


Introduction
Populations of large carnivores are increasingly threatened (Woodroffe, Reference Woodroffe2000), with loss of habitat (Hanski, Reference Hanski2005), depleted prey bases (Woodroffe, Reference Woodroffe2000), and persecution by livestock farmers (Woodroffe, Reference Woodroffe2000; Treves & Karanth, Reference Treves and Karanth2003; Woodroffe & Frank, Reference Woodroffe and Frank2005; Holmern et al., Reference Holmern, Nyahongo and Røskaft2007). Most viable populations of large carnivores in Africa are now restricted to protected areas (Bauer & Van Der Merwe, Reference Bauer and Van Der Merwe2004; Hayward & Kerley, Reference Hayward and Kerley2009). Populations of African lions Panthera leo have been particularly affected, with an estimated 32,000–35,000 free-ranging lions remaining in Africa, and protected areas are necessary to ensure their persistence (IUCN, 2013; Riggio et al., Reference Riggio, Jacobson, Dollar, Bauer, Becker and Dickman2013). However, populations in protected areas are still exposed to legal and illegal hunting, road kill, poisoning and snaring (Woodroffe & Ginsberg, Reference Woodroffe and Ginsberg1998; Woodroffe, Reference Woodroffe2000), which compromise their viability and likelihood of survival.
Growth and stability of lion populations are largely determined by social and ecological factors such as density, tenure of coalitions of residential male lions, the number of females in a pride, intergroup competition, group territoriality, and prey availability (Van Orsdol, Reference Van Orsdol1984; Packer et al., Reference Packer, Scheel and Pusey1990, Reference Packer, Hilborn, Mosser, Kissui, Borner and Hopcroft2005; Mosser & Packer, Reference Mosser and Packer2009). Frequent removal of resident males may influence successful reproduction, either through factors such as infanticide or through reproductive suppression until social stability within the pride is restored (Smuts, Reference Smuts1982; Whitman & Packer, Reference Whitman and Packer1997). The removal of adult females, the main breeding units of a pride, can also influence reproduction and population growth as larger groups have higher rates of reproductive success and tend to gain access to the best-quality habitats (Packer et al., Reference Packer, Herbst, Pusey, Bygott, Hanby, Cairns, Borgerhoff Mulder and Clutton-Brock1988; Loveridge et al., Reference Loveridge, Searle, Murindagomo and Macdonald2007; Mosser & Packer, Reference Mosser and Packer2009). The loss of cubs and subadults affects population size and future reproductive potential.
Many lion populations are restricted to small reserves that are geographically isolated from each other (Bauer & Van Der Merwe, Reference Bauer and Van Der Merwe2004; Slotow & Hunter, Reference Slotow, Hunter, Hayward and Somers2009). This introduces further difficulties for conservation managers, as small populations are slower to recover from stochastic events and genetic diversity is often limited (Hayward et al., Reference Hayward, O'Brien and Kerley2007). In southern Africa in particular many small populations have been fenced in an attempt to allow lions to persist in a human-dominated landscape (Hayward & Kerley, Reference Hayward and Kerley2009; Packer et al., Reference Packer, Loveridge, Canney, Caro, Garnett and Pfeifer2013). In the absence of anthropogenic threats lion populations have the potential to increase rapidly (Kettles & Slotow, Reference Kettles and Slotow2009), and therefore managers must intervene to limit population growth in small, isolated reserves (Slotow & Hunter, Reference Slotow, Hunter, Hayward and Somers2009). In unfenced reserves animals may attempt to disperse as population density increases but these animals are usually killed when surrounding land uses are incompatible with large predators (Woodroffe & Frank, Reference Woodroffe and Frank2005).
Population control methods in fenced reserves may include culling, contraception, translocation and male vasectomies (Kettles & Slotow, Reference Kettles and Slotow2009; Slotow & Hunter, Reference Slotow, Hunter, Hayward and Somers2009). As an alternative, trophy hunting is sometimes used as a management tool, with the added benefit of generating revenue for protected areas. Lions are highly sought after by trophy hunters and hunting is permitted in several African countries. Although this form of harvesting is typically conducted under the guise of sustainable utilization, the selective removal of individuals has been reported to have several negative effects, including population decline (Loveridge et al., Reference Loveridge, Searle, Murindagomo and Macdonald2007), distorted social organization (Yamazaki, Reference Yamazaki1996) and distorted sex ratios (Creel & Creel, Reference Creel and Creel1997; Loveridge et al., Reference Loveridge, Searle, Murindagomo and Macdonald2007). Perturbations are more evident in smaller and geographically isolated prides, which are more vulnerable to takeovers, resulting in lower reproductive success and increased vulnerability to kleptoparasitism (Packer et al., Reference Packer, Herbst, Pusey, Bygott, Hanby, Cairns, Borgerhoff Mulder and Clutton-Brock1988). The preference of trophy hunters for large adult males may negatively affect population dynamics (Whitman et al., Reference Whitman, Starfield, Quadling and Packer2004).
In contrast to trophy hunting, illegal killing of lions in and around unfenced reserves is predominantly indiscriminate, such as when individuals are caught in snares or exterminated by livestock owners (Woodroffe & Frank, Reference Woodroffe and Frank2005). The effects on social organization, growth rate and survival may therefore differ from those arising from selective harvesting.
It is important for conservation managers to understand how lion population dynamics respond to these various forms of anthropogenic mortality. Here we report our findings for two small populations of lions occurring only 10 km apart, one exposed to legal hunting as a form of population control (in a fenced reserve) and the other to illegal persecution (in a largely unfenced reserve). Taking prey availability into account we assess the mean pride size, number of adult females per pride, cub survival rate, and population growth rate, size and density in the two reserves. We then consider the relative influence of different types of human-induced mortalities on each population.
Study area
The Northern Tuli Game Reserve and the Venetia Limpopo Nature Reserve are two privately owned reserves located c. 10 km apart in southern Africa (Fig. 1). The Northern Tuli Game Reserve (720 km2) is in south-east Botswana, bordering Zimbabwe and South Africa. The western boundary of the Reserve is game-fenced and the southern boundary, bordering South Africa, has a poorly maintained fence (total fence length c. 86 km). These fences do not represent a significant barrier to the movement of large predators, and wild dogs Lycaon pictus, cheetahs Acinonyx jubatus and lions having been recorded traversing the fences (Jackson et al., Reference Jackson, McNutt and Apps2012). The south-western, northern and eastern boundaries (c. 71 km) are unfenced. The movement of carnivores between the Reserve and surrounding areas is thus largely unrestricted. No form of lion population control or management was implemented in the Reserve during the study period.
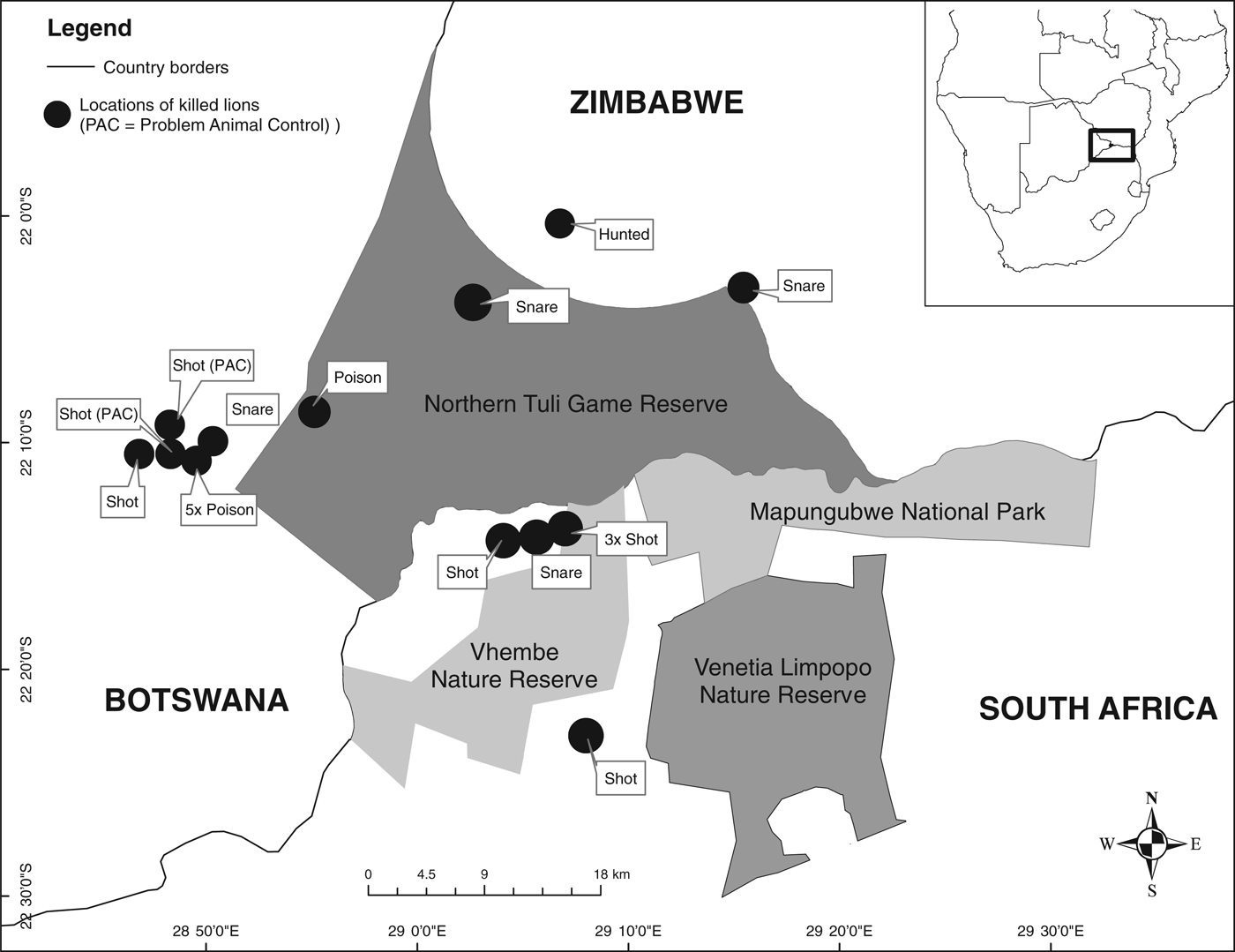
Fig. 1 The Northern Tuli Game Reserve in Botswana and the Venetia Limpopo Nature Reserve in South Africa. The locations where lions Panthera leo from the unfenced Northern Tuli Game Reserve were killed during 2000–2011 are marked on the map.
The Venetia Limpopo Nature Reserve (330 km2) is situated in the far north of South Africa, close to the border with Botswana and Zimbabwe. It is entirely fenced with high-quality electrified predator-proof fencing, which is inspected daily. Safari hunting and euthanasia were the only forms of lion population control implemented here during the study. No introductions were ever made into the lion population in the Reserve.
These two reserves are scheduled to become part of the Greater Mapungubwe Transfrontier Conservation Area, which will also incorporate other reserves in the area.
Methods
Historical records indicate that lions have been present in the Northern Tuli Game Reserve for many decades, and according to early reports the lion population has been persecuted, with at least 150 lions killed during the 1950s (Lind, Reference Lind1974). By the late 1960s lions were almost extirpated from the Reserve (McKenzie, Reference McKenzie1990) but the population began to recover in the early 1970s (Lind, Reference Lind1974). Since 1993 game rangers have completed a sightings datasheet after every game drive, providing a detailed long-term dataset. We analysed 15,067 records collected in the Northern Tuli Game Reserve during September 2000–December 2011.
The Venetia Limpopo Nature Reserve comprises former livestock farms and was designated a nature reserve in the early 1990s. It was recolonized by seven lions from surrounding areas (Cotterill, Reference Cotterill1996), and by 1994 reserve managers estimated that there were 12–15 resident lions. By 1996 the number had increased to 22 (Cotterill, Reference Cotterill1996). Since then researchers and Reserve management staff have monitored the population closely. Trophy hunting has been carried out since 2000 to regulate population growth and reduce the number of lions over time. Initially lion hunting was indiscriminate and 14 lions (10 males and four females) were shot during 2000–2003. In 2003 a continuous and detailed monitoring programme was introduced by Funston & Janse Van Rensburg (Reference Funston and Janse Van Rensburg2003), after which only specific individuals were removed from the population. Safari hunting to manage prey populations within the fenced reserve was also common practice.
We use monitoring data for 2000–2011 to compare and contrast demographic parameters of the lion populations within each reserve. Although carnivore density may vary significantly between different ecosystems, it is correlated with the biomass of preferred prey species present (Bertram, Reference Bertram1975; Hayward et al., Reference Hayward, O'Brien and Kerley2007; Hayward & Kerley, Reference Hayward and Kerley2008). To control for potential population-level effects arising from variations in prey availability we use prey census data to calculate the available biomass of the lion's preferred prey species according to Hayward et al. (Reference Hayward, O'Brien and Kerley2007). Preferred prey species include zebra Equus burchelli, wildebeest Connochaetes taurinus, giraffe Giraffa camelopardalis, gemsbok Oryx gazella and eland Taurotragus oryx. Eland are functionally similar to buffalo Syncerus caffer and serve as a substitute for buffalo in the lion's diet, given that buffalo are not present in either of the study areas.
Results
During 2005–2011 there were 19 reported incidences of lion mortality in the Northern Tuli Game Reserve: 11 (58%) adults (5 male, 6 female) and eight (42%) subadults (3 male, 5 female). These deaths resulted from illegal killings (n = 6), trophy hunting in Tuli Circle (n = 1), poisoning (n = 6), snaring (n = 4) and problem-animal control (n = 2). A further 19 lions (12 males and 7 females) dispersed from the area during 2000–2011. They were not seen again and it is not known whether or not they survived. In the Venetia Limpopo Nature Reserve we recorded 20 cases of adult mortality (11 males, 9 females) and 10 cases of subadult mortality (4 males, 6 females) during 2005–2011. The majority (60%, n = 18) of these were trophy hunted, 37% (n = 11) were euthanized during a management intervention in 2009 to reduce the population size, and one lion was found dead outside the borders of the Reserve. In Northern Tuli the majority of recorded mortalities (94.7%, n = 18) occurred outside the Reserve, whereas in Venetia only one mortality was recorded outside the Reserve (Fig. 1) and the other mortalities were recorded within its borders.
Based on the methods of Hayward et al. (Reference Hayward, O'Brien and Kerley2007) the lion populations of Nothern Tuli and Venetia were estimated at 73 and 21, respectively, giving estimated population densities of 0.101 and 0.064 lions per km2, respectively. Although the mean population size was similar in both reserves for the duration of the study (18.6 ± SE 1.0 in Northern Tuli, 16.5 ± SE 1.8 in Venetia) there was a significant difference (U = 43; P < 0.05, Mann–Whitney U-Test) in population density between the reserves (mean 2.6 lions per 100 km2 in Northern Tuli and 5.0 lions per 100 km2 in Venetia; Fig. 2a). This was despite the significantly higher prey biomass in Northern Tuli (1,216 ± SD 176.6 kg per km2) compared to Venetia (375 ± SD 18.3 kg per km2; U = 0; P = 0.016, Mann–Whitney U-Test). Population growth rates for both reserves were low (Northern Tuli, 1.1 ± SE 0.1, range 0.6–1.8; Venetia, 1.0 ± SE 0.1, range 0.2–1.6) and did not vary substantially over the study period (Fig. 2b).
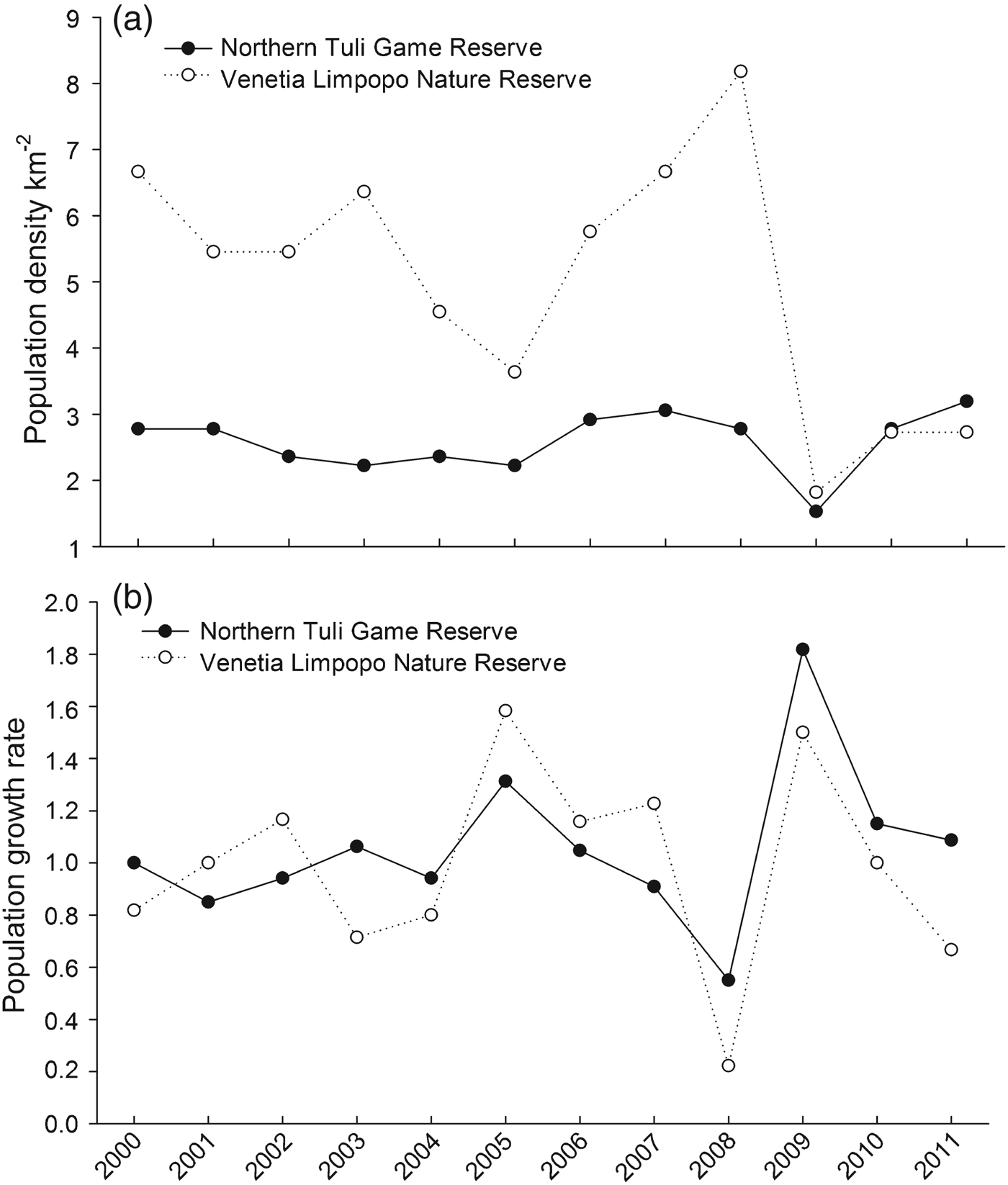
Fig. 2 Lion population density (a) and growth rates (b) in the Northern Tuli Game Reserve and Venetia Limpopo Nature Reserve (Fig. 1) during 2000–2011.
In addition to these population differences the total number of individuals in each pride differed significantly between Northern Tuli (4.7 ± SE 0.4, range 2–12) and Venetia (7.4 ± SE 1.1, range 3–10; U = 10.5; P < 0.05, Mann–Whitney U-Test). The mean number of adult females per pride was also significantly different (U = 13; P < 0.05, Mann–Whitney U-Test) between Northern Tuli (2.9 ± SE 0.2, range 2–4) and Venetia (2.1 ± SE 0.1, range 2–3).
During the study 23 litters of cubs (n = 60) were born in Northern Tuli and 11 litters (n = 28) were born in Venetia. In Northern Tuli eight cubs died before their sex could be determined, 81% (n = 48) reached 12 months of age, 65% (n = 38) reached 18 months, and 55% (n = 32) reached 24 months. The cub survival rate in Venetia was significantly higher, with 96% (n = 27) of cubs reaching 12 months of age (χ 2 = 5.13, df = 1, P < 0.05 with Yates correction), 93% (n = 26) reaching 18 months, and 86% (n = 24) reaching 24 months (Fig. 3). There was no significant difference (U = 102; P > 0.05, Mann–Whitney U-Test) in the mean litter size between Northern Tuli (2.6 ± SE 0.2, range 1–4) and Venetia (2.8 ± SE 0.2, range 1–4).

Fig. 3 Mean survival rates for lion cubs in the Northern Tuli Game Reserve and Venetia Limpopo Nature Reserve (Fig. 1) during 2000–2011.
On average, adults comprised 69% of the Northern Tuli population, with adult females outnumbering males by 1.7 : 1. Similarly in Venetia adults comprised 77% of the population, with females again outnumbering males by 1.7 : 1 (Table 1). The sex ratio among subadult lions in Northern Tuli was skewed slightly in favour of females (M : F, 1 : 1.4) and in Venetia was skewed slightly in favour of males (M : F, 1 : 0.9; Table 1). These differences were not significant (χ 2 = 0.60, df = 1, P > 0.05). The sex ratio amongst cubs was similar in both reserves (M : F, 1 : 0.9).
Table 1 Sex and age structure of lion Panthera leo populations in the Northern Tuli Game Reserve, Botswana, and Venetia Limpopo Nature Reserve, South Africa (Fig. 1), during 2000–2011.

Discussion
The importance of small lion populations
In a comprehensive inventory of lion populations, Bauer & Van Der Merwe (Reference Bauer and Van Der Merwe2004) documented 89 extant populations across Africa (for which population estimates were available). Of these populations 50 have an estimated 100 or fewer lions, with 33 populations having 50 or fewer. With so many small and isolated lion populations, the conservation and protection of these populations are important. Populations such as those in the Northen Tuli Game Reserve and the Venetia Limpopo Nature Reserve are exposed to a variety of sources of anthropogenic mortality. In Northern Tuli, which is largely unfenced, most human-caused lion mortalities are as a result of illegal activities such as snaring, poisoning and shooting, whereas in Venetia they are almost entirely legal and precipitated by management to control the population within the fenced reserve. Most of the lion mortalities from the Northern Tuli population occurred close to the Reserve boundary, indicating the intensity of edge effects in the absence of predator-proof fencing (Fig. 1). Lion numbers along the Shashe River, the boundary between Botswana and Zimbabwe, reportedly declined during 2000–2009, mainly as a result of snaring (B. Petty, pers. comm.).
Effects of different forms of anthropogenic mortality
The lion populations studied responded differently to different causes of mortality. The population density of the Northern Tuli lions was markedly lower than predicted based on prey biomass (Hayward et al., Reference Hayward, O'Brien and Kerley2007). This was associated with lower cub survival and significantly smaller prides, despite there being more adult females per pride in Northern Tuli. During 2000–2003 unselective hunting in the Venetia Limpopo Nature Reserve disrupted the population to such an extent that prides were absent during this period. Thus, excessively high and unselective mortality led to the breakdown of the fundamental fabric of lion society. Prides began to form again after indiscriminate and intense trophy hunting was halted in 2004 in favour of a more selective approach. These findings are useful for interpreting lion population structure in other areas.
Lions occurring in large, well-protected parks tend to live in prides of c. 12–15 individuals, with 4–6 adult lionesses per pride. Prides are defended by a coalition of males (Schaller, Reference Schaller1972; Smuts, Reference Smuts1976; Funston, Reference Funston2011). These populations are typically close to the predicted carrying capacity (Loveridge & Canney, Reference Loveridge and Canney2009; Ferreira & Funston, Reference Ferreira and Funston2010). However, there is an increasing body of literature that indicates that in many smaller protected areas, where lions are exposed to various intense forms of anthropogenic mortality, they do not occur at predicted densities (Kiffner et al., Reference Kiffner, Meyer, Mühlenberg and Waltert2009; Croes et al., Reference Croes, Funston, Rasmussen, Buij, Saleh, Tumenta and De Ionge2011; Groom et al., Reference Groom, Funston and Mandisodza2014) and populations comprise smaller prides and foraging groups (Bauer, Reference Bauer2003; Bauer et al., Reference Bauer, Nowell and Packer2012). These responses have been attributed to the ratios of lions to medium/large prey biomass (Bauer et al., Reference Bauer, Vanherle, Di Silvestre and De Iongh2008) or the effect of vegetation density (Trinkel et al., Reference Trinkel, Ferguson, Reid, Reid, Somers and Turelli2008). However, we suggest that the variability in group composition is more likely attributable to the combined effects of anthropogenic sources of mortality, and that once the adult segment of a lion population is reduced below a certain threshold the drivers for sociality (Packer et al., Reference Packer, Herbst, Pusey, Bygott, Hanby, Cairns, Borgerhoff Mulder and Clutton-Brock1988; Mosser & Packer, Reference Mosser and Packer2009) disappear, causing lion society to fragment. This is because prides are predominantly regulated by social factors (Van Orsdol, Reference Van Orsdol1984; Packer et al., Reference Packer, Scheel and Pusey1990, Reference Packer, Hilborn, Mosser, Kissui, Borner and Hopcroft2005), with food availability influencing these social mechanisms (Van Orsdol et al., Reference Van Orsdol, Hanby and Bygott1985).
Our study indicates that even when a lion population has been reduced below a certain threshold, cub survival and the rates of dispersal of subadults can nevertheless be maintained at high levels. As further evidence of a threshold effect no incidences of infanticide or inter-pride mortality were reported in either population, even though large groups of lionesses were not recorded. The lower survival rate of cubs in the Northern Tuli Game Reserve is probably a result of the high density of spotted hyaena Crocuta crocuta and may have a significant effect, given the small pride sizes and lack of male coalitions. Male lions typically reach maximum reproductive success at the age of 7–8 years (Smuts, Reference Smuts1978; Packer et al., Reference Packer, Herbst, Pusey, Bygott, Hanby, Cairns, Borgerhoff Mulder and Clutton-Brock1988). However, in both reserves we noted that almost all males that mated were < 5 years of age, often as young as 3 years. Furthermore, with relatively few adult males, we recorded several cases of mating between close relatives, such as mother and son (AS, pers. obs.). These examples indicate that processes that lead to inbreeding depression are likely to occur in lion populations affected by human persecution.
Conservation implications
Small, fragmented wilderness areas are difficult to manage and the wildlife populations inhabiting such areas are exposed to many threats that directly compromise their viability. Fencing off these areas further isolates the resident wildlife (Hayward & Kerley, Reference Hayward and Kerley2009). Current approaches attempt to alleviate these problems by creating large conservation areas with buffer zones and implementing conflict-mitigation initiatives (Woodroffe et al., Reference Woodroffe, Thirgood and Rabinowitz2005). One particular concept is that of transfrontier conservation areas, where fences between wildlife areas in two or more neighbouring countries are removed to create a larger protected area and facilitate free movement of animals, with a buffer zone between the protected areas and human settlements. Both the Northern Tuli Game Reserve and the Venetia Limpopo Nature Reserve fall within the proposed Greater Mapungubwe Transfrontier Conservation Area. Our results indicate the threats to carnivores in the area, which need to be addressed if the intended conservation goals are to be achieved. Illegal killing, poaching, problem-animal control, and loss of habitat are believed to be more serious threats to lion conservation than legalized safari hunting (Whitman & Packer, Reference Whitman and Packer1997), as highlighted by our results. Although fences limit the movement of some species they may also provide desirable protection for lion populations when the greater landscape harbours anthropogenic threats.
Acknowledgements
We thank President Lt Gen. Ian Khama Seretse Khama and the Department of Wildlife and National Parks, Botswana, for permission to conduct our research, Mashatu Game Reserve, Pete le Roux and David Evans for extensive financial and logistical support, Tshwane University of Technology for financial support, the Northern Tuli Game Reserve and all the land owners for transversing rights, and Warwick Mostert, Rox Brummer and Bruce Petty for their personal communications. At the Venetia Limpopo Nature Reserve data were collected by De Beers staff, students of Tshwane University of Technology, employees of the Endangered Wildlife Trust, and Global Vision International Wildlife Research Expeditions staff.
Biographical sketches
Andrei Snyman is head of the Northern Tuli Predator Project in eastern Botswana. His research focuses on the behavioural ecology and population dynamics of large African carnivores, and the conservation of the species. Craig Jackson's research focuses on the ecology, behaviour and distribution of African mammals, with a specific emphasis on conservation biology. Paul Funston has worked on the behavioural ecology and conservation of African lions for over 25 years. He oversees PANTHERA's lion programme, which is built on partnerships and funding models for lion conservation projects throughout Africa.




