


Introduction
The Asiatic wild dog or dhole Cuon alpinus is a primarily pack-living, strictly terrestrial large canid, typically weighing 12–20 kg, usually with a reddish or brown coat and a darker bushy tail (Johnsingh, Reference Johnsingh1985; Durbin et al., Reference Durbin, Venkataraman, Hedges, Duckworth, Sillero-Zubiri, Hoffman and Macdonald2004). It is the only species in the genus Cuon, with 11 subspecies (Durbin et al., Reference Durbin, Venkataraman, Hedges, Duckworth, Sillero-Zubiri, Hoffman and Macdonald2004; Iyengar et al., Reference Iyengar, Babu, Hedges, Venkataraman, MacLean and Morin2005), of which five are reported from south Asia (Johnsingh, Reference Johnsingh1985; Durbin et al., Reference Durbin, Venkataraman, Hedges, Duckworth, Sillero-Zubiri, Hoffman and Macdonald2004). In some areas it was customarily and incorrectly categorized as vermin and assumed to reduce natural populations of wild ungulates and livestock. Organized reward hunting and poisoning exterminated the species over much of its range (Davidar, Reference Davidar and Fox1975). Recently, habitat degradation, extinction of prey populations, and conflicts with and persecution by humans has led to a marked reduction in the dhole's range (Iyengar et al., Reference Iyengar, Babu, Hedges, Venkataraman, MacLean and Morin2005), and it is categorized as Endangered on the IUCN Red List (Durbin et al., Reference Durbin, Hedges, Duckworth, Tyson, Iyengar and Venkataraman2008).
Little is known of the dhole in north-east India and north of the river Ganges, and only a few short-term surveys have been carried out (Johnsingh, Reference Johnsingh1985; Stewart, Reference Stewart1993, Reference Stewart1994). In this area the species has been recorded in the states of Arunachal Pradesh, Assam, Meghalaya, West Bengal (Durbin et al., Reference Durbin, Hedges, Duckworth, Tyson, Iyengar and Venkataraman2008) and Sikkim (Sathyakumar et al., Reference Sathyakumar, Bashir, Bhattacharya and Poudyal2011). Dhole–livestock conflict has been studied in Bhutan (Johnsingh et al., Reference Johnsingh, Yonten and Wangchuck2007), and the species’ ecology is being studied in Pakke Tiger Reserve (Gopi et al., Reference Gopi, Lyngdoh and Selvan2010). The presence of two subspecies of dhole was reported in Sikkim in the 19th century (Gammie, Reference Gammie and Risley1894). Jerdon (Reference Jerdon1874) and Blanford (Reference Blanford1891) mentioned one species of dhole Cuon dukhunensis in Sikkim but that local people believed there were two, differing in colour, size and habits. These were a large type, brownish, with a black muzzle, occurring only in pairs or in groups of three or four, and a small type, reddish in colour, occurring in packs of 10–12 and hunting prey such as wild pig Sus scrofa, barking deer Muntiacus muntjak, goat Capra aegagrus and other livestock. The large type was believed to have medicinal properties and was hunted to meet this demand.
Here we present the first information in c. 100 years on the dhole in the high elevation habitats of Khangchendzonga Biosphere Reserve. Based on camera-trap photographs we present information on the species’ morphology, activity patterns and distribution, and using scat analysis we investigate the dhole's diet. We also highlight the need for further research on the species and for appropriate protection to safeguard this particular subspecies.
Study area
Sikkim, a small mountainous state of India, is the westernmost part of the Eastern Himalayan biodiversity hotspot (Myers et al., Reference Myers, Mittermeier, Mittermeier, da Fonseca and Kent2000). This study was carried out in Khangchendzonga National Park and Biosphere Reserve (Fig. 1) from February 2008 to August 2010. The Reserve encompasses temperate, subalpine and alpine habitats (1,220–5,000 m) and rocky slopes, glacial moraines and permafrost zones (>5,000–8,586 m). There are seven water catchments, of which Prek Chu was selected as the study area as it contains all of the habitat types occurring in the Reserve (Sathyakumar et al., Reference Sathyakumar, Bashir, Bhattacharya and Poudyal2011). The 182 km2 of Prek Chu lies over 1,220 to 6,691 m and receives a total annual rainfall of 1,750–2,250 mm (Tambe, Reference Tambe2007). The major habitats and their percentage of the total area (Fig. 2) are mixed subtropical and mixed temperate (17%), subalpine and krummholtz (36%), alpine pastures (5%), rock and snow cover (41%) and water bodies (1%).

Fig. 1 Location of Khangchendzonga Biosphere Reserve in Sikkim, showing the Khangchendzonga National Park and the catchment of Prek Chu, in which we conducted this study. The shaded area on the inset indicates the location of Sikkim in north-east India.

Fig. 2 The study area, Prek Chu catchment (Fig. 1), showing the locations of camera traps in a 2 × 2 km grid and the locations where dhole Cuon alpinus scats were found.
Methodology
Camera trapping
Prek Chu was divided into 4 km2 blocks, using ArcGIS v. 9.0 (ESRI, Redlands, USA), and categorized into three survey zones according to habitat: temperate (1,200–3,000 m), sub-alpine (3,000–4,000 m) and alpine (> 4,000 m). There were seven, 12 and five blocks in the temperate, subalpine and alpine zones, respectively. The number of camera traps deployed corresponded to the area of the three zones and their accessibility, with a total of 27 camera traps deployed at 71 sites (24, 27 and 20 in the temperate, subalpine and alpine zones, respectively; Fig. 2) for at least 30 days. We used four models of infrared-triggered camera units: two DeerCam (Non Typical, Inc., Park Falls, USA), two Wildview (Stealth Cam, LLC, Grand Prairie, USA), 18 Stealth Cam (Stealth Cam, LLC, Grand Prairie, USA) and five Moultrie (Moultrie Feeders, Alabaster, USA). As we were focusing on a rare species over a large area our strategy was to survey more trap sites less intensively rather than fewer trap sites more intensively (Mackenzie & Royle, Reference Mackenzie and Royle2005). All cameras were set with a 1 minute delay between photographs and 24 h operation, and in four-picture burst mode.
Camera units were attached to trees 15–30 cm above the ground and 3–5 m from a trail or location where animal movement could be expected. Geographical coordinates, elevation and forest type were recorded at each camera-trap location. Camera traps were monitored at least twice per month, when batteries and memory cards were changed.
Sign surveys
Repeated surveys for dhole signs were carried out along 24 trails (with lengths of 1.5–7 km). We walked six trails (a total of 67 trail walks, with a total of 201.15 km of effort) in the temperate zone, 12 trails (108 walks, 299.11 km) in the subalpine zone and six trails (48 walks, 129.17 km) in the alpine zone.
Scat identification, collection and analysis
Identification of dhole scat was straightforward as dholes tend to defecate in the middle of trails, unlike large felids, which defecate along the edges of trails, and because the members of a pack of dholes defecate in the same spot (Kumaraguru et al., Reference Kumaraguru, Saravanamuthu, Brinda and Asokan2011). Dhole scats were found in clusters, exposed on soil (Plate 1a), and could be distinguished easily from the scats of felids, which were stickier and deposited on grass (Johnsingh, Reference Johnsingh1983). We collected only those scats that were deposited in the middle of trails and were either in an aggregation or clustered over 1–1.5 m along a trail. All scats of each aggregation were collected as a group. We excluded all scats found singly, to avoid confusion with scats of red fox Vulpes vulpes in the subalpine zone and jackal Canis aureus in the temperate zone. Scats were collected during the sign surveys. The location, date and other data such as the occurrence of tracks around scats were also recorded.
A reference key was developed for the identification of prey species on the basis of hair structure and morphology. All scat samples were sun-dried in the field and later oven-dried at 60 °C (Sankar & Johnsingh, Reference Sankar and Johnsingh2002) for 48 h and preserved in tagged paper bags for later analysis. Each scat was broken, soaked and washed with running water, using fine-mesh (1.0 and 0.5 mm) filters (Juarez & Filho, Reference Juarez and Filho2002), to separate prey remains such as hair, bones, hooves, teeth, feathers, nails and other undigested remains. Washed samples of hairs from the scats were dehydrated in absolute alcohol for 12 hours and then placed in xylene for 15–20 minutes (Koppikar & Sabnis, Reference Koppikar and Sabnis1976). Hairs were then slide-mounted and examined at 10× and 40× magnification. At least 20 hairs were examined from each scat (Mukherjee et al., Reference Mukherjee, Goyal and Chellam1994) and prey species were identified by comparison with the reference collection, using features such as colour, length, thickness, characteristic medullar configurations (Koppikar & Sabnis, Reference Koppikar and Sabnis1976; Reynolds & Aebischer, Reference Reynolds and Aebischer1991; Mukherjee et al., Reference Mukherjee, Goyal and Chellam1994) and cortex-to-medulla ratio.
Status, distribution and activity
To estimate the relative abundance of the dhole we calculated the photographic and photo-capture rates. The former is the number of camera days required to capture one independent photographic event of a dhole summed across all camera traps in the study (Carbone et al., Reference Carbone, Christie, Conforti, Coulson, Franklin and Ginsberg2001). The latter is the number of independent photographic events of dholes divided by the number of trap-days per site. As the camera traps were set in four-picture burst mode we looked at the revisit rate of the known individuals/groups and found that 54 minutes was the maximum time between revisits in a day. Based on this, we considered any capture > 54 minutes apart as an independent event for calculating photographic and photo-capture rates. Mean photo-capture rates were calculated for all of Prek Chu and for the three survey zones separately (Carbone et al., Reference Carbone, Christie, Conforti, Coulson, Franklin and Ginsberg2001). The number of camera-trap days was calculated from the date of deployment to the date of retrieval (if the memory card was not full) or to the date of the final photograph. Time of day of the photographs was used to determine the daily activity pattern (Pei, Reference Pei1998) of the dhole. A daily activity index was calculated as the number of photographs within a 2-hour duration × 100/total number of photographs. We also calculated the encounter rate of signs per km in each survey zone and used the Mann–Whitney U test to test for differences between zones.
Food habit analysis
Frequency of occurrence (F) of mammalian prey in scats was calculated as F = n/N, where n is the number of dhole scats having that particular species and N is the total number of dhole scats analysed (Karanth & Sunquist, Reference Karanth and Sunquist1995). We subjected the results of the scat analysis to resampling using the bootstrap method, with Simstat (Peladeau, Reference Peladeau2000). Subsamples equalling the original sample size of scats were iterated 10,000 times, to generate means and bias-corrected 95% confidence intervals for percentage frequency of prey items in scats (Mukherjee et al., Reference Mukherjee, Goyal, Johnsingh and Pitman2004). The frequency of occurrence of prey species in the scats was converted to relative biomass (Karanth & Sunquist, Reference Karanth and Sunquist1995) as this provides the best approximation of actual diet (Klare et al., Reference Klare, Kamler and Macdonald2011). We used Y = 0.035+0.020X (Floyd et al. Reference Floyd, Mech and Jordan1978), where Y is kg of prey consumed per scat and X is the mean weight of an individual of a particular prey type (Ackerman et al., Reference Ackerman, Lindzey and Hemker1984). Multiplying each Y by the number of scats found to contain a particular prey species gave the relative weight of each prey type consumed. These values were used to estimate the percentage biomass contribution of each prey species in the dhole's diet (Klare et al., Reference Klare, Kamler and Macdonald2011).
Results
A sampling effort of 6,278 camera-days across 71 sample sites was achieved in the three survey zones (1,407, 3,061 and 1,810 camera-days in the temperate, subalpine and alpine zones, respectively), resulting in 4,517 photographs (2,668 of wild animals and 1,849 of domestic animals and people). We recorded 42 mammal species (Sathyakumar et al. Reference Sathyakumar, Bashir, Bhattacharya and Poudyal2011), of which three were canids (n = 518 photographs): the red fox (n = 456; 152 photo-capture events), dhole (n = 61; nine photo-capture events) and golden jackal (n = 1; one photo-capture event). Of the 61 dhole photographs, obtained at six camera-trap sites, 22 contained more than one individual (Table 1). Based on a wide-angle photograph depicting one stationary animal investigating the camera trap and four resting on the ground, pack size was probably no more than five. Subadult individuals (Plate 1b) were photo-captured twice, once with a pack and once alone (Table 1). Thirty-nine photographs of the dhole were of a single individual (Table 1). All dhole photo-captures were during daytime, with a peak (45% of the captures) at 08.00–10.00.
Table 1 Details of photographs of the dhole Cuon alpinus captured at six camera-trap sites in the Prek Chu watershed of Khangchendzonga Biosphere Reserve (Figs 1–2) during 2009–2010.
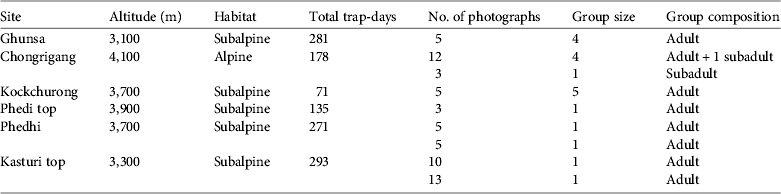
All the adult dholes photographed had a long reddish brown coat and a dark bushy tail, and the pinnae were white inside and reddish brown on the outer side. The muzzle was blackish and relatively short and slightly convex in profile. The nose was black and short white whiskers were present over the white upper and lower lips (Plate 1c). The dorsal and lateral pelage was reddish brown with a yellowish tinge and darker at the neck (Plate 1d). The fore neck, chest and underside were white, with long and dense white fur on the upper chest and throat. Long fur, either white or reddish brown, was present on each paw of adult individuals (Plate 1c,d).

Plate 1 (a) Dhole Cuon alpinus scats found in a cluster in the middle of a trail in temperate forest in the Prek Chu catchment in Khangchendzonga Biosphere Reserve (Figs 1–2). (b) Dhole pack, with one subadult (in the foreground). (c) Adult individual in subalpine–alpine edge forest. (d) Adult in subalpine forest.
The photographic rate (i.e. the minimum number of days required to capture one dhole photographic event) was 541 days. The overall mean photo-capture rate was 0.12 ± SE 0.05 per 100 days and was highest (0.26 ± SE 0.10 per 100 days) in the subalpine zone. Dhole photographs were obtained at five sites (seven photo-capture events) in subalpine forests at elevations of 3,100–3,900 m and at one site (two photo-capture events) at 4,100 m in the alpine zone (Table 1). The sites were dominated by Abies–Betula–Rhododendron forest or dwarf Rhododendron and Juniperus above 3,700 m.
Whilst walking trails we did not encounter dholes and the only signs we found were 41 scats. The encounter rate of scats was significantly higher (P = 0.03, Mann–Whitney U test) in the subalpine (0.21 ± SE 0.1 scats km−1, 31 scats) than in the temperate zone (0.02 ± SE 0.01 scats km−1, 10 scats). We did not detect any evidence of canids in the lower temperate zone (1,850–2,500 m), although we made frequent sightings of jackal below 1,850 m.
Only 29.3% of dhole scats in a group had more than one prey item. Undigested matter was present in the following decreasing order: hair (100%, present in all scats), bone (80.5%), nail (24.4%), teeth (24.4%), hoof (7.3%) and undigested remains of grass (7.3%). Unidentified rodents were the most frequently found prey item, followed by the serow Nemorhaedus sumatraensis, Himalayan tahr Hemitragus jemlahicus, goral Nemorhaedus goral and pika Ochotona sp., unidentified remains and vegetable matter (Table 2). Serow was the largest prey (mean adult body weight 91–100 kg) consumed, followed by the Himalayan tahr (80 kg) and goral (30 kg; Prater, Reference Prater1971). The estimated relative biomass of prey and the relative number of individual prey are also presented in Table 2. The biomass estimates indicate that although dholes killed more pikas and rodents, ungulate prey species (serow, Himalayan tahr and goral) contributed 98.7% of the total biomass consumed.
Table 2 Diet of the dhole in Khangchendzonga Biosphere Reserve (Fig. 1) as indicated by analysis of 41 scats, with the mean percentage frequency of prey items (with 95% confidence intervals, CI, from bootstrapping), estimated % relative biomass of prey, and relative number of individual prey consumed (see text for further details).

Discussion
The reddish-brown long coat and long white or reddish brown hairs on the paws indicate that the dhole population in Khangchendzonga Biosphere Reserve could be the subspecies C. alpinus primaevus (Durbin et al., Reference Durbin, Venkataraman, Hedges, Duckworth, Sillero-Zubiri, Hoffman and Macdonald2004), which has also been reported at lower and middle altitudes in Bhutan (Johnsingh et al., Reference Johnsingh, Yonten and Wangchuck2007). The presence of thick white fur on the chest and throat also distinguishes it from the dhole of southern India C. alpinus dukhunensis, which is known to be genetically distinct from C. alpinus primaevus (Iyengar et al., Reference Iyengar, Babu, Hedges, Venkataraman, MacLean and Morin2005). Iyengar et al. (Reference Iyengar, Babu, Hedges, Venkataraman, MacLean and Morin2005) did not find evidence for the recognition of all 11 subspecies of dhole and showed there is admixture of subspecies over a vast area, and therefore genetic analysis is required to determine the true identity of the dhole of Khangchendzonga Biosphere Reserve.
The photographic rate for the dhole in Khangchendzonga Biosphere Reserve (541 days) lies within the range (29–6,024 days) of that in seven other camera-trap studies in south Asia (Table 3). Trail surveys in Pakke Tiger Reserve in Arunachal Pradesh, India, resulted in a low encounter rate of dhole signs (0.26 km−1; Gopi et al., Reference Gopi, Lyngdoh and Selvan2010), similar to our encounter rate (0.21 km−1) in Khangchendzonga Biosphere Reserve. Comparison of our results with those of other studies is only tentative, however, as there were differences in study design and camera-trapping effort. More robust methods such as occupancy-based abundance estimation or non-invasive DNA-based capture–recapture studies are required to elucidate the status of the dhole in Khangchendzonga Biosphere Reserve and elsewhere.
Table 3 Number of days required to obtain one camera-trap photograph of the dhole (the photographic rate) in Khangchendzonga Biosphere Reserve (Fig. 1) and in seven other protected areas in south Asia (Datta et al., Reference Datta, Anand and Naniwadekar2008).
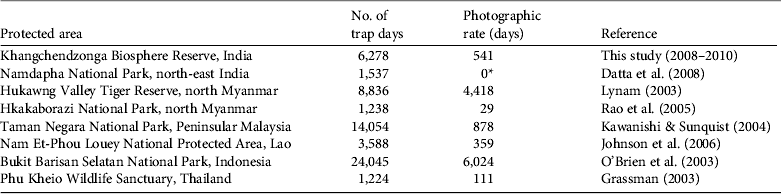
* Datta et al. (Reference Datta, Anand and Naniwadekar2008) did not secure any photographs of the dhole but confirmed its presence through indirect evidence, and Mishra et al., (Reference Mishra, Madhusudan and Datta2006) reported skins of dholes from six valleys at high altitudes in Arunachal Pradesh.
Factors that may influence habitat selection by dholes include the availability of medium to large ungulate prey species, water, the presence of other large carnivore species, human population density and suitability of breeding sites (Durbin et al., Reference Durbin, Venkataraman, Hedges, Duckworth, Sillero-Zubiri, Hoffman and Macdonald2004). In the subalpine forests, serow and goral were relatively abundant and Himalayan tahr was also present (Bhattacharya et al., Reference Bhattacharya, Bashir, Poudyal, Sathyakumar, Bisht and Saha2010). No other large or medium-sized carnivore species has been reported from these subalpine forests except the Asiatic golden cat Pardofelis temminckii (Bashir et al., Reference Bashir, Bhattacharya, Poudyal and Sathyakumar2011), which is reported to be sympatric with the dhole in Peninsular Malaysia (Kawanishi & Sunquist, Reference Kawanishi and Sunquist2008). Presence of large and medium ungulate prey, absence of large carnivores such as the common leopard Panthera pardus and relatively less anthropogenic pressure may be the reasons for the presence of the dhole primarily in subalpine forests in our study area.
Other studies have also indicated that the dhole is diurnal and obtains most of its dietary biomass from ungulate prey (Johnsingh, Reference Johnsingh1992; Karanth & Sunquist, Reference Karanth and Sunquist1995; Venkataraman et al., Reference Venkataraman, Arumugum and Sukumar1995; Kawanishi & Sunquist, Reference Kawanishi and Sunquist2008; Borah et al., Reference Borah, Deka, Dookia and Gupta2009; Kumaraguru et al., Reference Kumaraguru, Saravanamuthu, Brinda and Asokan2011; Kamler et al., Reference Kamler, Johnson, Vongkhamheng and Bousa2012). The presence of pikas and rodent species in the diet of the dhole could be because of its ability to flush out and hunt smaller and cryptic prey species (Venkataraman, Reference Venkataraman1996; Kumaraguru et al., Reference Kumaraguru, Saravanamuthu, Brinda and Asokan2011). Although too small for the pack as a whole, such small prey is sufficient for an individual dhole (Kumaraguru et al., Reference Kumaraguru, Saravanamuthu, Brinda and Asokan2011). Dholes also occasionally consume grass and other vegetation (Cohen et al., Reference Cohen, Fox, Johnsingh and Barnett1978; Johnsingh, Reference Johnsingh1983; Durbin et al., Reference Durbin, Venkataraman, Hedges, Duckworth, Sillero-Zubiri, Hoffman and Macdonald2004), which explains the presence of undigested vegetation remains in some scats.
Our results indicate that probably only one subspecies of dhole, C. alpinus primaevus, is present in the subalpine forests of Prek Chu. Iyengar et al., (Reference Iyengar, Babu, Hedges, Venkataraman, MacLean and Morin2005) suggested that this subspecies, reported to be ‘very rare’ in a survey in the early 1980s (Johnsingh, Reference Johnsingh1985), and distinct from C. alpinus dukhunensis, should be accorded a high priority for conservation action. Other catchments in Khangchendzonga Biosphere Reserve need to be surveyed, with intensive camera trapping, for the dhole. In Bhutan dholes have caused livestock loss because villagers allowed their livestock to graze unsupervised (Johnsingh et al., Reference Johnsingh, Yonten and Wangchuck2007). We recommend that livestock depredation cases should be monitored in Khangchendzonga Biosphere Reserve as unsupervised grazing is also practised in this area. Our findings have been shared with the relevant State and Central authorities, for enhancement of the conservation management of the dhole in this region, and a further project has now been initiated to monitor the dhole population in Prek Chu and other catchments in Khangchendzonga Biosphere Reserve.
Acknowledgements
We are grateful to the Department of Forests, Environment and Wildlife Management, Government of Sikkim, for granting us permission to work in the state. We thank the Wildlife Institute of India, Dehradun, for providing grants and support, and two anonymous reviewers for their valuable comments.
Biographical sketches
Tawqir Bashir is interested in the ecology and conservation of carnivores in mountain ecosystems, with special reference to predator–prey interactions and reducing carnivore–human conflicts. Tapajit Bhattacharya's interests are the ecology and conservation of mountain ungulates, including their habitats and ungulate–habitat interactions, and community-based conservation. Kamal Poudyal's interests are the ecology and conservation of the wildlife of the Eastern Himalaya and community participation in conservation. Manjari Roy's interests are the ecology and conservation of large carnivores, and mangrove ecosystems. Sambandam Sathyakumar is interested in the ecology and conservation of the wildlife of the Himalaya, where he has been following research interests in mountain ungulates, bears, other large carnivores and Galliformes. He is a member of the IUCN/Species Survival Commission Caprinae, Bear, Galliformes, and Cultural and Spiritual Values of Protected Areas Specialist Groups.






