



Introduction
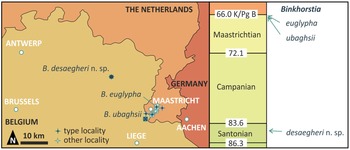
The brachyuran genus Binkhorstia Noetling, Reference Noetling1881 is known exclusively from the Upper Cretaceous of the south-east Netherlands and north-east Belgium (Figure 1). Until now, it comprised the type species, Dromilites ubaghsii van Binkhorst, Reference Binkhorst1857, occurring mainly in the upper Nekum Member (Maastricht Formation; upper Maastrichtian), and Binkhorstia euglypha Collins, Fraaye & Jagt, Reference Collins, Fraaye and Jagt1995, which appears to be restricted to the overlying Meerssen Member (Maastricht Formation; uppermost Maastrichtian) (see Fraaije, Reference Fraaije2003, fig. 5). Here we add a third species, B. desaegheri nov. sp., from the upper middle Santonian of Houthalen (Campine area, north-east Belgium).

Figure 1. Schematic map of the Maastricht (Netherlands) and Campine (Belgium) areas, with localities (quarries, outcrops) that have yielded species of Binkhorstia, and stratigraphical ranges of B. ubaghsii, B. euglypha and B. desaegheri nov. sp.
The history of Binkhorstia is convoluted, serving as a prime example of how attempts to unravel the higher-level taxonomic position of late Mesozoic crabs may prove difficult. Over time, the genus has been referred to various families or subfamilies, either podotreme or putative eubrachyuran, as follows:
-
Cyclodorippidae, by Feldmann & Villamil (Reference Feldmann and Villamil2002, p. 721);
-
Necrocarcinidae, by Collins (Reference Collins2003, p. 85), Collins & Jakobsen (Reference Collins and Jakobsen2004, p. 71) and Larghi (Reference Larghi2004, p. 530);
-
Torynommatidae, by Glaessner (Reference Glaessner1980, p. 181), De Grave et al. (Reference De Grave, Pentcheff, Ahyong, Chan, Crandall, Dworschak, Felder, Feldmann, Fransen, Goulding, Lemaitre, Low, Martin, Ng, Schweitzer, Tan, Tshudy and Wetzer2009, p. 29), Van Bakel et al., (Reference Van Bakel, Jagt, Fraaije, Coole, Jagt, Van Bakel and Fraaije2003, pp. 85–87) and Schweitzer et al., (Reference Schweitzer, Feldmann, Garassino, Karasawa and Schweigert2010, p. 78);
-
Dorippidae, by Glaessner (Reference Glaessner and Moore1969, p. R492) and Quayle & Collins (Reference Quayle and Collins1981, p. 738);
-
Carcineretidae, by Wright & Collins (Reference Wright and Collins1972, p. 91), Collins et al., (Reference Collins, Fraaye and Jagt1995, p. 199), Fraaye (Reference Fraaye1996, p. 271), Jagt et al., (Reference Jagt, Fraaye and Van Bakel2000, p. 40) (see also Wright, Reference Wright1997, p. 138);
-
Longusorbiidae, by Schweitzer & Feldmann (Reference Schweitzer and Feldmann2011, p. 16).
However, more recently it has been recognised that Binkhorstia would warrant a family of its own (Schweitzer & Feldmann, Reference Schweitzer and Feldmann2011, p. 16, Guinot et al., Reference Guinot, Tavares and Castro2013, p. 201). Members of this genus are characterised by a flattened, subsquare carapace, with undivided lateral margins, a wide thoracic sternum, a spatulate rostrum and reduced P5, positioned laterally to the first pleonal somite. We shall here examine the previous hypothesis put forward by Guinot et al., (Reference Guinot, Tavares and Castro2013, p. 201) that Binkhorstia might have affinities with members of the superfamily Retroplumoidea Gill, Reference Gill1894, erect a new family, Binkhorstiidae nov. and a new species, B. desaegheri nov. sp., and discuss its position within that superfamily.
The two late Maastrichtian species of Binkhorstia co-occur mainly with (in order of commonness) ghost shrimp, palaeocorystoids, raninoids, xanthoids and paguroids. The new late middle Santonian species is associated with the ghost shrimp Mesostylus faujasi (Desmarest, 1822) (see also Goolaerts & Mottequin, Reference Goolaerts and Mottequin2023, fig. 5B–D). Binkhorstia is known only from the study area and probably was endemic to it, being adapted to fairly fine-grained silty sands (Santonian) and biocalcarenitic intra-reefal environments of the upper Maastrichtian of the south-east Netherlands and surrounding areas (Figure 1).
Material and methods
Binkhorstia ubaghsii is fairly common and numerous specimens of variable carapace sizes are housed in the MAB and NHMM collections, inclusive of a few corpses (Figure 2) (see Jagt et al., Reference Jagt, Fraaye and Van Bakel2000; Van Bakel et al., Reference Van Bakel, Jagt, Fraaije, Coole, Jagt, Van Bakel and Fraaije2003). The second species, B. euglypha, is much less common. Following the description of the type specimen (Figure 3C; see Collins et al., Reference Collins, Fraaye and Jagt1995), a handful of additional individuals has been brought to our attention. The single specimen of the new species, B. desaegheri nov. sp. (Figure 3B), is housed in the IRSNB collections; this was found amongst the material collected from bulk sampling during construction of colliery shafts.

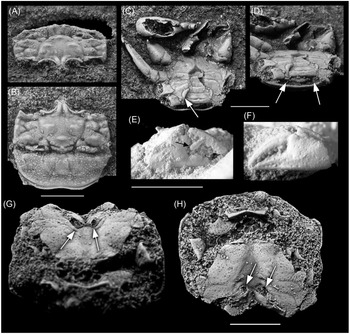
Figure 2. Binkhorstia ubaghsii (van Binkhorst, Reference Binkhorst1857): (A–D) male specimen (MAB 15928, leg. Robert Pieters) in oblique dorsal (A), dorsal (B), ventral (C) and oblique ventral (D) aspects, CBR-Romontbos quarry (Eben Emael, province of Liège, north-east Belgium), upper third of Nekum Member, Maastricht Formation (upper Maastrichtian), arrow in C indicates vertically oriented sternite 7, arrows in D indicate coxae of fifth pereiopods; (E, F) right (major) and left (minor) cheliped, arrow indicates molariform crushing tooth; (G, H) female specimen (MAB15929, leg. Yvonne Coole) in oblique ventral, and ventral view, arrows indicate vulvae at extremity of suture (see text for details), CBR-Romontbos quarry (Eben Emael, province of Liège, north-east Belgium), upper third of Nekum Member, Maastricht Formation (upper Maastrichtian). All scale bars equal 5 mm.

Figure 3. Carapaces (dorsal view) of Binkhorstia spp.: (A) Binkhorstia ubaghsii, for data see Figure 2; (B) B. desaegheri sp. nov., holotype (IRSNB 11512, shaft I of Houthalen colliery, eastern Campine Basin (north-east Belgium), depth 583–585 m below surface, Vaals Formation, either middle/upper part of Asdonk Member, or basal part of overlying Sonnisheide Member, upper middle Santonian; (C) B. euglypha Collins, Fraaye & Jagt, Reference Collins, Fraaye and Jagt1995, holotype (MAB15927), former Blom quarry (Berg en Terblijt, southern Limburg, the Netherlands), upper part of Meerssen Member, Maastricht Formation (uppermost Maastrichtian). Not to scale.
The following abbreviations are used in specimen descriptions: P1–P5, first to fifth pereopods (P1 as chelipeds; P2–P5 corresponding to ambulatory legs 1–4). Pleonal somites are numbered from 1 to 6. Thoracic sternal sutures are designated by the number of the two thoracic sternites that they involve and thus are numbered from 1/2 to 7/8.
Institutional abbreviations . RBINS, Royal Belgian Institute of Natural Sciences, Brussels, Belgium [note: IRSNB in collection registration]; MAB, Oertijdmuseum, Boxtel, the Netherlands; MGSB, Museo Geológico del Seminario de Barcelona, Barcelona, Catalonia, Spain; MNHN, Muséum National d’Histoire Naturelle Paris, France; NHMM, Natuurhistorisch Museum Maastricht, Maastricht, the Netherlands; NHMUK, Natural History Museum, London, UK.
Comparative material
The following extinct and present-day retroplumoid taxa have been compared in detail with representatives of the genus Binkhorstia:
Extant
-
1. Retropluma denticulata Rathbun, Reference Rathbun1932 – one male and two females (MNHN-IU 2022-3465, MNHN-IU-2022-3466), South China Sea, depths 99 m and 113 m (April 18, 1959);
-
2. Retropluma notopus (Alcock & Anderson, Reference Alcock and Anderson1894) (type species of Retropluma) – one male (MNHN-IU-2022-3462 = MNHN-B29874), Fiji, Campagnes MUSORSTOM 10;
-
3. Retropluma planiforma Kensley, Reference Kensley1969 – two males (MNHN-IU-2022-3463 = MNHN-B7015), Madagascar, Campagne CREVETTIERE 1973 (Crosnier Collection);
-
4. Retropluma plumosa Tesch, Reference Tesch1918 – two females (MNHN-IU-2022-3460 = MNHN-B28866), Indonesia, KARUBAR Expedition 1991;
-
5. Retropluma quadrata de Saint Laurent, Reference Saint Laurent and Forest1989 – male (holotype, MNHN-IU-2017-8034 = MNHN-B7009), Philippines, MUSORSTOM 1;
-
6. Retropluma serenei de Saint Laurent, Reference Saint Laurent and Forest1989 – male (holotype, MNHN-IU-2017-8033 = MNHN-B19531), north-west Philippines, MUSORSTOM 3;
-
7. Retropluma solomonensis McLay, Reference McLay, Richer de Forges and Justine2006 – male (holotype, MNHN-IU-2017-8024 = MNHN-B29870), Solomon Islands, SALOMON 1;
-
8. Bathypluma forficula de Saint Laurent, Reference Saint Laurent and Forest1989 – male (holotype, MNHN-IU-2017-8040 = MNHN-B11229), Strait of Makassar, CORINDON 2;
-
9. Bathypluma spinifer de Saint Laurent, Reference Saint Laurent and Forest1989 – male (holotype, MNHN-IU-2014-20496 = MNHN-B11226), Philippines, MUSORSTOM 2.
Fossil
-
1. Costacopluma concava Collins & Morris, Reference Collins and Morris1975 – holotype (NHMUK In. 44642) and paratypes (NHMUK In. 44643–44648), Campanian–?Maastrichtian, south-east Nigeria; and NHMUK IC 1284, Cenomanian of Texas (see Collins et al., Reference Collins, Garvie and Mellish2018);
-
2. Gaudipluma bacamortensis Artal, Van Bakel, Fraaije & Jagt, Reference Artal, Van Bakel, Fraaije and Jagt2013 – holotype (MGSB75283) and paratypes (MGSB75284, 75285a, b, 75286a, b, MAB15918, 15919), Ypresian (Lower Eocene), Bacamorta, Huesca, Spain;
-
3. Loerenthopluma danielae Van Bakel, Artal, Fraaije & Jagt, Reference Van Bakel, Artal, Fraaije, Jagt, Castro, Davie, Ng and Richer de Forges2010 – holotype (MAB15920) and twelve paratypes (MAB15921A–L), Ypresian (Lower Eocene), Egem, north-west Belgium;
-
4. Retrocypoda almelai Vía Boada, Reference Vía Boada1959 – holotype (MGSB 20123),, Lutetian (Middle Eocene), Catalonia, Spain;
-
5. Retropluma eocenica Vía Boada, Reference Vía Boada1959 – MGSB68417, upper Ilerdian (Lower Eocene), Bacamorta, Huesca, Spain); MGSB68420, middle Lutetian (Middle Eocene), Vilada, Catalonia, Spain; MGSB23738, middle Ilerdian (Lower Eocene), Terrades, Catalonia, Spain;
-
6. Retropluma gallica Artal, Van Bakel & Castillo, Reference Artal, Van Bakel and Castillo2006 – holotype (MGSB68408), paratype (MGSB68409) and additional material (MGSB68410–13), Ilerdian (Lower Eocene), Fontcouverte, Corbières, southern France;
-
7. Serrablopluma diminuta Artal, Van Bakel, Fraaije & Jagt, Reference Artal, Van Bakel, Fraaije and Jagt2013 – holotype (MGSB75287), paratypes (MGSB75289, MGSB75290a, b, MGSB78334a, b, MGSB78335a, b, c, MAB15922-15926), Priabonian (Upper Eocene), Yebra de Basa, Huesca, Spain.
Systematic palaeontology
Infraorder Brachyura Latreille, Reference Latreille1802
Section Eubrachyura de Saint Laurent, Reference Saint Laurent1980
Superfamily Retroplumoidea Gill, Reference Gill1894
Family Binkhorstiidae nov.
urn:lsid:zoobank.org:act:BF441D0A-0343-4B22-98F5-AAE3080027F8
Type, and sole, genus included. Binkhorstia Noetling, Reference Noetling1881.
Species included. In addition to the type species, Dromilites ubaghsii van Binkhorst, Reference Binkhorst1857, B. euglypha Collins, Fraaye & Jagt, Reference Collins, Fraaye and Jagt1995 and B. desaegheri nov. sp. (see below) (Figure 3).
Stratigraphical range. Late middle Santonian to latest Maastrichtian.
Palaeoenvironment. Sandy/silty and biocalcarenitic, shallow-water settings.
Diagnosis. Carapace small-sized, subrectangular to subsquare, widest at about one-third of total carapace length from front; flattened in both directions. Lateral margins weakly arched; anterolateral margins shorter than posterolateral margins. Carapace sides perpendicular to dorsal carapace surface, no linea brachyura visible. Posterior margin wide, slightly narrower than orbitofrontal margin, sinuous, axially concave, rimmed, bearing fine granules. Orbitofrontal width nearly equal to maximum carapace width; front narrow, with concave sides, longitudinally bilobed, distally conspicuously spatulate. Supraorbital margin with three short indentations, outer orbital corner developed as short triangular tooth. Infraorbital margin salient, long. Orbits deep, horizontal. Dorsal regions well-defined anteriorly may be with central tubercle. Cardiac grooves weak; anterior groove system complex, gastric grooves well defined. Cuticle surface entirely granular, granules may be clustered. Pterygostome narrow, inflated.
Chelae of male robust, heterochelous, heterodontous, palm short, fingers elongated, dactylus of major (right) chela with proximal crushing tooth; merus short, stout, globose. P2–P4 coxae in same plane, in ventral view progressively more closely spaced towards posterior. P2–P4 long, articles flattened. P5 coxae strongly reduced in size.
Thoracic sternum wide, subelliptical, flat; male sterno-pleonal cavity well-defined, narrow, deep, reaching halfway sternite 4; female sterno-pleonal cavity wider, shallow, subtriangular, reaching sternite 2. Episternites prominent, triangular. Sternite 5 widest, subhorizontal; sternite 6 narrower, slightly tilted towards posterior. Sternite 7 with exposed part conspicuously narrow, oriented obliquely; in females presence of distinct median (axial) line (i.e. medial line) (male disposition unknown). Sternite 8 completely covered by pleon in both sexes, likely distinctly reduced. Sternal sutures 4/5, 5/6 largely interrupted; suture 6/7 appearing to be complete (i.e. crossing thoracic sternum entirely). Pleon completely flexed under cephalothorax, all pleonal somites free. Sexual dimorphism markedly distinct: pleon wider in females, with narrow first somite. Pleonal holding structures not exposed in available material. Vulva at extremity of suture 5/6, suboval, large, closely spaced; aperture margin raised.
Genus Binkhorstia Noetling, Reference Noetling1881
Type species. Dromilites ubaghsii Van Binkhorst, Reference Binkhorst1857, by monotypy.
Binkhorstia desaegheri nov. sp.
Figs 3B
urn:lsid:zoobank.org:act:F70993F8-5C06-4E04-B23A-3C3F96017C91
Derivation of name. Named after Patrick De Saegher, RBINS volunteer, in recognition of his help in digitising the archives of the old ‘palaeontology department’, and in particular for his assistance in constructing a specimen-based inventory of Cretaceous invertebrates collected during shaft construction of collieries in the north-east Belgian Campine Basin, including the one at Houthalen that has yielded the type specimen of the new species.
Material. The holotype (maximum length 18 mm) (Figure 3B) is IRSNB 11512 (previously Invert-9780-0324), from a depth of 583–585 m below surface in shaft I of the Houthalen colliery, eastern Campine Basin, north-east Belgium (Figure 1). This specimen was initially registered on May 25, 1932 as IG (Inventaire Général) 9780. It was collected from the Vaals Formation, either from the middle/upper part of the Asdonk Member, or the basal part of the overlying Sonnisheide Member. The boundary between both members lies between a depth of 585 and 571 m in shaft I at Houthalen (for further discussion, see Goolaerts & Mottequin, Reference Goolaerts and Mottequin2023). This interval can be dated as late middle Santonian (Gonioteuthis westfalicagranulata belemnite Zone, sensu germanico). Associated ammonites include Placenticeras polyopsis (Dujardin, Reference Dujardin1837), which allows substantiating the Santonian age (for more details, see Jagt et al., Reference Jagt, Kennedy, Burnett, Christensen and Dhondt1995; Goolaerts & Mottequin, Reference Goolaerts and Mottequin2023).
Description. Although the carapace is compressed at an oblique angle during fossilisation, the carapace is essentially similar in outline to that of B. ubaghsii and B. euglypha (Figure 3). Orbitofrontal width very wide, equalling maximum carapace width, with two short notches, dividing into three triangular lobes; front narrow, with concave sides, longitudinally bilobed, distally not preserved. Well-defined dorsal regions anteriorly with curved epi- and mesobranchial lobes and a mesogastric lobe with a very pronounced and elongated, forwardly protruding anterior process. Clear, somewhat depressed urogastric. Cardiac grooves weak. Large single tubercles on protogastric, mesogastric, hepatic, epibranchial, mesobranchial and metabranchial regions, largest and dual tubercle on the cardiac region.
Cuticle microstructure is entirely granular.
Comparison. Binkhorstia desaegheri nov. sp. Differs from B. ubaghsii and B. euglypha in having a longer and more pronounced mesogastric process which also continues more anteriorly. Binkhorstia desaegheri nov. sp. has also the most distinct tuberculose carapace (see Figure 3).
Discussion
Several Cretaceous crab genera present dorsal carapaces that are superficially similar to that of Binkhorstia, such as Longusorbis Richards, Reference Richards1975, Carcineretes Withers, Reference Withers1922 and Ophthalmoplax Rathbun, Reference Rathbun1935. Spiridonov (Reference Spiridonov2020, p. 144) noted for these genera that, ‘In general, the carapace outline and symmetry/asymmetry patterns are characters of considerable taxonomic value at the genus or family level’. Similarities in orbitofrontal configuration are obvious: a narrow front, wide orbits, short supramarginal notches, similar supramarginal teeth, and a rather similar disposition of carapace grooves and regions. Rather than being a podotreme, as has often been thought, Binkhorstia is an eubrachyuran, clearly demonstrated by the vulvae preserved in one specimen before us, so that comparisons below will exclusively relate to eubrachyuran families.
The new family, Binkhorstiidae nov., is here assigned to the Retroplumoidea, based on a set of characters (Figures 2 and 3) that leads to a configuration known only in members of this superfamily, as follows: flattened, squarish to rectangular carapace; vertical anterolateral and posterolateral margins undefined from each other; angular lateral margin with perpendicular walls; wide orbits that occupy the entire frontal margin and are visible dorsally; thin, spatulate rostrum; P5 coxae conspicuously reduced, placed posteriorly and adjacent to the pleon (see Figure 2D); subequal chelipeds; flattened P2–P4; long pleon with all somites free; presence of a rainure in front of the sterno-pleonal cavity (i.e. in front of the telson) in which are lodged the extremities of the first gonopods in males and female pleopods; wide thoracic sternum, with long sutures; strongly reduced thoracic sternite 8, with exposed part so tiny that it is barely discernible.
The superfamily Retroplumoidea, revised by de Saint Laurent (Reference Saint Laurent and Forest1989) and until now considered to contain only a single family, the Retroplumidae (Martin & Davis, Reference Martin and Davis2001; Števčić, Reference Števčić2005; Ng et al., Reference Ng, Guinot and Davie2008), composed of only two extant genera, Retropluma Gill, Reference Gill1894 and Bathypluma de Saint Laurent, Reference Saint Laurent and Forest1989, but involving a much higher number of extinct genera. It is considered an ‘ancient’, deeply rooted clade (Guinot et al., Reference Guinot, Tavares and Castro2013, 2019), an assertion supported recently by genetic analyses (Wolfe et al., Reference Wolfe, Ballou, Luque, Watson-Zink, Ahyong, Barido-Sottani, Chan, Chu, Crandall, Daniels, Felder, Mancke, Martin, Ng, Ortega-Hernández, Theil, Palacios, Pentcheff, Robles, Thoma, Tsang, Wetzer, Windsor and Bracken-Grissom2023) and supported by the Cretaceous occurrence of Retroplumidae (Costacopluma Collins & Morris, Reference Collins and Morris1975; Cristipluma Bishop et al., Reference Bishop1983) and Binkhorstiidae fam. nov.
Several characters of the type species, B. ubaghsii, are fairly unique, notably: the heterochelous and heterodontous chelipeds with the presence of a molariform tooth on the larger chela (being indicative of a shell-breaking behaviour; see Schäfer, Reference Schäfer1954; Van Bakel et al., Reference Van Bakel, Jagt, Fraaije, Coole, Jagt, Van Bakel and Fraaije2003, figs 1.1, 1.2; Dietl & Vega, Reference Dietl and Vega2008, p. 292; Schweitzer & Feldmann, Reference Schweitzer and Feldmann2010), as well as the narrow, vertically oriented thoracic sternite 7 in the male (unknown in the female). These traits are not shared by any extant retroplumid (see de Saint Laurent, Reference Saint Laurent and Forest1989) that normally have a long male pleon, with all somites free (3, 4 and 5 fused in extant retroplumids); the male pleon with somite 6 ridged and laterally expanded in its posterior part (corresponding to developed locking pleonal structures; see Guinot, Reference Guinot1979, fig. 30C; even present, yet less marked in females; see Guinot, Reference Guinot1979, fig. 30E; de Saint Laurent, Reference Saint Laurent and Forest1989, figs 13, 15), and a wide thoracic sternum with exposed part of sternite 7 directed obliquely (Figure 2C). In extant retroplumid species, pleonal somites 3–5 are fused in adult males but free in females (de Saint Laurent, Reference Saint Laurent and Forest1989, fig. 13), which is not the case in either B. ubaghsii or B. desaegheri nov. sp. In addition, extant retroplumids show a dorsal carapace with crests in species of Retropluma (de Saint Laurent, Reference Saint Laurent and Forest1989, figs 2, 11, 12, 14), but this is much attenuated in Bathypluma. The vertically oriented thoracic sternite 7 may be considered a basal trait; it is also known in other basal eubrachyurans such as Componocancer Feldmann, Schweitzer & Green, Reference Feldmann, Schweitzer and Green2008, Eogeryon Ossó, Reference Ossó2021, Marocarcinus Guinot, De Angeli & Garassino, Reference Guinot, De Angeli and Garassino2008 and Styracocarcinus Schweitzer & Feldmann, Reference Schweitzer and Feldmann2012.
All these features support the erection of a new family, Binkhorstiidae nov., the third to be recognised within the Retroplumoidea. Details of in-depth comparisons between the new family and other eubrachyuran families are as follows:
-
1. Binkhorstiidae vs Retroplumidae
A diagnosis of the Retroplumidae was provided by for example, Davie et al., (2015: 1108) and Feldmann & Schweitzer (Reference Feldmann and Schweitzer2018, p. 1).
Guinot et al., (Reference Guinot, Tavares and Castro2013, p. 201) stated that Binkhorstia could have affinities with the Retroplumoidea, based on ‘the wide thoracic sternum, spatulate rostrum, flattened P2–P4, and P5 which are reduced and placed laterally to the first abdominal somite’. In fact, Binkhorstia shares several characters with the Retroplumidae, including small size, a dorsoventrally flattened body; subrectangular or subsquare carapace; sharp lateral margins; carapace with vertical flanks and long posterior margin; orbitofrontal margin usually nearly as wide as carapace; large orbits; narrow, deflected and spatulate front; flat thoracic sternum, sternite 8 covered by male pleon (sternite 8 hardly discernible in Retroplumidae, see Guinot et al., Reference Guinot, Tavares and Castro2013, fig. 34B, D); long and flattened P2–P4; P5 coxae much more closely interspaced; at base of pleon, P5 coxae distinctly reduced (see Figure 2C, D); subcircular and large vulvae, at extremity of suture 5/6 (see fig. 6). In Retroplumidae, however, the carapace regions or grooves are weakly defined (regions well marked, defined by a complex system of grooves in Binkhorstiidae nov.); usually three blunt to acute crests are present on the dorsal surface (absent in Binkhorstiidae nov.); the carapace cuticle is without dense granulation or tuberculation (present in Binkhorstiidae nov.); and the supraorbital margin is continuous, without notches or teeth (notches and teeth present in Binkhorstiidae nov.). Additionally, in Retroplumidae sternite 3 is short, horizontal, undivided (low triangular, with median depression in Binkhorstiidae nov.); sternite 4 bears a distinct, curved tooth (not visible in Binkhorstiidae nov.); sternites 5, 6 are horizontal, with rather parallel long sutures (curved, with more or less oblique sutures in Binkhorstiidae nov.); sternite 7 similar to preceding sternites (conspicuously narrow and oblique in Binkhorstia); episternites are small (robust in Binkhorstiidae nov.); a salient prominence on sternite 5 corresponds to the structure for pleonal holding, see Guinot & Bouchard, Reference Guinot and Bouchard1998, fig. 17B (absent in Binkhorstiidae nov.); pleonal somite 6 is typically modified by a marked lateral projection for pleonal holding, see Guinot & Bouchard, Reference Guinot and Bouchard1998, fig. 17A (normal, i.e. rectangular, in Binkhorstiidae nov.) and all sternal sutures are incomplete (suture 6/7 complete in females of Binkhorstiidae nov.; fig. 2H) (males unknown).
Fossil and extant retroplumid crabs (see Artal et al., Reference Artal, Van Bakel, Fraaije and Jagt2013, pp. 348, 351, figs 2D, 4F) have a fairly well-developed vulva near the extremity of suture 5/6 (Guinot, Reference Guinot1979, fig. 30E; de Saint Laurent, Reference Saint Laurent and Forest1989, fig. 5); Binkhorstiidae nov. has conspicuously large apertures (Figure 2G, H). Additionally, in extant retroplumids the chelipeds are flattened, with slender fingers which lack shell-breaking structures. These additional differences warrant exclusion of Binkhorstia from the Retroplumidae. However, the numerous similarities allow Binkhorstia to be accommodated within the Retroplumoidea, assigned to its own family.
Several extinct retroplumid crabs have been described, in particular from the Eocene, some with a well-preserved sternum. The monotypical, small-sized Serrablopluma Artal, Van Bakel, Fraaije & Jagt, Reference Artal, Van Bakel, Fraaije and Jagt2013, from the upper Eocene (Priabonian) of Huesca (north-east Spain), shows a typical retroplumid sternum (see Artal et al., 2023, figs 1, 2) with a subtriangular (narrow anterior) sternite 4, a normal (wide, developed) sternite 7, and a short, rather wide sternite 8 (Artal et al., 2023, fig. 2d), unlike the configuration in Binkhorstiidae nov. In Serrablopluma the pleonal somites have horizontal crests, the episternites being of normal development (large in Binkhorstia).
The large-sized and distinctly ornamented Gaudipluma Artal, Van Bakel, Fraaije & Jagt, Reference Artal, Van Bakel, Fraaije and Jagt2013 (see Artal et al., Reference Artal, Van Bakel, Fraaije and Jagt2013, Figs 3, 4) shows a characteristic thoracic sternum with oblique suture 4/5, a rather narrow female pleon, a distinct median line up to the level of suture 5/6, a conspicuously large and wide sternite 7, a sternite 8 that is the only small, inclined and subvertical sternite, and virtually absent episternites. In all these characters, Gaudipluma differs from Binkhorstiidae nov. In Gaudipluma the sternites 3, 4 are trapezoidal, and the anterior sternite 4 is wide, which is closer to Binkhorstia than to other retroplumids.
Retrocypoda Vía Boada, Reference Vía Boada1959 is a retroplumoid known from a single species, R. almelai Vía Boada, Reference Vía Boada1959, widely distributed in northern Spain, Italy and Iran (see de Saint Laurent, Reference Saint Laurent and Forest1989, pl. 7A-D; Khodaverdi Hassan-vand et al., Reference Khodaverdi Hassan-vand, Bahrami, Yazdi, Ossó, Safari, Martínez and Vega2016), with an unpublished record from Hungary (M. Hyžný, pers. comm., May 2024). It is characterised by a carapace with subhorizontal and oblique sinuous crests (see Khodaverdi Hassan-vand et al., Reference Khodaverdi Hassan-vand, Bahrami, Yazdi, Ossó, Safari, Martínez and Vega2016, fig. 4), pleonal somites and wide thoracic sternites with strongly developed horizontal crests, and, as befits the family, large and robust chelipeds (Ferratges, Reference Ferratges2017, fig. 30C, D). These characters do not match those of the Binkhorstiidae nov.
Loerenthopluma Beschin, Busulini, De Angeli & Tessier, Reference Beschin, Busulini, De Angeli and Tessier1996, known from two species from the Eocene–Oligocene of Belgium, Italy and Hungary (see Beschin et al., Reference Beschin, Busulini, De Angeli and Tessier1996; Hyžný & Müller, Reference Hyžný and Müller2010; Van Bakel et al., Reference Van Bakel, Artal, Fraaije, Jagt, Castro, Davie, Ng and Richer de Forges2010), was ‘considered to be closely related to Retropluma’ (Van Bakel et al., Reference Van Bakel, Artal, Fraaije, Jagt, Castro, Davie, Ng and Richer de Forges2010, p. 46). Indeed, the thoracic sternum and pleon of Loerenthopluma are closely similar to those of Retropluma (compare Van Bakel et al., Reference Van Bakel, Artal, Fraaije, Jagt, Castro, Davie, Ng and Richer de Forges2010, fig. 1, and Beschin et al., Reference Beschin, Busulini, De Angeli and Tessier1996, fig. 4.3).
Several fossil species of Retropluma are known. Retropluma minuta Gašparič, Hyžný, Jovanović, Ćorić & Vrabac, Reference Gašparič, Hyžný, Jovanović, Ćorić and Vrabac2019, from the middle Miocene of Slovenia, is small sized, has a squarish carapace and slender chelipeds; pleonal segment 6 is widened for the pleonal locking system, and somites 3–5 are apparently fused. The genus appears to have originated in the early Eocene. Retropluma eocenica is first known from upper Ilerdian (lower Eocene) deposits in Spain; R. gallica from the Ilerdian of Corbières, southern France.
As far as the Cretaceous is concerned, Costacopluma is known from the Coniacian and survived the K/Pg event into the Eocene; it is widespread from Nigeria and Senegal to Greenland, from the USA to Brazil and Argentina (see Hyžný et al., Reference Hyžný, Perrier, Robin, Martin and Sarr2015, table 1; fig. 9). Costacopluma is characterised by a subovate carapace outline, with distinct, inflated, transverse crests on the dorsal surface, devoid of distinct tubercles as in Binkhorstia. The thoracic sternites are wide and show acute oblique ridges (flat in Binkhorstia), the episternites are weakly developed (conspicuously large in Binkhorstia), and the vulvae on sternite 6 are smaller (compare Armstrong et al., Reference Armstrong, Nyborg, Bishop, Ossó-Morales and Vega2009, fig. 5.17).
In summary, characters shared by the families Retroplumidae and Binkhorstiidae nov. are sternite 8 very reduced, not dorsally visible or hardly discernible; a similar rainure in front of the sterno-pleonal cavity (in which extremities of the first gonopods in males and of pleopods in females are lodged); the vulva at the extremity of suture in Binkhorstiidae nov. (fig. 6) as in extant retroplumids (Guinot, Reference Guinot1979, fig. 30E; de Saint Laurent, Reference Saint Laurent and Forest1989, fig. 5; Guinot et al., Reference Guinot, Tavares and Castro2013, fig. 5D).
Binkhorstiidae vs Longusorbiidae
Schweitzer & Feldmann (Reference Schweitzer and Feldmann2011, p. 16) provisionally placed Binkhorstia in the Longusorbiidae Karasawa, Schweitzer & Feldmann, Reference Karasawa, Schweitzer and Feldmann2008, a family erected to contain a single genus, Longusorbis, which includes three species (see Schweitzer et al., Reference Schweitzer, Feldmann, Garassino, Karasawa and Schweigert2010, p. 106). Schweitzer & Feldmann (Reference Schweitzer and Feldmann2011, p. 16) listed the similarities between Binkhorstia and Longusorbiidae, noting that, ‘a front that can be interpreted as lying between the interior-most orbital notches; a long, spatulate rostrum with a strongly downturned tip; very broad, sinuous, notched orbits; a fronto-orbital width that is about equal to the maximum carapace width; short gastric regions and long branchial regions; sternites 1 and 2 fused; sternal suture 3/4 developed as a notch laterally; sternite 4 long; sternal sutures 4/5 and 5/6 not parallel; sternite 8 not visible in ventral view; and all male abdominal somites free’. The same authors also drew attention to a number of significant differences: ‘The male abdomen does not seem to fill the entire space between the coxae of the fifth pereiopods in Binkhorstia, and the overall carapace shape and ornamentation is different in Binkhorstia than in Longusorbis’. In addition to these differences, we note that:
-
- Longusorbis has a subtrapezoidal carapace (subrectangular in Binkhorstia);
-
- Longusorbis lacks a shell-breaking tooth on the major chela dactylus (present in Binkhorstia);
-
- Longusorbis has a normal P5 (strongly reduced in Binkhorstia);
-
- Longusorbis has a normal thoracic sternite 7 (narrow and obliquely directed in Binkhorstia).
Binkhorstia cannot be included in the Longusorbiidae on the basis of the differences outlined above. Longusorbiids are currently included in the superfamily Portunoidea (Spiridonov, Reference Spiridonov2020), a position is in need of further testing.
Binkhorstiidae vs Dorippoidea
Glaessner (Reference Glaessner and Moore1969, p. R492) assigned Binkhorstia to the Dorippidae H. Milne Edwards, Reference Milne Edwards1837, followed by Quayle & Collins (Reference Quayle and Collins1981, p. 738). The squarish carapace with well-defined anterior regions of B. ubaghsii (only with an ornament of granules and tubercles in B. euglypha and B. desaegheri nov. sp.) does not resemble the ovate dorippoid carapace that has special grooves, often delineating a human-like face. Several other characters exclude Binkhorstiidae nov. from the Dorippoidea (see Guinot, Reference Guinot2023, p. 370), such as:
-
- P4 and P5 reduced in Dorippoidea (only P5 in Binkhorstiidae nov.);
-
- posterolateral margin not well defined and with perpendicular edge in Dorippoidea (well-defined, perpendicular in Binkhorstiidae nov.);
-
- Dorippoidea without a shell-breaking tooth on major chela dactylus (present in Binkhorstiidae nov.);
-
- pleon curved, with first somites dorsally exposed, in prolongation with carapace in Dorippoidea (entirely flexed, held under body in Binkhorstiidae nov.);
-
- rostrum bidentate, with more or less concave axial notch in Dorippoidea (narrow, spatulate in Binkhorstiidae nov.);
-
- thoracic sternum strongly flexed posteriorly in Dorippoidea (flat in Binkhorstiidae nov.); thoracic suture 5/6 strongly concave (normal in Binkhorstiidae nov.);
-
- sternite 8 with exposed part dorsally visible (see Guinot, Reference Guinot2023, fig. 8B, C) (strongly reduced in Binkhorstiidae nov.).
The extinct dorippoid family Telamonocarcinidae Larghi, Reference Larghi2004, originally erected as a subfamily, but elevated to family status by Guinot et al., (Reference Guinot, Tavares and Castro2013, p. 306), and recently re-considered (Luque, Reference Luque2015; Van Bakel et al., Reference Van Bakel, Mychko, Spiridonov, Jagt and Fraaije2023), now contains the genera Eodorippe Glaessner, Reference Glaessner1980 and Telamonocarcinus Larghi, Reference Larghi2004 (type genus). Telamonocarcinids differ from Binkhorstia, in addition to differences with the Dorippoidea listed above, in lacking orbital fissures and spines and in having strong outer orbital spines.
Binkhorstiidae vs Carcineretidae
On several occasions, Binkhorstia has been placed in the Carcineretidae Beurlen, Reference Beurlen1930, for instance by Wright & Collins (Reference Wright and Collins1972, p. 91), Collins et al., (Reference Collins, Fraaye and Jagt1995, p. 199), Fraaye (Reference Fraaye1996b, p. 271) and Jagt et al., (Reference Jagt, Fraaye and Van Bakel2000, p. 40), in view of an overall similar appearance: squarish, flat carapace with wide orbits, and regions ornamented with tubercles. At the time, the Carcineretidae also contained Ophthalmoplax, which later was transferred to the Portunoidea (Polybiniidae; see Schweitzer et al., Reference Schweitzer, Feldmann and Karasawa2007, p. 22) and subsequently to the portunoid family Macropipidae by Karasawa et al., (Reference Karasawa, Schweitzer and Feldmann2008, p. 100). Ossó-Morales et al., (Reference Ossó-Morales, Artal and Vega2010, p. 219) provisionally concurred with that placement. The type genus Carcineretes has strongly developed P5 with flattened distal elements, transformed into highly specialised swimming tools (see Jagt et al., Reference Jagt, Van Bakel, Guinot, Fraaije, Artal, Castro, Guinot, Schram and Von Vaupel Klein2015, fig. 71-15.2C), very unlike the conspicuously reduced P5 of Binkhorstia. The front in Carcineretes is rather wide, downturned but not spatulate (vs very narrow and spatulate in Binkhorstiidae nov.). In Carcineretes, the male pleonal somites 3–5 are fused, and the male pleonal somites 1 and 2 are in contact with the P5 coxae, unlike the disposition in the new family. Thoracic sternite 7 is not conspicuously narrow and vertical, but similar to the preceding thoracic sternites. The lower margin of the chelae is sharply rimmed, while that of Binkhorstia is rounded. The dentation, including a shell-crushing molar, is otherwise similar. These differences exclude Binkhorstia from the Carcineretidae.
Binkhorstiidae vs Archaeopidae
The monotypical family Archaeopidae Karasawa, Kishimoto, Ohara & Ando, Reference Karasawa, Kishimoto, Ohara and Ando2019, recently established to accommodate the genus Archaeopus Rathbun, 1908 (type species, by monotypy, A. antennatus Rathbun, 1908), contains eight species of Late Cretaceous to Eocene age from the Pacific coast of North America and Japan (Nyborg et al., Reference Nyborg, McLachlan, Garassino, Vega, Phillippe and Champagne2019). This had previously been included in the Retroplumidae, but this status was considered doubtful (see de Saint Laurent, Reference Saint Laurent and Forest1989; McLay, Reference McLay, Richer de Forges and Justine2006; Guinot et al., Reference Guinot, Tavares and Castro2013). The genus was described at length by Guinot et al., (Reference Guinot, Tavares and Castro2013, p. 140), and species were revised and added by Nyborg et al., (Reference Nyborg, McLachlan, Garassino, Vega, Phillippe and Champagne2019, p. 48). It was distinguished from the Retroplumidae on the basis of carapace, sternal and pleonal characters (Karasawa et al., Reference Karasawa, Kishimoto, Ohara and Ando2019).
The female thoracic sternum of the type species, A. antennatus, shows a conspicuously large, elongate pleonal somite 6 (see Nyborg et al., Reference Nyborg, McLachlan, Garassino, Vega, Phillippe and Champagne2019, fig. 5B). The thoracic sternum of Archaeopus is subcircular, wide, with oblique thoracic sternites, which are weakly ridged. Sternite 8 is covered by the male pleon, P5 appears to be strongly reduced. The carapace lateral margins bear several blunt spines and nodes, more or less comparable with Binkhorstia.
Members of Binkhorstiidae nov. appear to be similar in dorsal aspect to Archaeopidae, sharing the subsquare carapace outline, the undifferentiated, subvertical and sharp-edged lateral margins, the horizontal orbital margin that is practically as wide as the carapace, the narrow and spatulate rostrum (at least in some members of the Archaeopidae; see Nyborg et al., Reference Nyborg, McLachlan, Garassino, Vega, Phillippe and Champagne2019, figs 7, 8) and the reduced P5. However, there are several significant differences between the Archaeopidae and Binkhorstiidae nov., including:
-
Archaeopids have inflated carapace regions, with horizontally elongated nodes on the dorsal carapace (weakly vaulted regions, only in gastric area with central tubercle in Binkhorstiidae nov.);
-
In archaeopids, the lowest point of the cervical groove, thus the gastric pits, are below carapace mid-length (rgastic pits more anteriorly situated, above carapace mid-length in Binkhorstiidae nov.);
-
Archaeopids have very wide protogastric regions (narrow in Binkhorstiidae nov.);
-
In archaeopids, thoracic sternites 3 and 4 are clearly distinguished by a prominent, oblique groove (only a weak lateral incision in Binkhorstiidae nov.);
-
Female sternoabdominal depression stops on sternite 4 in Archaeopidae (see Nyborg et al., Reference Nyborg, McLachlan, Garassino, Vega, Phillippe and Champagne2019, fig. 5B), but continues up to sternite 2 in Binkhorstiidae nov.;
-
Male sternite 7 is normal in Archaeopidae (arched, narrow and oriented obliquely in Binkhorstiidae nov.);
-
Chelae in archaeopids are robust, with long, pointed fingers without shell-crushing molar (small, more compact and with shell-crushing molar in Binkhorstiidae nov.).
Binkhorstia cannot be included in the Archaeopidae because of the differences outlined above. Both families could be basal retroplumoids.
Binkhorstiidae vs Lithophylacidae
The monotypical Lithophylacidae Van Straelen, Reference Van Straelen1936, occurring in the Cenomanian (Upper Cretaceous) of France (see Guinot & Breton, Reference Guinot and Breton2006), shows some similarities to Binkhorstia in that P5 is strongly reduced, the orbital margin occupies the maximum carapace width and there is a narrow, spatulate rostrum. Lithophylacids were considered as basal Portunoidea by Karasawa et al., (Reference Karasawa, Schweitzer and Feldmann2008, p. 84), and regarded as closely related to the Icriocarcinidae (Phillips et al., Reference Phillips, Nyborg and Vega2013, p. 151; Teodori et al., Reference Teodori, Ossó, Nyborg, Vega and Krobicki2013, p. 73). Lithophylax differs from Binkhorstia in numerous ways; most significantly, Lithophylax has:
-
transverse ridges on the carapace;
-
a carapace that is wider than long;
-
strongly divergent lateral margins;
-
no orbital notches and teeth;
-
the female sterno-pleonal depression does not reach sternite 3 (crossing sternite 3 in Binkhorstia);
-
thoracic sternite 7 is not conspicuously narrow and directed obliquely directed, but normal, as preceding sternites;
-
there is no shell-crushing tooth on the major chela dactylus;
-
there is a stridulating apparatus in Lithophylax, absent in Binkhorstia.
Binkhorstiidae vs Ophthalmoplax
Ophthalmoplax, previously included in the portunoid family Macropipidae (see Karasawa et al., Reference Karasawa, Schweitzer and Feldmann2008, p. 100; Ossó-Morales et al., Reference Ossó-Morales, Artal and Vega2010, p. 215; Schweitzer et al., Reference Schweitzer, Feldmann, Garassino, Karasawa and Schweigert2010, p. 107), currently included in the portunoid family Carcinidae (Polybiinae) (Feldmann et al., Reference Feldmann, Schweitzer and Goedert2018: 580), presents a squarish carapace with wide orbits, a sinuous orbital margin divided by short notches, a relatively well-marked gastrohepatic groove, and discontinuous horizontal crests [i.e. widened tubercles] in dorsal regions. However, the rostrum is wide, with four blunt nodes in two different planes and P5 is well-developed, flattened and paddle-like (Schweitzer et al., Reference Schweitzer, Feldmann and Karasawa2007, p. 18, table 1; Vega et al., Reference Vega, Phillips, Nyborg, Flores-Ventura, Clements, Espinosa and Solís-Pichardo2013, fig. 3.4). Ossó-Morales et al., (Reference Ossó-Morales, Artal and Vega2010, p. 219) mentioned that Ophthalmoplax might warrant its own subfamily.
Binkhorstiidae vs Palicidae
McLay (Reference McLay, Richer de Forges and Justine2006: 375) opined that some of the fossils placed in the Retroplumidae probably belong rather to the Palicidae Bouvier, 1898. Palicids have a subquadrate to rounded carapace outline, granular carapace surface, a sharp lateral margin, and P5 strongly reduced; these characters are shared with Binkhorstiidae fam. nov. The rostrum of Palicidae typically is wide, with multiple spines or lobes; a different morphology as in Binkhorstiidae fam. nov. or any other Retroplumoidea. The P5 coxae are placed wide apart in Palicidae, unlike the narrow placement in Binkhorstiidae fam. nov. Also, the female thoracic sternum of Palicidae shows a distinctive configuration, with anterior advancement of sternites (Guinot et al., Reference Guinot, Tavares and Castro2013: 47) and the vulvae (see Guinot, Reference Guinot1979: fig. 31), which is unlike the disposition in Binkhorstia.
Conclusions
Members of the family Binkhorstiidae nov. are known exclusively from Santonian and upper Maastrichtian strata in the extended type area of the Maastrichtian Stage (south-east Netherlands and north-east Belgium). The groups appear to have been endemic to the area, ranging from the upper middle Santonian (siliciclastics) to uppermost Maastrichtian (carbonates). The history of Binkhorstia is a turbulent one; its higher-level taxonomic position has changed considerably over time, having been assigned to various families or subfamilies, either podotreme or putative eubrachyuran. Here we place Binkhorstiidae nov. in the superfamily Retroplumoidea, following a detailed comparison with the families to which Binkhorstia had previously been referred.
Acknowledgements
We thank Paula Martin-Lefèvre (MNHN, Direction Générale Déléguée aux Collections – Invertébrés) for curatorial support, and the journal reviewers, Àlex Ossó (Tarragona, Iberia) and Matus Hyžný (Comenius University Bratislava, Slovakia), for constructive comments on an earlier version of the typescript.


 Open access
Open access