and wild soybean accession JWS156-1. The CSSL population was cultivated in the field conditions for 3 years (2018, 2019 and 2020), and the seeds harvested from each line were analysed for protein and oil contents by InfraTec Nova instrument. Quantitative trait locus (QTL) analysis with 243 SSR markers identified 12 QTLs associated with seed protein, oil and protein + oil contents. Among these QTLs, qPro8 and qPro19, two major and stable QTLs for protein content, were detected on chromosomes 8 and 19, respectively. No QTL for oil content was detected in the vicinity of qPro19, indicating that qPro19 did not influence the seed oil content. The effect of qPro19 was validated using near-isogenic lines (NILs) of qPro19. By introducing the qPro19 allele from wild soybean into another soybean variety, ‘Tachiyutaka’, a BC4 line, T-678, that showed enhanced seed protein content, without reducing the seed oil content. This study implied that the qPro19 allele from wild soybean could be a potential genetic resource for breeding programmes aimed to improve soybean seed quality.