


Introduction
Oceanic islands have captured the attention of conservation biologists because of the high species endemism and vulnerability to a variety of threats (Courchamp et al., Reference Courchamp, Hoffmann, Russell, Leclerc and Bellard2014). Mexico is an important region for biodiversity conservation, given its high number of species, many of which are endemic (Martínez-Meyer et al., Reference Martínez-Meyer, Sosa-Escalante and Álvarez2014). A high proportion of Mexican mammal species are endemic (32%), of which 20% are found only on islands (Ceballos & Rodríguez, Reference Ceballos, Rodríguez, Medellín and Ceballos1993). Endemic insular mammal species are particularly vulnerable; for example, 80% of extinctions of endemic Mexican mammals have occurred on islands (Ceballos & Navarro, Reference Ceballos, Navarro, Mares and Schmidly1991; Mellink, Reference Mellink and Tell2000; Tershy et al., Reference Tershy, Donlan, Keitt, Croll, Sanchez, Wood, Veitch and Clout2002).
A prominent example of an insular threatened Mexican mammal is the fish-eating bat Myotis vivesi (Vespertillionidae; Herrera & Flores-Martínez, Reference Herrera and Flores-Martínez2001). The bat is endemic to islands of the Gulf of California, with a few reports of its presence on the Gulf and Pacific coastlines (Blood & Clark, Reference Blood and Clark1998), and is categorized as Vulnerable on the IUCN Red List (Arroyo-Cabrales & Ospina-Garces, Reference Arroyo-Cabrales and Ospina-Garces2016) and Endangered, with risk of extinction, by Mexican authorities (SEMARNAT, 2010). The fish-eating bat generally roosts within rock crevices on talus slopes, making it easy prey for introduced domestic cats Felis catus and black Rattus rattus and brown rats Rattus norvegicus. The bat forms maternity colonies during spring–summer (Flores-Martínez et al., Reference Flores-Martínez, Floyd, Herrera, May, Medellín and Sánchez-Cordero2005) and, unlike other bat species, it feeds mainly on marine fish and crustaceans (Otálora-Ardila et al., Reference Otálora-Ardila, Herrera M., Flores-Martínez and Voigt2013).
The fish-eating bat has been reported historically from almost 50 localities (McLellan, Reference McLellan1927; Miller & Allen, Reference Miller and Allen1928; Burt, Reference Burt1932; Walker, Reference Walker1950; Reeder & Norris, Reference Reeder and Norris1954; Cockrum & Bradshaw, Reference Cockrum and Bradshaw1963; Maya, Reference Maya1968; Patten & Findley, Reference Patten and Findley1970; Villa, Reference Villa1979; Blood & Clark, Reference Blood and Clark1998) but there is no recent account of its distribution. Here we present the results of several expeditions conducted to verify the current geographical distribution of the fish-eating bat. We identified the localities in which maternity colonies are formed, estimated the population size of the Partida Norte colony in 2003, and monitored the presence of bats on Partida Norte Island during 2004–2016. We also recorded the presence of introduced domestic cats and rats (Villa, Reference Villa1979). Based on the information derived in our surveys along with published information on the biology of the fish-eating bat, we discuss the adequacy of its conservation status on the IUCN Red List and that established by Mexican authorities.
Methods
We visited 56 sites (48 islands and eight mainland sites) in the Gulf of California (181,000 km2; Case et al., Reference Case, Cody and Ezcurra2002) in north-west Mexico during expeditions in 2001–2003, 2005–2007, 2009, 2011 and 2014–2016 (Supplementary Table S1). Sites were selected based on historical records of the presence of fish-eating bats, but we also included sites where their presence was not reported previously. Most sites visited were islands; only eight were on the mainland. We opened 2–3 mist nets (12 × 2 m) during 1–2 nights at each site visited; nets were open from sunset to 06.00 and they were checked every 15–20 minutes. Captured bats were sexed and their reproductive condition determined based on external morphological evidence. To determine the presence of introduced domestic cats and rodents on each island, we set 3–5 Tomahawk (Hazelhurst, USA) and 80–100 Sherman (Tallahassee, USA) traps during 1–2 nights; we also surveyed the area for the presence of cat scats. Some islands were surveyed for bats and introduced fauna on more than one occasion. We used ArcGIS 10.5 (ESRI, Redlands, USA) to estimate the extent of occurrence (‘the area contained within the shortest continuous imaginary boundary which can be drawn to encompass all the known, inferred or projected sites of present occurrence of a taxon’; IUCN, 2012) based on the sites where we found the bat. We estimated the area of each island surveyed using ArcGIS 10.5.
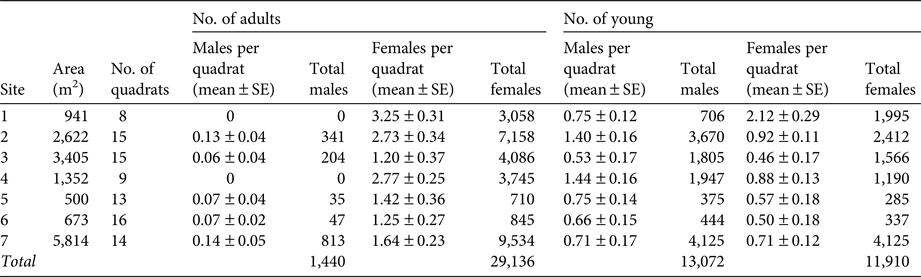
We estimated the size of the adult population in the bat colony on Partida Norte Island in May 2003 using the method used by Flores-Martínez et al. (Reference Flores-Martínez, Floyd, Herrera, May, Medellín and Sánchez-Cordero2005). The population size of this colony had been estimated in May 2001 using quadrats located at two sites used as roosts by bats (sites 5 and 6 in Flores-Martínez et al., Reference Flores-Martínez, Floyd, Herrera, May, Medellín and Sánchez-Cordero2005). Fish-eating bats also roost on cliffs but surveying these areas was not possible. We recorded presence of fish-eating bats in 8–16 quadrats (1 × 1 m) placed randomly in seven roosting areas (Table 1). We measured the area covered by each roosting site using a metric tape and extrapolated the number of bats present using the mean number of bats per quadrat at each site. We considered the total adult population size to be the sum of the number of bats estimated for each roosting area. We recorded the presence of young and estimated their total number similarly. We conducted our sampling in the southern portion of the island, including sites 3, 5 and 6 from Flores-Martínez et al. (Reference Flores-Martínez, Floyd, Herrera, May, Medellín and Sánchez-Cordero2005) and four additional sites. We did not sample the northern part of the island (sites 1, 2 and 4 from Flores-Martínez et al., Reference Flores-Martínez, Floyd, Herrera, May, Medellín and Sánchez-Cordero2005). We also monitored the presence of the bat on Partida Norte Island in 1–5 periods during 2004–2006 and 2008–2016. We did not attempt to repeat the extensive surveying conducted in 2001 and 2003, to avoid disturbing the roosts. Instead, we conducted low-intensity, non-systematic sampling as part of other projects to study the bat's ecology. We extracted fish-eating bats from 4–10 quadrats (1 × 1 m) in each sampling period, and returned them to their roosts following processing. Therefore, our sampling protocol did not use a standard methodology to produce estimates of population size or to establish patterns in sex ratio; rather it was conducted opportunistically while monitoring the long-term temporal presence of fish-eating bats on the island.
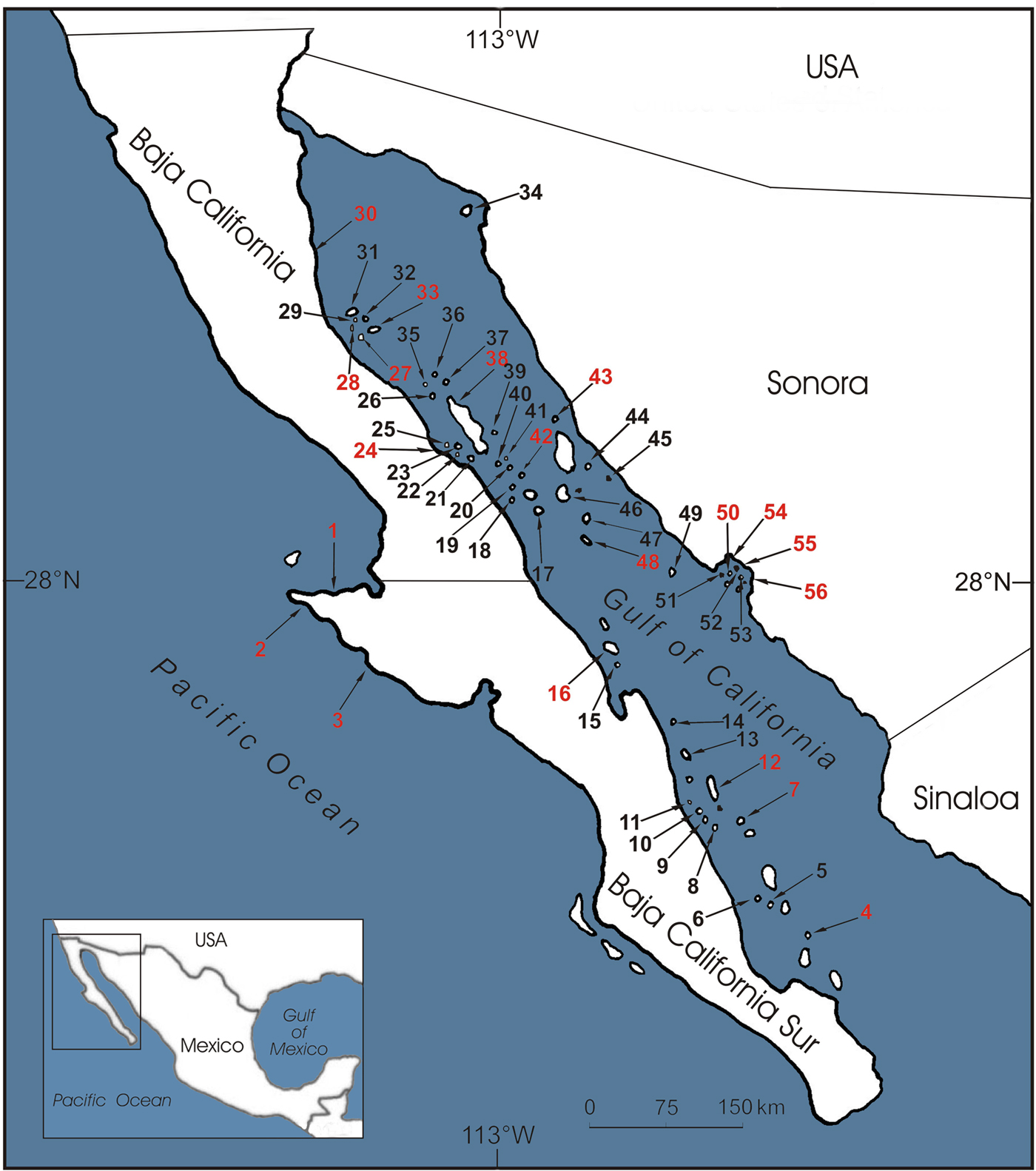
Fig. 1 Sites in the Gulf of California surveyed for the presence of the fish-eating bat Myotis vivesi during 2001–2016 (see Supplementary Table S1 for details). Numbers in black represent sites where the bat was found; numbers in red represent sites where the bat was absent.
Table 1 Population estimates for the fish-eating bat Myotis vivesi at seven roosting sites at the southern end of Partida Norte Island in May 2003 (site 41, Fig. 1), with the site area, number of 1 × 1 m quadrats, and numbers of adult and young males and females.
Results
We captured or observed fish-eating bats on 36 islands, 17 of which were new reports (Supplementary Table S1). We recorded absence of the bats at 20 sites, of which 13 had been reported previously (Supplementary Table S1). Maternity colonies of the fish-eating bat were found on 19 islands (Supplementary Table S1). We captured black rats on five islands where the bat was present and on one island where the bat was absent; we captured mice Mus musculus on one island where the bat was present, and black rats and mice on one island with and one island without bats (Supplementary Table S1). We captured no cats but we found their scats on one island where the bat was present and on three islands where the bat was absent (Supplementary Table S1). The extent of occurrence estimated for the bat was 54,665 km2. The sum of the areas of the islands on which fish-eating bats were found was 93.12 km2, and the sum of the areas of the islands on which the bat was absent was 283.30 km2 (Supplementary Table S1).
We estimated a population size of 30,576 adult fish-eating bats in May 2003 on Partida Norte Island, of which c. 95% were females (Table 1). We estimated the presence of 24,982 young, with a slightly higher proportion of males (52.3%) than females (47.6%; Table 1). We captured fish-eating bats in all 13 years in which we visited Partida Norte Island (Supplementary Table S1; Table 1).
Discussion
Given the high levels of endemism on its > 250 islands, the Gulf of California is a biodiversity hotspot for conservation (Case et al., Reference Case, Cody and Ezcurra2002; Tershy et al., Reference Tershy, Donlan, Keitt, Croll, Sanchez, Wood, Veitch and Clout2002). The uniqueness of the biology of the fish-eating bat makes it a particularly threatened species. Our expeditions found that this bat is widespread in the Gulf of California, although we failed to find evidence of its presence at some sites where it was recorded historically. At some sites (San Carlos Bay and San Sebastián Vizcaíno) the absence is probably temporary; for example, records from the mainland probably reflect temporary roosting when foraging (Maya, Reference Maya1968). Similarly, some of the islands where we did not observe fish-eating bats are probably used occasionally at night; for example, there is a historical record of the presence of fresh guano on Carmen (Maya, Reference Maya1968), and it seems the bats use Rasa to perch while foraging, as we found fresh guano there during our visit. In contrast to most historical accounts of the bat's distribution, we are certain that the sites where we recorded presence of the fish-eating bat represent established colonies on islands used as daytime roosts (but see below for Mejía Island). The absence of fish-eating bats on some islands may simply indicate that their use of islands is temporary, and that some islands are abandoned when new sites are colonized; for example, although Partida Norte and Cardonosa Islands are occupied by an equal number of males and females during autumn and winter, most adult males abandon these islands during spring–summer, with the remaining bats comprising mostly pregnant and lactating females with young (Maya, Reference Maya1968; Flores-Martínez et al., Reference Flores-Martínez, Floyd, Herrera, May, Medellín and Sánchez-Cordero2005). It is possible that males use different islands at different times of the year, and therefore the timing of surveys is critical for their detection at a particular site. In most cases we are not certain if the sites where we found maternity colonies have similar presence–absence temporality to Partida Norte and Cardonosa Islands, as they were visited mostly during spring–summer.
Most islands where fish-eating bats were observed seemed to offer adequate conditions for their survival, as no introduced domestic cats or rats were found. However, these introduced species are predators of the fish-eating bat (Villa, Reference Villa1979; Vázquez-Domínguez et al., Reference Vázquez-Domínguez, Ceballos and Cruzado2004) and are probably one of the main threats to its survival (Ceballos & Navarro, Reference Ceballos, Navarro, Mares and Schmidly1991). Invasive mammals are responsible for most extinctions and extirpations in north-west Mexico (Tershy et al., Reference Tershy, Donlan, Keitt, Croll, Sanchez, Wood, Veitch and Clout2002) and we found cats and rats on several islands where fish-eating bats were present. Cats have been eradicated from Mejía (1999–2001), Danzante (2000) and Estanque Islands (1999), and rats have been eradicated from Rasa (1995–1996), San Pedro Mártir (2007) and San Jorge Islands (2000–2002) (Ceballos & Ramírez, Reference Ceballos and Ramírez1995; Aguirre-Muñoz et al., Reference Aguirre-Muñoz, Samaniego-Herrera, García-Gutiérrez, Luna-Mendoza, Rodríguez-Malagón, Casillas-Figueroa, Sánchez, Peters, Márquez-Huitzil, Vega, Portales, Valdez and Azuara2005, Reference Aguirre-Muñoz, Croll, Donlan, Henry, Hermosillo and Howald2008). We found fish-eating bats on Danzante, Estanque and San Jorge Islands after these predators had been eradicated. We found both fish-eating bats and rats on Mejía Island in 2001 and 2006, but we found only rats in 2013 and 2014. It is probable that the bats use the island transiently while foraging, and that their permanent presence is prevented by the presence of rats. Rat eradication actions were conducted on Mejía in 2013 (J.J. Flores-Martínez, pers. comm.) but eradication was not confirmed afterwards. In our surveys in 2015 and 2016 we captured no rats on Mejía but nor did we capture any bats. We did not observe fish-eating bats on San Pedro Mártir (2009) or Rasa Islands after eradications, although these islands are probably used as nocturnal perches.
The fish-eating bat is categorized as Vulnerable on the IUCN Red List (Arroyo-Cabrales & Ospina-Garces, Reference Arroyo-Cabrales and Ospina-Garces2016) based on a > 30% population decline over the last three generations, an area of occurrence of 130,000 km2 with a severely fragmented distribution, a declining area of occupancy (‘the area within its extent of occurrence which is occupied by a taxon’; IUCN, 2012), and a low rate of recolonization of restored habitat because of a strong female philopatry. We consider this categorization to be appropriate but we disagree in part with some of the reasoning. There is no published evidence that the population size (‘total number of individuals of the taxon’; IUCN, 2012) has decreased by > 30%. The only colony in which the population size has been estimated is on Partida Norte Island, where there were estimated to be 10,000–12,000 adults in June–July 1963 (Maya, Reference Maya1968), c. 8,000 adults in May 2001 (Flores-Martínez et al., Reference Flores-Martínez, Floyd, Herrera, May, Medellín and Sánchez-Cordero2005), and 30,576 adults in May 2003 (this study). These estimates indicate that the size of the colony did not decrease in the 40-year period in which the surveys were conducted. Additionally, we have recorded a continuous presence of fish-eating bats on Partida Norte Island since 2003, with no obvious declining trend. Our estimate of the area of occurrence (54,665 km2) is less than half that estimated by IUCN, which is partly explained by the fact that we did not record the presence of bats at mainland sites, which are probably used only as resting sites. Although the area of occurrence of fish-eating bats is highly fragmented, with daytime roost sites separated by large expanses of open water, nuclear DNA analysis has indicated high rates of gene flow among colonies (Floyd et al., Reference Floyd, Flores-Martínez, Herrera M., Mejía and May2010). The area of occupancy of the species has probably declined over time, as indicated by the IUCN assessment and the fact that we did not find bats in some locations where they had been recorded previously. The area of occupancy of fish-eating bats includes both terrestrial and oceanic habitats, which play different roles and have different properties. The oceanic part of the area of occupancy is used for foraging, and preliminary global positioning system tracking of bats from Partida Norte Island revealed that foraging is consistently restricted to an area of c. 1,300 km2 (E. Hurme, Y. Yovel & L. G. Herrera M., unpubl. data). The terrestrial component of the area of occupancy is represented by the suitable portion of the islands used as roosting habitat. The total area of all islands where we found bat roosts was 93.12 km2 but the sum of the actual areas used as roost by bats on each island may be smaller; for example, on Partida Norte Island areas with rock slides that are suitable for establishing roosting sites comprise < 1% of the total surface of the island. Therefore, the area of occupancy for the oceanic and terrestrial habitats may be only a small portion of the habitat available for the fish-eating bat within its extent of occurrence. We agree that strong female philopatry could reduce the probability of fish-eating bats recolonizing restored habitats (Floyd et al., Reference Floyd, Flores-Martínez, Herrera M., Mejía and May2010), although we know of no attempts to evaluate this.
The fish-eating bat is listed as an Endangered species by Mexican conservation authorities, based on its geographical distribution in Mexico, the quality of the habitat in relation to the requirements of the species, its intrinsic biological vulnerability, and the impact of human activity (Sánchez et al., Reference Sánchez, Medellín, Aldama, Goettsch, Soberón and Tambutti2007). We consider this categorization to be appropriate, as the species’ extent of occurrence is < 5% of Mexico's total land area. The species is intrinsically vulnerable because its roosting habits expose it to a large range of terrestrial predators and it aggregates in large numbers (thousands of individuals) in relatively small areas. The bat is susceptible to the effects of human activities because the islands are used regularly by local fishers to camp, and to the presence of cats and rats (which are also vectors of emerging diseases).
The Gulf of California is exposed to the effects of interannual climate events, industrialized, sport and artisanal fisheries, and coastal zone usage that can have profound effects on biota (Lluch-Cota et al., Reference Lluch-Cota, Aragón-Noriega, Arreguín-Sánchez, Aurioles-Gamboa, Bautista-Romero and Brusca2007). Several of the islands on which we recorded fish-eating bats are considered to be of high priority for biodiversity conservation in this area (WWF, 2006). Management strategies for the conservation of this species are similar to currently established policies for other threatened vertebrates inhabiting these islands; for example, monitoring of the presence of introduced vertebrates, and implementation of eradication programmes where necessary; restriction of human activities on the islands; monitoring of the population size of selected colonies (especially maternity colonies); periodic surveys of the geographical distribution of the species; examination of bat tissues for the presence of anthropogenic contaminants (e.g. heavy metals; Méndez & Alvarez-Castañeda, Reference Méndez and Alvarez-Castañeda2000); and assessment of the effects of fisheries on food resource availability. Given its feeding habits, the conservation of the fish-eating bat is relevant for the nutritional and energy budget of terrestrial ecosystems where it roosts, because it transports marine-derived nutrients and energy (Otálora-Ardila et al., Reference Otálora-Ardila, Herrera M., Flores-Martínez and Voigt2013). Protecting the species involves the conservation of a unique adaptation to marine environments not present in any other bat species.
Acknowledgements
This study was funded by grants to LGHM and JJFM from Bat Conservation International and Programa para la Conservación de los Murciélagos de México, by a grant to JJFM from Consejo Nacional de Ciencia y Tecnología (#203671), and by grants to LGHM from Dirección de Asuntos del Personal Académico (IN201108, IN202113), Fondo Sectorial de Investigación Ambiental (#C01-15), Comisión Nacional Para el Uso y Conocimiento de la Biodiversidad (#HK002), and Consejo Nacional de Ciencia y Tecnología (#237774). The study was conducted with permits from Secretaría de Medio Ambiente y Recursos Naturales. Ulalume Hernández A., Valeria B. Salinas R., Aída Otálora A. and Marcia Carmona M. participated in the projects from which the long-term monitoring of the bat colony on Partida Norte Island was derived. Transportation was generously provided by Secretaría de Marina-Armada de México. Alfredo Zavala, Carlos Godínez, Juan Pablo Gallo, Tad Pfister, Lorayne Meltzer, the Prescott College Kino Bay Center and personnel from the Área de Protección de Flora y Fauna Islas del Golfo de California-Baja California provided invaluable logistic support. We thank Leonel Moreno M. and Vaporub for making the expeditions safer and more enjoyable.
Author contributions
LGHM and JJFM designed the study and collected data. LGHM, JJFM and VSC wrote the article.
Biographical sketches
L. Gerardo Herrera and J. Juan Flores-Martínez have led several projects on the biology and conservation of the fish-eating bat during the last 16 years. Herrera conducts research in bat ecology, physiology and conservation. Flores-Martínez is carrying out research on mammal biology, and supervises projects for the eradication of introduced fauna. Víctor Sánchez-Cordero conducts research in natural protected areas.



