



No CrossRef data available.
Article contents
Characterization of Western Himalayan small-seeded red beans (Phaseolus vulgaris L.) for yield, quality and resilience
Published online by Cambridge University Press: 17 April 2023
Abstract
Western Himalayan region is a rich repository of common bean genetic resources evolved over generations of farmer selection. The bean variability encompasses various use categories such as dry, snap and shelled beans. The present study aimed to characterize 40 small-seeded red common bean genotypes collected from different districts of Kashmir valley as well as Poonch, Rajouri and Bhaderwah areas of Jammu which are world famous for their beans. Based on 5-year evaluation, there was substantial variability for various qualitative traits as well as quantitative traits. Among variability parameters, higher PCV and GCV values were recorded for plant height, seed yield per plant and number of pods per plant and low for days to maturity, seed length and pod length. High heritability was recorded for all the traits ranging from 78.72 to 94.87%. There was substantial variability in root traits scored in laboratory and greenhouse as well as canopy temperature depression and relative water content. Various genotypes were found resistant to bean common mosaic virus (WB-N-1, WB-N-11, WB-1318, WB-401, WB-451, WB-1634); anthracnose (WB-1634 WB-83, WB-258, WB-1282, WB-1304); angular leaf spot (WB-6, WB-1634, WB-258, WB-451, N-1 and N-10); Fusarium wilt (WB-341, WB-451, WB-N-1 and WB-1587) and Ascochyta blight (WB-6). Similarly WB-92 was highly resistant to shattering. The genotypes WB-341, WB-1634, WB-N-1 performed better under water stress, WB-N-1 and WB-1634 were resistant to multiple diseases and WB-1634 had better cooking quality. The genotypes identified can be used in bean improvement programmes and have also been submitted to NBPGR for long-term storage.
- Type
- Research Article
- Information
- Copyright
- Copyright © The Author(s), 2023. Published by Cambridge University Press on behalf of NIAB
References
Akbulut, B, Karakurt, Y and Tonguc, M (2013) Molecular characterization of common bean (Phaseolus vulgaris L.) genotypes. Akdeniz University Journal Faculty of Agriculture 26, 105–108.Google Scholar
Amin, T (2018) Effect of planting techniques and organic amendments on the management of Fusarium Wilt of common bean (MSc thesis). submitted to SKUAST-Kashmir. 88p.Google Scholar
Asfaw, A, Blair, MW and Almekinders, C (2009) Genetic diversity and population structure of common bean (Phaseolus vulgaris L.) landraces from the East African highlands. Theoretical and Applied Genetics 120, 1–12.10.1007/s00122-009-1154-7CrossRefGoogle ScholarPubMed
Banoo, A, Nabi, A, Rasool, RS, Shah, MD, Ahmad, M, Sofi, PA and Padder, BA (2020) North-western Himalayan common beans: population structure and mapping of quantitative anthracnose resistance through genome wide association study. Frontiers in Plant Sciences 11, 1459.Google ScholarPubMed
Barr, HD and Weatherley, PE (1962) A re-examination of the relative turgidity technique for estimating water deficit in leaves. Australian Journal of Biological Sciences 15, 413–428.10.1071/BI9620413CrossRefGoogle Scholar
Beebe, S, Rengifo, J, Gaitan, E, Duque, MC and Tohme, J (2001) Diversity and origin of Andean landraces of common bean. Crop Science 41, 854–862.10.2135/cropsci2001.413854xCrossRefGoogle Scholar
Bishnoi, S and Khetarpaul, N (1993) Variability in physico-chemical properties and nutrient composition of different pea cultivars. Food Chemistry 47, 371–373.10.1016/0308-8146(93)90179-JCrossRefGoogle Scholar
Black, CA (1965) Methods of Soil Analysis: Part I. Physical and Mineralogical Properties. Madison, WI: American Society of Agronomy.10.2134/agronmonogr9.1CrossRefGoogle Scholar
Burton, GW and Devane, EH (1953) Estimating heritability in tall fescue (Festuca arundinacea) from replicated clonal material 1. Agronomy Journal 45, 478–481.CrossRefGoogle Scholar
Chavez-Servia, JL, Heredia-García, E, Mayek-Perez, N, Aquino-Bolaños, EN, Hernández-Delgado, S, Carrillo-Rodríguez, JC and Vera-Guzmán, AM (2016) Diversity of common bean (Phaseolus vulgaris L.) landraces and the nutritional value of their grains. In Goyal, AK (ed.), Grain Legumes. London, UK: IntechOpen, pp. 1–33.Google Scholar
Choudhary, N, Hamid, A, Singh, B, Khandy, I, Sofi, PA, Bhat, MA and Mir, RR (2017) Insight into the origin of common bean (Phaseolus vulgaris L.) grown in the state of Jammu and Kashmir of north-western Himalayas. Genetic Resources and Crop Evolution 65, 963–977.10.1007/s10722-017-0588-zCrossRefGoogle Scholar
Choudhary, N, Bawa, V, Paliwal, R, Singh, B, Bhat, MA, Mir, JI, Gupta, M, Sofi, PA, Thudi, M, Varshney, RK and Mir, RR (2018) Gene/QTL discovery for Anthracnose in common bean (Phaseolus vulgaris L.) from North-western Himalayas. PLoS ONE 13, e0191700.10.1371/journal.pone.0191700CrossRefGoogle ScholarPubMed
Correa, PC, Goneli, ALD, Júnior, PCA, De Oliveira, GHH and Valente, DSM (2010) Moisture sorption isotherms and isosteric heat of sorption of coffee in different processing levels. International Journal of Food Science and Technology 45, 2016–2022.10.1111/j.1365-2621.2010.02373.xCrossRefGoogle Scholar
Dar, WA, Parrey, EA and Khan, MM (2020) Evaluation of bean germplasm lines for resistance to Angular Leaf spot (Phaseoreopsis griseola) under high altitude conditions of Gurez. Journal of Plant Pathology and Microbiology 11, 509–511.Google Scholar
Drijfhout, E and Davis, JHC (1989) Selection of a new set of homogeneously reacting bean (Phaseolus vulgaris) differentials to differentiate races of Colletotrichum lindemuthianum. Plant Pathology 38, 391–396.CrossRefGoogle Scholar
Drijfhout, E, Silbernagel, MJ and Burke, DW (1978) Differentiation of strains of bean common mosaic virus. Netherlands Journal of Plant Pathology 84, 13–26.10.1007/BF01978099CrossRefGoogle Scholar
Dwivedi, SL, Ceccarelli, S, Blair, MW, Upadhyaya, HD, Are, AK and Ortiz, R (2016) Landrace germplasm for improving yield and abiotic stress adaptation. Trends in Plant Sciences 21, 31–42.CrossRefGoogle ScholarPubMed
Ganie, SA, Wani, BA, Zargar, BA, Sofi, PA and Masoodi, TH (2020) Evaluation of common bean (Phaseolus vulgaris L.) germplasm for resistance to angular leaf spot disease [Phaeoisariopsis griseola (Sacc.) Ferraris] under cold arid conditions of Ladakh. Indian Journal of Agricultural Research 54, 383–387.Google Scholar
Hamid, A, Ahmad, M, Padder, BA, Shah, MD, Sofi, TA and Mohaddin, FA (2016) Distribution of BCMV strains in Kashmir valley and identification of resistant sources of Phaseolus vulgaris. Indian Journal Genetics and Plant Breeding 76, 107–110.CrossRefGoogle Scholar
Haug, W and Lantzsch, HJ (1983) Sensitive method for the rapid determination of phytate in cereals and cereal products. Journal of Science Food and Agriculture 34, 1423–1426.CrossRefGoogle Scholar
Haware, MP and Nene, YL (1982) Races of Fusarium oxysporum f. sp. ciceri. Plant Diseases 66, 809–810.10.1094/PD-66-809CrossRefGoogle Scholar
Ho, MD, Rosas, JC, Brown, KM and Lynch, JP (2005) Root architectural trade-offs for water and phosphorus acquisition. Functional Plant Biology 32, 737–748.CrossRefGoogle Scholar
Horsfall, JG and Barratt, RW (1959) An improved grading system for measuring plant disease. Phytopathology 3, 105–110.Google Scholar
Johnson, HW, Robinson, HF and Comstock, RE (1955) Estimates of genetic and environmental variability in soybeans. Agronomy Journal 47, 314–318.10.2134/agronj1955.00021962004700070009xCrossRefGoogle Scholar
Joshi, B and Mehra, K (1993) Adaptability in French bean (Phaseolus vulgaris L.). Indian Journal of Plant Genetic Resources 6, 73–77.Google Scholar
Joshi, BD and Rana, RS (1995) French Bean in India. New Delhi: National Bureau of Plant Genetic Resources, pp. 1–4.Google Scholar
Kumar, V, Sharma, S, Sharma, AK, Kumar, M, Sharma, S, Malik, S, Singh, KP, Sanger, RS and Bhat, KV (2008) Genetic diversity in Indian common bean (Phaseolus vulgaris L.) using random amplified polymorphic DNA markers. Physiology and Molecular Biology of Plants 14, 383–387.CrossRefGoogle ScholarPubMed
Lhome, A (2005) Creating varieties together. A report submitted to Wagnengin University pp. Vol. 106: 2005.Google Scholar
Lush, JL (1940) Intra-sire correlations or regressions of offspring on dam as a method of estimating heritability of characteristics. Journal of Animal Sciences 1, 293–301.Google Scholar
Narayanan, S, Mohan, A, Gill, KS and Prasad, PV (2014) Variability of root traits in spring wheat germplasm. PLoS One 9, e100317.CrossRefGoogle ScholarPubMed
Ozkan, G, Haliloglu, K, Turkoglu, A, Ozturk, HI, Elkoca, E and Poczai, P (2022) Determining genetic diversity and population structure of common bean (Phaseolus vulgaris L.) landraces from Turkiye using SSR markers. Genes 13, 1410.CrossRefGoogle ScholarPubMed
Panchbhaiya, A, Singh, DK, Verma, P, Jatav, V and Maurya, AK (2017) Genetic analysis of French bean (Phaseolus vulgaris l.) germplasm through principal component analysis and D2 cluster analysis. Journal of Pharmacognosy and Phytochemistry 6, 537–542.Google Scholar
Parveen, S, Yousuf, V, Bhat, F and Bhat, M (2021) Ascochyta blight of common bean: disease status in Kashmir and screening for host plant resistance. Journal of Mycopathological Research 58, 253–257.Google Scholar
Piergiovanni, AR and Laghetti, G (1999) The common bean landraces from Basilicata (Southern Italy): an example of integrated approach applied to genetic resources management. Genetic Resources and Crop Evolution 46, 47–52.CrossRefGoogle Scholar
Rana, JC, Sharma, TR, Tyagi, RK, Chahota, RK, Gautam, NK, Singh, M, Sharma, PN and Ojha, SN (2015) Characterization of 4274 accessions of common bean (Phaseolus vulgaris L.) germplasm conserved in the Indian gene bank for phenological, morphological and agricultural traits. Euphytica 205, 441–57.CrossRefGoogle Scholar
Rawal, V and Navarro, DK (2019) The Global Economy of Pulses. Rome, Italy: Food and Agriculture Organization, p. 190.Google Scholar
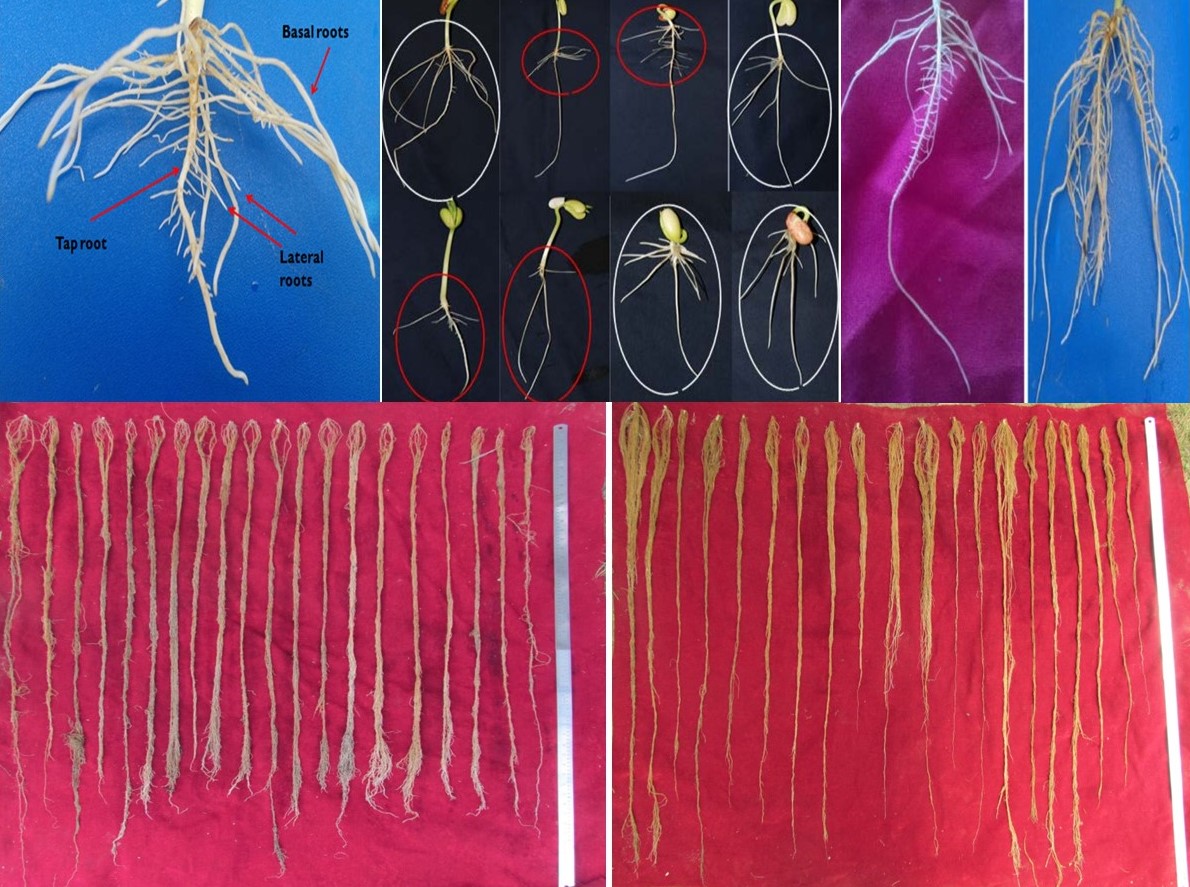
Rehman, K, Sofi, PA, Nida, Y and Bhat, MA (2015) Evaluation of common bean for root traits in relation to drought tolerance. Trends in Biosciences 8, 6859–6865.Google Scholar
Revilla, I and Vivar-Quintana, AM (2008) Effect of canning process on texture of faba beans (Vicia faba). Food Chemistry 106, 310–314.10.1016/j.foodchem.2007.02.046CrossRefGoogle Scholar
Saba, I (2016) Characterization of Rajmash (Phaseolus vulgaris L.) Genotypes for Morphological and Quality traits (PhD thesis). Submitted to SKUAST Kashmir. 232 p.Google Scholar
Saba, I, Sofi, PA, Zeerak, NA, Bhat, MA and Mir, RR (2016) Characterisation of a core set of common bean (Phaseolus vulgaris L.) germplasm for seed quality traits. SABRAO Journal of Breeding and Genetics 48, 359–376.Google Scholar
Schoonhoven, VA and Corrales, MAP (1987) Measurement of damage caused by diseases. In Van Shoonhoven, A (ed.), Standard System for the Evaluation of Bean Germplasm. Cali, Colombia: Centro InternationaI de Agricultura Tropical, pp. 16–44.Google Scholar
Shama, R, Jabeen, N and Sofi, PA (2019) Principal component analysis for assessment of variability in phenological and morphological traits in French bean (Phaseolus vulgaris L). Electronic Journal Plant Breeding 10, 1569–175.CrossRefGoogle Scholar
Singh, SP (2001) Broadening the genetic base of common bean cultivars: a review. Crop Science 41, 1659–1675.10.2135/cropsci2001.1659CrossRefGoogle Scholar
Singh, G, Dukariya, G and Kumar, A (2020) Distribution, importance and diseases of soybean and common bean: a review. Biotechnology Journal International 24, 86–98.10.9734/bji/2020/v24i630125CrossRefGoogle Scholar
Sofi, PA, Rana, JC and Bhat, NA (2014 a) Pattern of variation in common bean (Phaseolus vulgaris L.) genetic resources of Jammu and Kashmir. Journal of Food Legumes 27, 197–201.Google Scholar
Sofi, PA, Zargar, MY, Razvi, SM, Sheikh, FA, Saba, I and Shafi, T (2014 b). Genetic variability and factor analysis in common bean (Phaseolus vulgaris L.) germplasm collection for yield related traits. Electronic Journal Plant Breeding 5, 254–259.Google Scholar
Sofi, PA, Djanaguiraman, M, Siddique, KHM and Prasad, PVV (2018) Reproductive fitness in common bean (Phaseolus vulgaris L.) under drought stress is associated with root length and volume. Indian Journal of Plant Physiology 23, 796–809.CrossRefGoogle Scholar
Sofi, PA, Zargar, SM, Mir, RA and Salgotra, RK (2019) Role of gene banks in maintaining crop genetic resources. In Zargar, SM and Salgotra, RK (eds), Rediscovery of Genetic and Genomic Resources for Future Food Security. Singapore: Springer Nature, pp. 211–224.Google Scholar
Sofi, PA, Gul, M and Ara, A (2020) GY*T biplot approach for genotypic selection in multiple trait evaluations: a case study of cowpea (Vigna unguiculata l.). Agriculture Research Journal 57, 140–147.10.5958/2395-146X.2020.00023.XCrossRefGoogle Scholar
Sofi, PA, Rehman, K, Gull, M, Kumari, J, Djanaguiraman, M and Prasad, PVV (2021) Integrating root architecture and physiological approaches for improving drought tolerance in common bean (Phaseolus vulgaris L.). Plant Physiology Reports 26, 4–22.CrossRefGoogle Scholar
Sofi, PA, Saba, I, Ara, A and Rehman, K (2022). Comparative efficiency of GY* T approach over GT approach in genotypic selection in multiple trait evaluations: case study of common bean (Phaseolus vulgaris) grown under temperate Himalayan conditions. Agricultural Research 11, 373–381.CrossRefGoogle Scholar
Van Schoonhoven, A and Pastor-Corrales, MA (1987) Standard System for the Evaluation of Bean Germplasm. Cali: Inter-American Center of Tax Administrators.Google Scholar
Wani, AB, Bhat, MA, Mir, ZA, Dar, NA and Sofi, PA (2017) Screening of genotypes for identification of resistant genotypes for BCMV. Research Journal of Agricultural Sciences 8, 320–323.Google Scholar
Yan, W and Rajcan, I (2002) Biplot analysis of test sites and trait relations of soybean in Ontario. Crop Science 42, 11–20.CrossRefGoogle ScholarPubMed
Youssef, MM (1978) A Study of Factors Affecting the Cook Ability of Faba Beans (Vicia faba L.) [Doctoral dissertation] (PhD thesis). Egypt: College of Agricultural University of Alexandria.Google Scholar
Zhang, X, Blair, MW and Wang, S (2008) Genetic diversity of Chinese common bean (Phaseolus vulgaris L.) landraces assessed with simple sequence repeat markers. Theoretical and Applied Genetics 117, 629–640.CrossRefGoogle ScholarPubMed

Sofi et al. supplementary material
Sofi et al. supplementary material 1

Image
271.4 KB
Sofi et al. supplementary material
Sofi et al. supplementary material 2
Image
318.6 KB

Sofi et al. supplementary material
Sofi et al. supplementary material 3

File
25.4 KB