No CrossRef data available.
Published online by Cambridge University Press: 15 December 2023
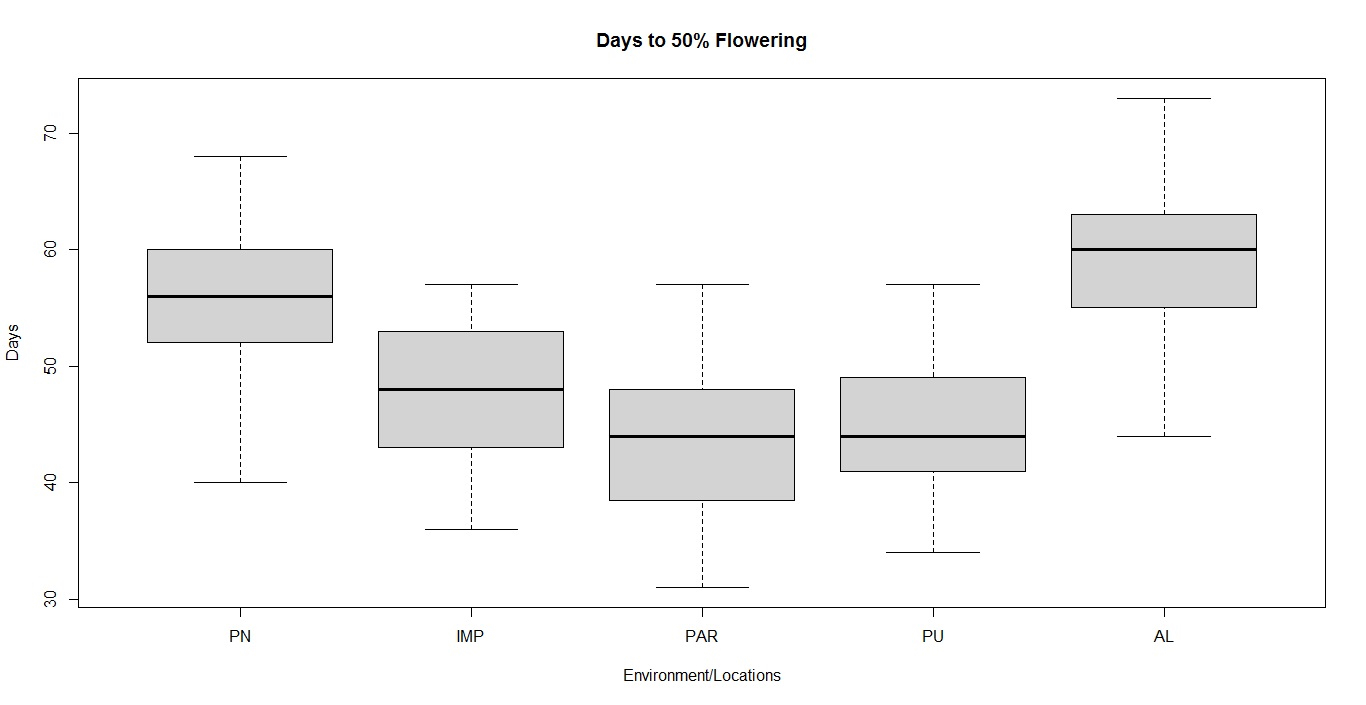
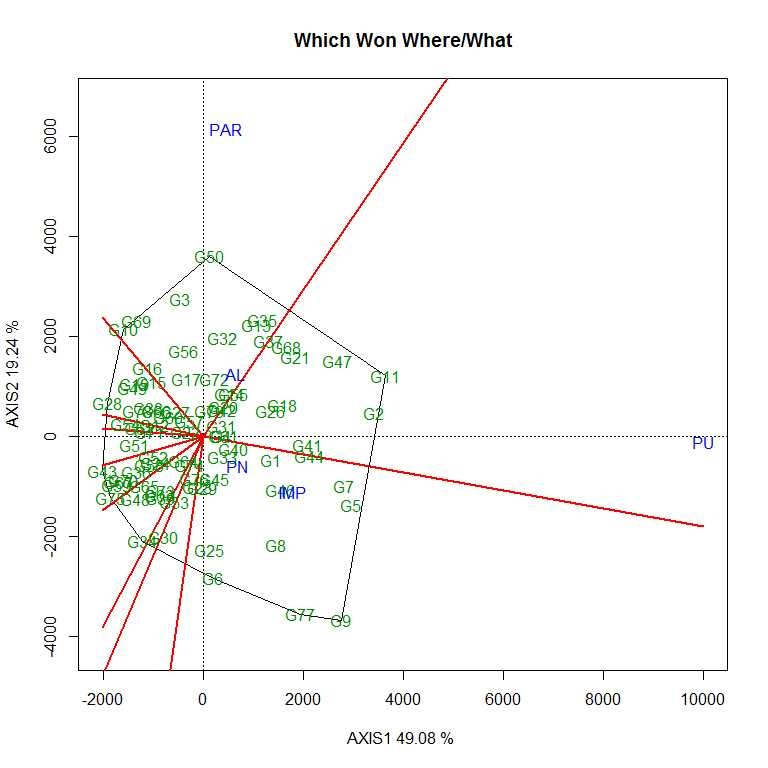
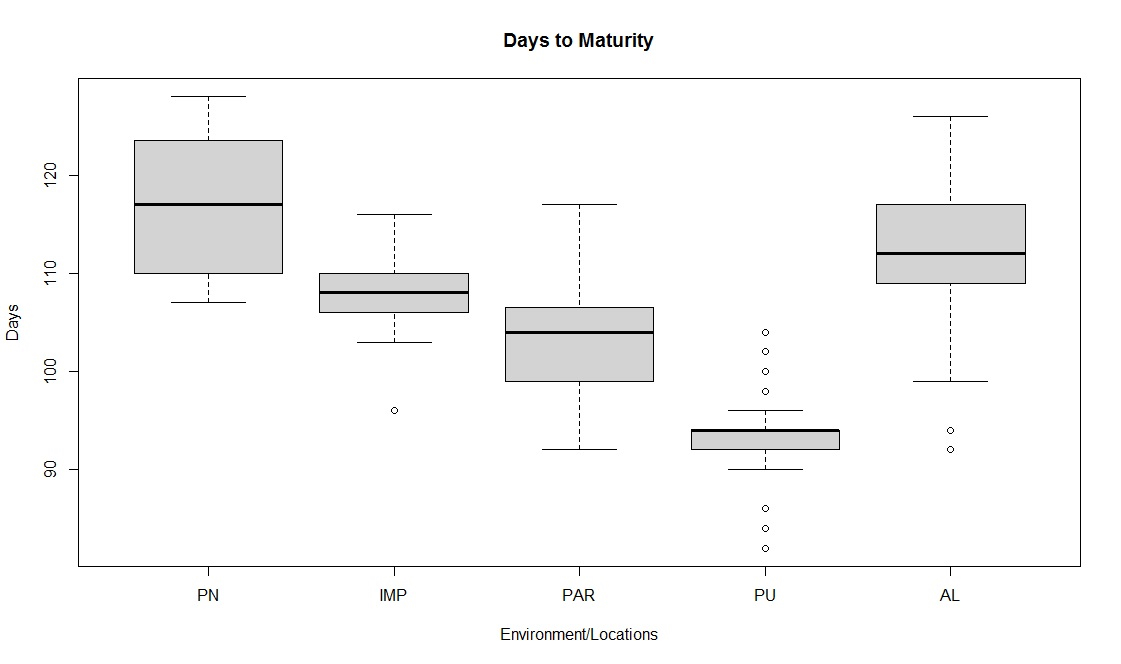
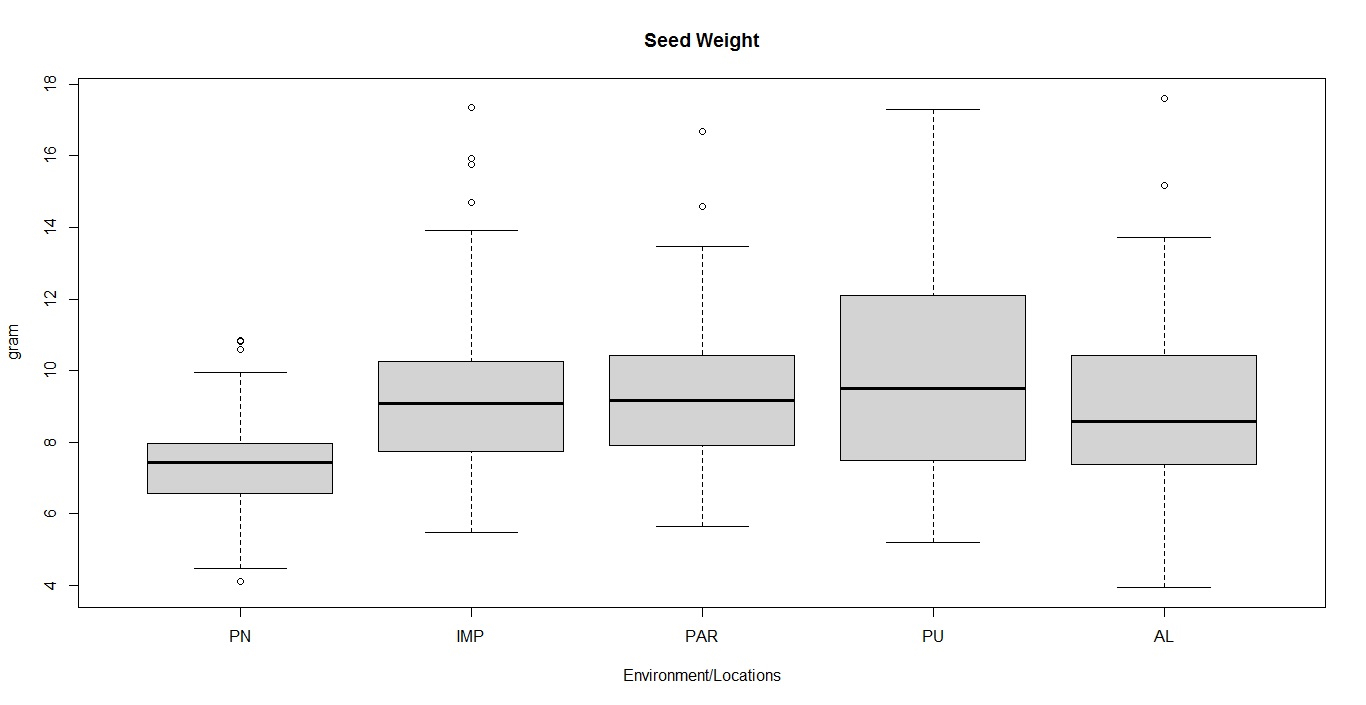
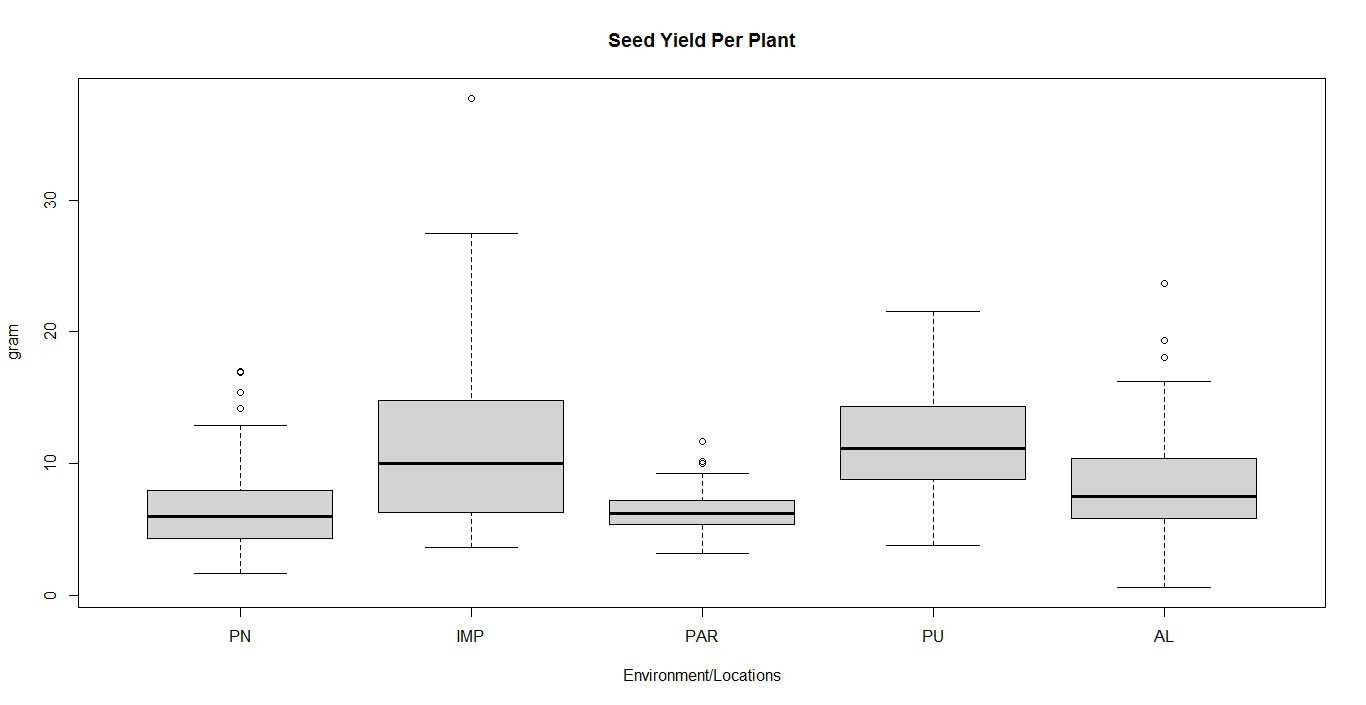
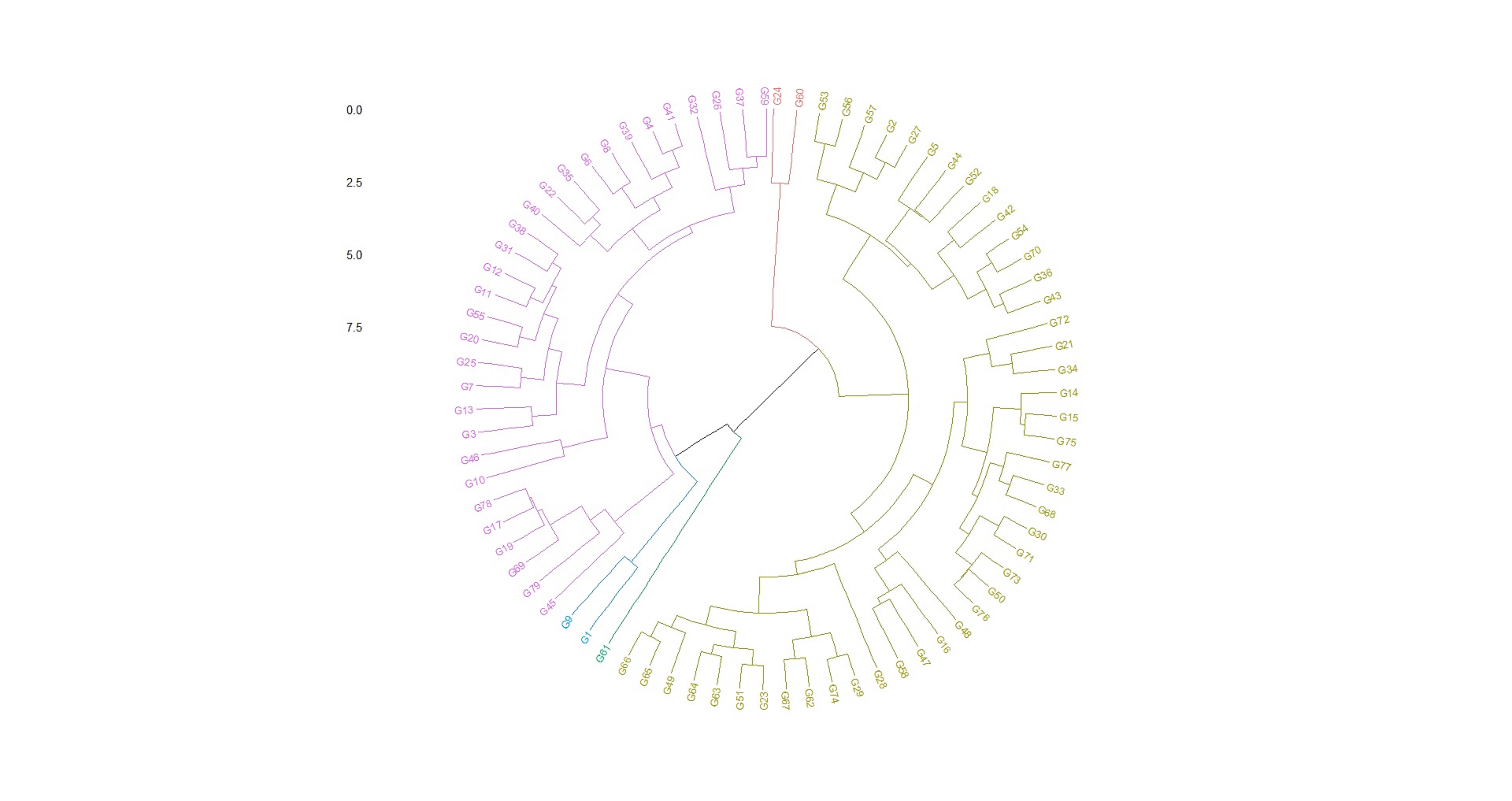
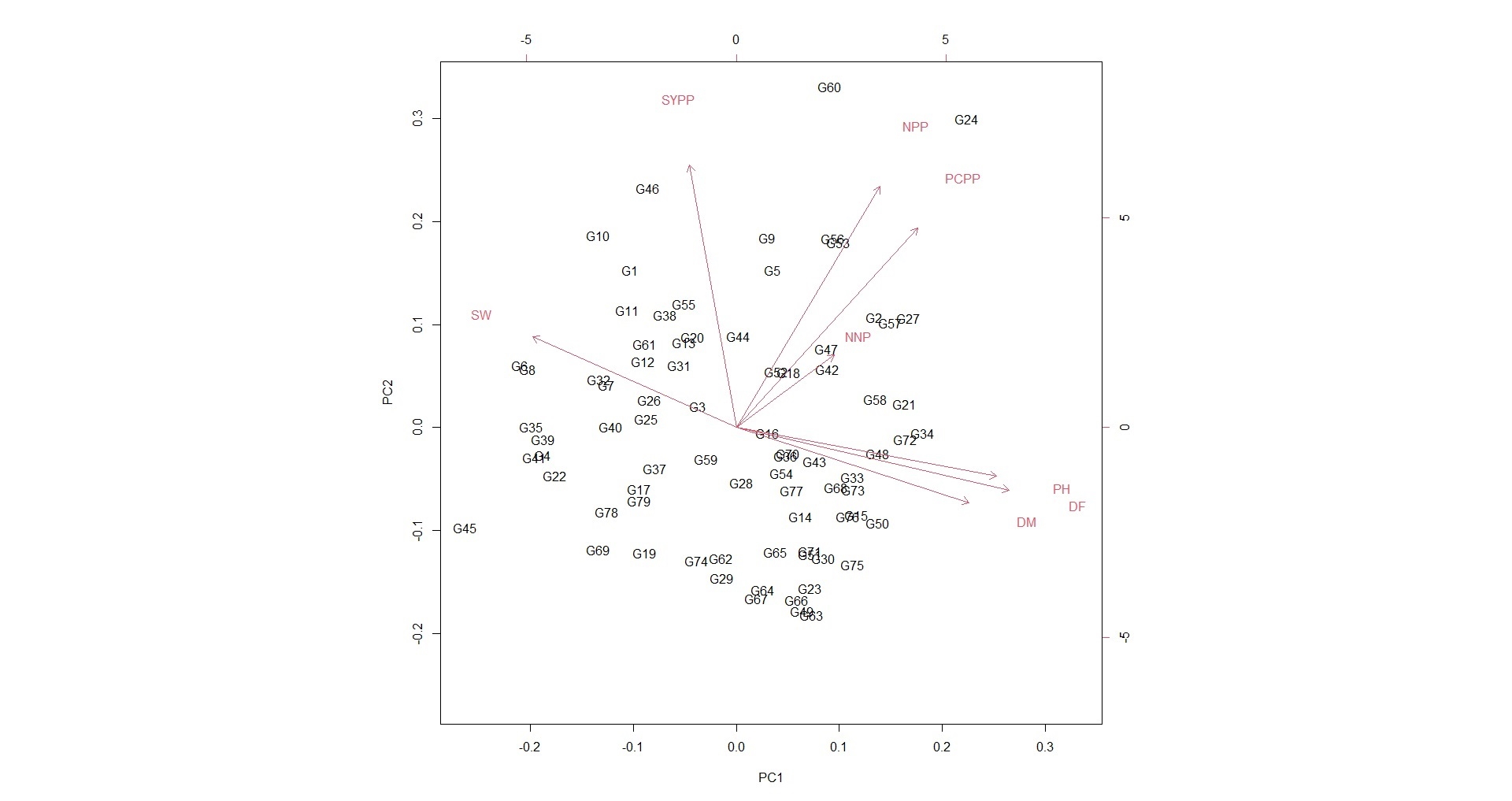
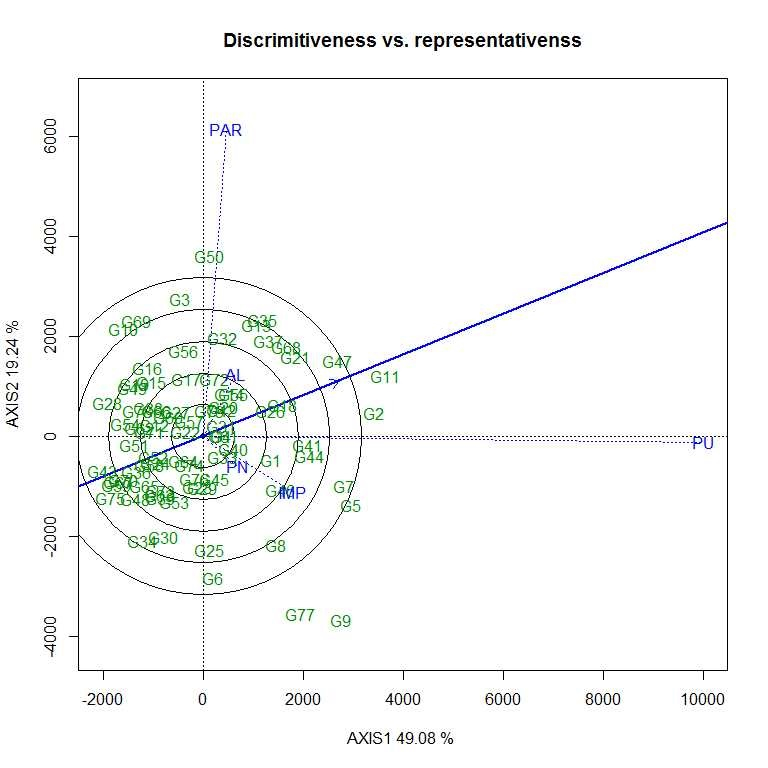
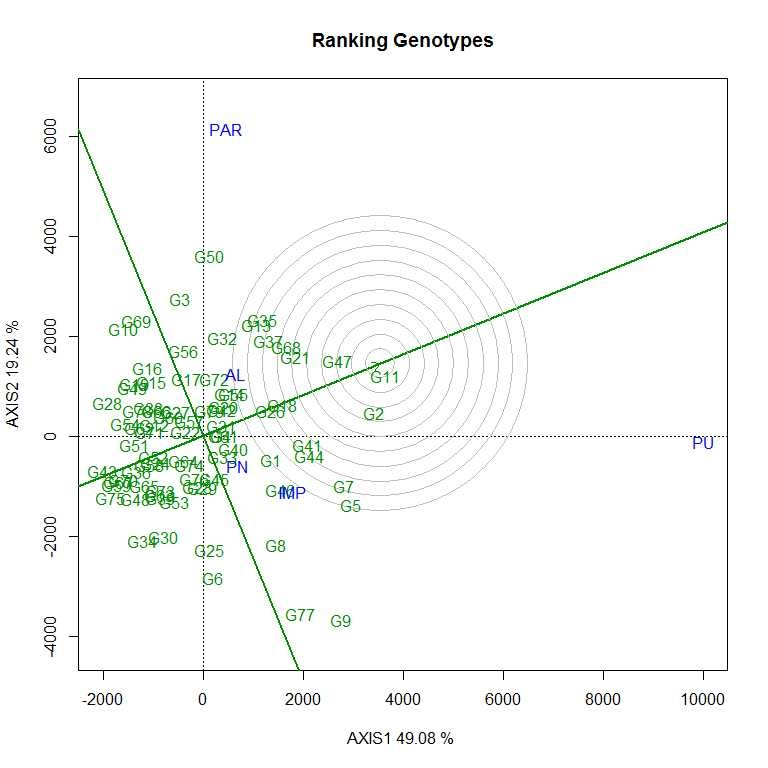
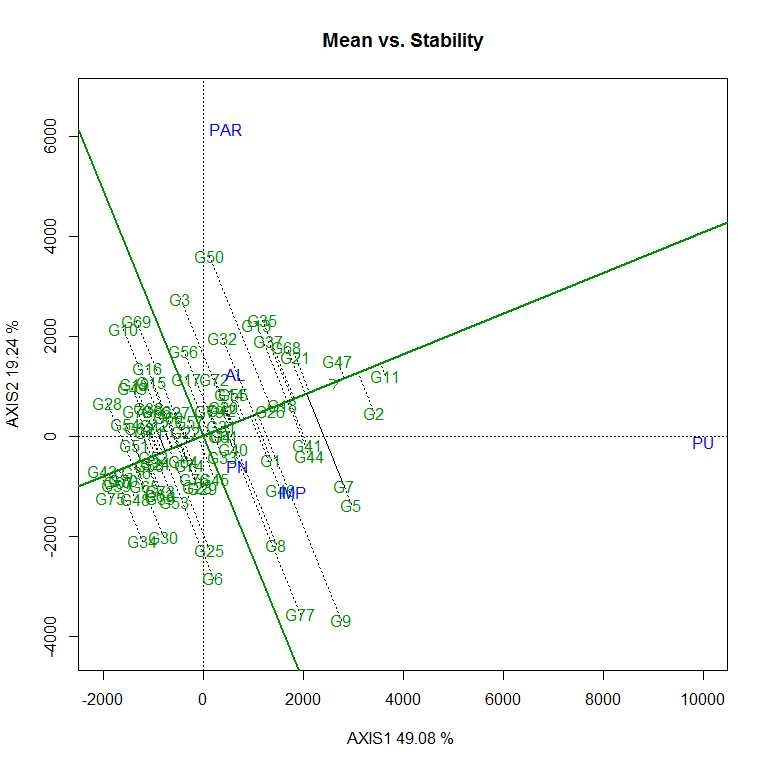
Soybean is a major source of vegetable oil and protein worldwide. Globally, India is among the top five producers where soybean is a major oilseed grown under diverse agro-climatic conditions by small and marginal farmers. The present study aims to identify soybean varieties with higher yield levels, resistance to pestdiseases and adaptability to climatic fluctuations. One hundred and twenty-five (125) indigenous and exotic soybean germplasm accessions and five checks were evaluated and characterized for eight agro-morphological traits at five testing locations and also screened for frog-eye leaf spot (FLS) and yellow mosaic virus (YMV) diseases under hot-spot locations during the rainy season. A wide range of variability was observed among accessions for days to 50% flowering (39–59), plant height (41–111 cm), number of nodes/plant (10–30), pod clusters/plant (14–39), number of pods/plant (40–102), days to maturity (96–115), grain yield/plant (4.89–16.54 g) and 100-seed weight (6.02–13.72 g). Among various traits, 100-seed weight (0.45), number of pods/plant (0.60) and number of pod clusters/plant (0.38) were found to be major yield-contributing traits as they exhibited highly significant correlation with grain yield/plant. Principal components PCI and PCII with eigen value >1 accounted for 42.66 and 27.08% of the total variation, respectively. Accessions G24 (EC 393222) from Taiwan and G40 (IMP-1) from the USA belonging to cluster IV were found promising for multiple yield traits and JS 20–38 from cluster III for earliness as per cluster analysis. GGE biplot average environment coordination (AEC) view revealed that the accessions viz., G11 (EC 333872), G2 (EC 251506) and G47 (TNAU-S-55) were the best performing stable genotypes in terms of grain yield/plant across locations. Twelve accessions had a high level of resistance against both FLS and YMV diseases under natural hot-spot conditions which can be utilized as promising donors in the soybean breeding programme.