



Ritual killings or sacrifices were an important component of the cultural life of various prehispanic populations in the Andean region, with both human and domestic animal sacrifices being practiced (Goepfert Reference Goepfert2012; Klaus and Toyne Reference Klaus and Toyne2016; Verano Reference Verano and Dillehay1995, Reference Verano, Benson and Anita2001). Domestic camelids, namely llamas (Lama glama) and alpacas (Vicugna pacos), were the principal animals involved in such practices (Goepfert Reference Goepfert2011, Reference Goepfert2012; Wheeler et al. Reference Wheeler, Russel and Redden1995). The sacrifices could have fulfilled several functions and taken various forms (Goepfert Reference Goepfert2012). However, all the sacrificed animals were part of a supernatural gift economy in a highly ritualized context, involving a set of complex relations between the living and the spirits and ancestors (Flores Ochoa et al. Reference Flores Ochoa, MacQuarrie and Portus1994; Szpak et al. Reference Szpak, Millaire, White, Bourget, Longstaffe, Klaus and Toyne2016). Recent studies have explored the life history of the sacrificed individuals, in particular their geographical origin (Andrushko et al. Reference Andrushko, Buzon, Gibaja, McEwan, Simonetti and Creaser2011; Tung and Knudson Reference Tung and Knudson2010) and the special treatments they received. For example, the children sacrificed by the Inca in a practice known as capacocha had a special diet in the months preceding their death. Notably, the Llullaillaco children discovered in Argentina seem to have ingested alcohol and coca in the months before their death (Wilson et al. Reference Wilson, Brown, Villa, Lynnerup, Healey, Ceruti, Reinhard, Previgliano, Araoz, Diez and Taylor2013). This raises the question whether a special final meal was also given to these camelids before they were sacrificed.
This article aims to understand the final diet of the sacrificed domestic camelids from the Huanchaquito-Las Llamas (HLL) site, which is located on the northern coast of Peru (Figure 1). HLL, dated to the fifteenth century AD in the Late Intermediate period (LIP), is thus far a unique example of such a context because of the exceptional preservation of the animal corpses due to the arid climatic conditions that prevail on the Pacific coast (Goepfert and Prieto Reference Goepfert, Prieto, Capriles and Tripcevich2016; Goepfert et al. Reference Goepfert, Elise Dufour and Verano2020; Prieto et al. Reference Prieto, Verano, Goepfert, Kennett, Quilter, Leblanc, Fehren-Schmitz, Forst, Lund, Dement, Dufour, Tombret, Calmon, Gadison and Tschinkel2019). Although the exact reasons for this mass killing are unknown, one motivation could be linked to a sudden climatic event (Prieto et al. Reference Prieto, Verano and Goepfert2017, Reference Prieto, Verano, Goepfert, Kennett, Quilter, Leblanc, Fehren-Schmitz, Forst, Lund, Dement, Dufour, Tombret, Calmon, Gadison and Tschinkel2019). Various studies have already been done on some of these animal corpses. The paleoparasitological analysis of gut contents and feces by Le Bailly and colleagues (Reference Le Bailly, Goepfert, Prieto, Verano and Dufour2020) described the internal parasites and aimed to determine the health status of the animals. Zooarchaeological and stable isotopic analysis (SIA) have revealed some of the criteria used for animal selection (Dufour et al. Reference Dufour, Goepfert, Neün, Prieto and Verano2020). Only young animals were chosen, and most had a brown coat, although some were completely beige (Goepfert et al. Reference Goepfert, Elise Dufour and Verano2020; Prieto et al. Reference Prieto, Verano, Goepfert, Kennett, Quilter, Leblanc, Fehren-Schmitz, Forst, Lund, Dement, Dufour, Tombret, Calmon, Gadison and Tschinkel2019). The relationship between the stable isotopic composition of an animal and that of its diet and environment allows the reconstruction of an animal's life history (e.g., DeNiro and Epstein Reference DeNiro and Epstein1978; Schoeninger and DeNiro Reference Schoeninger and DeNiro1984; Zazzo et al. Reference Zazzo, Balasse, Passey, Moloney, Monahan and Schmidt2010). This approach has been used in the Andean region to study camelid herding and management strategies (e.g., Dufour et al. Reference Dufour, Goepfert, Léon, Chauchat, Jordan and Sánchez2014; Samec et al. Reference Samec, Yacobaccio and Panarello2018; Szpak et al. Reference Szpak, Millaire, White and Longstaffe2014) and has suggested that the HLL camelid offerings were from several different herds, raised in the Lowlands. It does not support the existence of specific herds dedicated to ritual purposes (Dufour et al. Reference Dufour, Goepfert, Neün, Prieto and Verano2020).
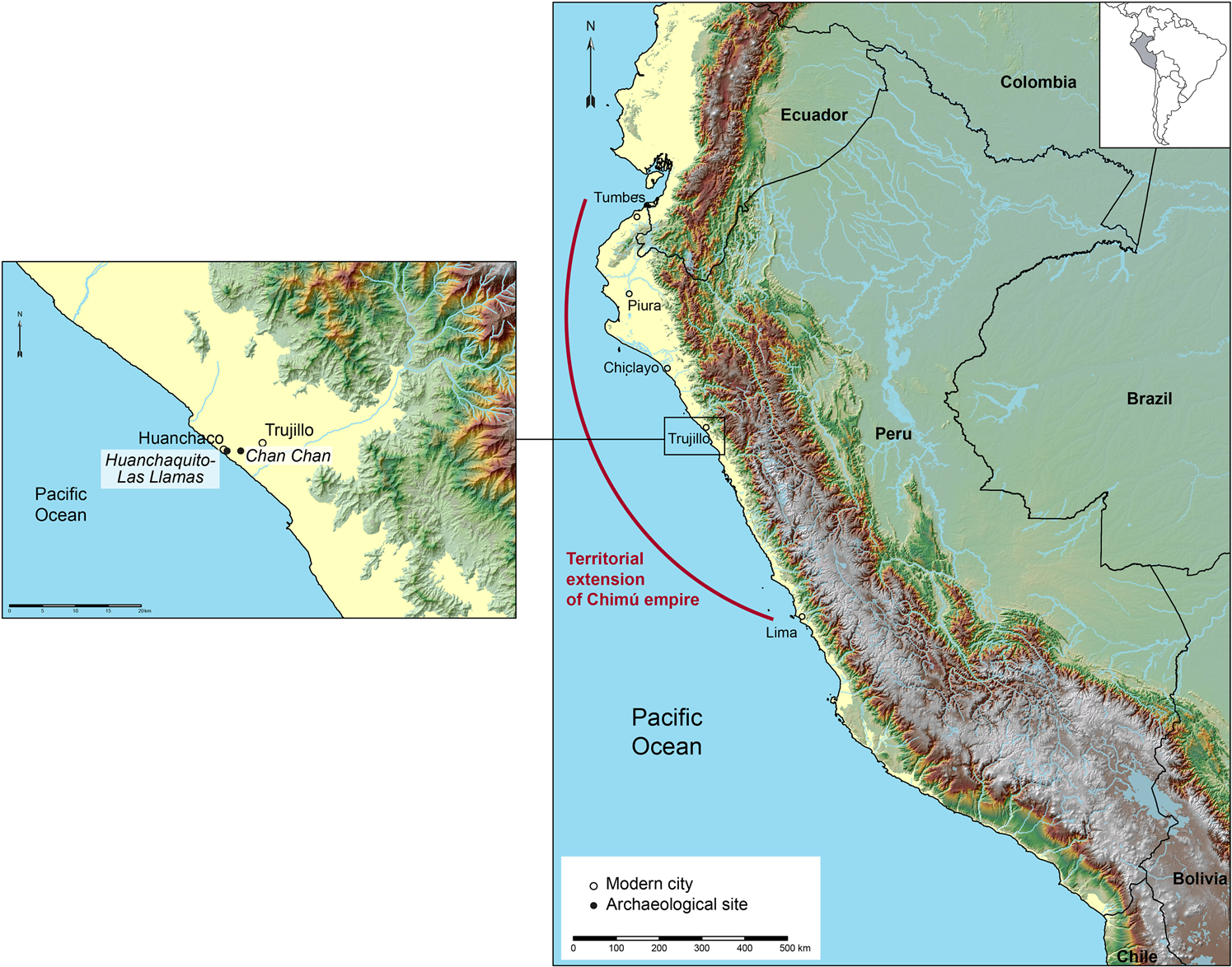
Figure 1. Location of Huanchaquito-Las Llamas (map by Jean-François Cuénot and Nicolas Goepfert). (Color online)
The SIA of bone collagen represents the weighted average of dietary intake. Over their lives, the HLL camelids consumed diverse wild and cultivated plants in variable proportions, depending on the herders’ practices and choices (Dufour et al. Reference Dufour, Goepfert, Neün, Prieto and Verano2020). Diet variability was large, but in all cases, the proportion of C4 plants—plants with a photosynthesis pathway that minimizes photorespiration and that are thus adapted to arid conditions and are mostly present in tropical regions—was moderate to large compared with that of C3 plants. However, SIA can only point to the exploitation of a habitat type, such as lomas (areas of fog-watered vegetation), or quantify the consumption of food categories such as plants using C3 and C4 photosynthesis pathways; thus, it can only be suggestive of specific plant species. Maize (Zea mays), the principal cultivated C4 plant, is thought to represent a significant dietary input and could explain the very high C4 plant isotopic signature of some of the individuals. However, grama salada (Distichlis spicata), a wild, salt-tolerant C4 grass, could have also constituted a sustainable grazing resource because it is widespread. Given that the SIA of bone collagen represents a dietary average over a long period of time, it cannot track short-term dietary changes. Ethnographic data may be a good source of information about short-term changes in diet before sacrifice, but none are available in this case. Therefore, to address whether a special (perhaps ritually significant) meal was given to the camelids before their sacrifice, we studied starch grains, derived principally from the associated gut contents of the sacrificed camelids.
Starch Grain Analysis
Starch grains are the energy-storage units of plants. Composed of two different glucose chains, they are tiny particles, ranging in size from 1 to 100 μm. They are stored in various organs in a green plant; however, dense amounts of starch are often concentrated in seeds, tubers, and fruits (Haslam Reference Haslam2004). In archaeology, starch has been successfully extracted from tools and ceramic vessels (Cagnato and Ponce Reference Cagnato and Ponce2017), organic containers (Duncan et al. Reference Duncan, Pearsall and Benfer2009), dental calculus (Piperno and Dillehay Reference Piperno and Dillehay2008), coprolites (Vinton et al. Reference Vinton, Perry, Reinhard, Santoro and Teixeira-Santos2009), and sediments (Therin et al. Reference Therin, Fullagar, Torrence, Gosden and Hather1999). The presence of starch has played an important role in tropical areas, where macrobotanical remains may not be preserved as often or as well as they are in other parts of the world (Dickau et al. Reference Dickau, Ranere and Cooke2007). The long-term survival of starch grains makes them especially useful for better identifying plant parts that do not preserve well and are thus underrepresented in the macrobotanical archive, including underground storage organs (i.e., tubers and rhizomes; Piperno and Holst Reference Piperno and Holst1998). These elements are useful because they tend to be ubiquitous in plants, they have a series of morphological features that often allow them to be identified to the taxon (genus or even species) level, and their size and shape can be highly diagnostic (Gott et al. Reference Gott, Barton, Samuel, Torrence, Torrence and Barton2006). The recovery, and identification, of starch grains in different archaeological contexts in South America, and more particularly in the Andean region, is not novel (see Duncan et al. Reference Duncan, Pearsall and Benfer2009; Ikehara et al. Reference Ikehara, Paipay and Shibata2013; Piperno and Dillehay Reference Piperno and Dillehay2008; Vinton et al. Reference Vinton, Perry, Reinhard, Santoro and Teixeira-Santos2009). Although microbotanical research has been carried out on camelid feces remains from Peru and Argentina (see Bonavia Reference Bonavia2008a; Caruso Fermé et al. Reference Caruso Fermé, Velázquez, Tosto, Yagueddú, Burry and Civalero2018; Jones Reference Jones1990; Jones and Bonavia Reference Jones and Bonavia1992; Korstanje Reference Korstanje and O'Connor2005; Mosca Torres et al. Reference Mosca Torres, Urquiza and Aschero2018), only one study revealed the presence of starch grains, belonging to Typha sp. (Bonavia Reference Bonavia2008a:403). Our study is unique because, through the recovery and identification of starch grains in gut remains, it identified multiple species and thus provides some insights into the final diet of these camelids before their sacrifice. We compare the short-term dietary history as recorded by starch grain results with the long-term dietary history as recorded by SIA to distinguish between the regular and the special diet of these camelids.
Materials and Methods
The site of Huanchaquito-Las Llamas is located in the Moche Valley, on a dune 300 m from the shoreline. It is 1.5 km northwest of Chan Chan, the ancient Chimú capital, and 4 km from the modern city of Trujillo, in the La Libertad region. It yielded 206 sacrificed domestic camelid individuals, probably llamas, and 140 human individuals, of whom 137 were children and 3 were adults (Prieto et al. Reference Prieto, Verano, Goepfert, Kennett, Quilter, Leblanc, Fehren-Schmitz, Forst, Lund, Dement, Dufour, Tombret, Calmon, Gadison and Tschinkel2019). The camelids were, for the most part, in a good state of preservation, with their fleece still present (Figure 2a), their natural mummification made possible by the dry conditions of the Peruvian coast (Collin Delavaud Reference Collin Delavaud1984). For some of the individuals, their gut contents (Figure 2b, c), feces (Figure 2d), and ropes used to handle them were also still present. The sacrificial event has been radiocarbon dated to between AD 1400 and 1450 (Prieto et al. Reference Prieto, Verano, Goepfert, Kennett, Quilter, Leblanc, Fehren-Schmitz, Forst, Lund, Dement, Dufour, Tombret, Calmon, Gadison and Tschinkel2019). At that time, the Chimú empire was at its apogee, having reached its maximum territorial extension—from northern Peru (the area bordering Ecuador) to the central coast (modern-day Lima). Major sites controlled this vast territory, which extended 1,000 km north to south, administering local populations and their intensive agriculture (Moore and Mackey Reference Moore, Mackey, Silverman and Isbell2008).

Figure 2. Camelid, gut, and feces remains from Huanchaquito-Las Llamas: (a) camelid (CA-07); (b, c) gut contents at different levels of digestion (CA-20 and CA-47); and (d) feces (CA-05). Photos by Nicolas Goepfert. (Color online)
We collected gut contents and feces from 52 individuals in the field over the course of three excavation seasons (2011, 2014, and 2016). The samples were exported with the permission of the Ministerio de Cultura of Peru (N°050-2016-VMPCIC-MC). We carried out starch grain analysis on the gut contents of 11 camelids, ranging from 6 months to 1.5 years in age (based on observations of tooth eruption and tooth wear; Table 1, Figure 2b, c), and on the feces of one of these individuals (CA-05). This corpus represents 21% of the available gut and fecal samples. The other 79% of the samples had already been used for paleoparasitological (Le Bailly et al. Reference Le Bailly, Goepfert, Prieto, Verano and Dufour2020) and phytolith analysis (in an ongoing study), both of which rendered them unusable for subsequent starch grain analysis.
Table 1. Starch Grain Results Obtained from HLL Camelid Gut Contents and Feces.

a See text for discussion on identification.
b Fecal sample.
The samples were treated at the MSH Mondes following previously outlined laboratory extraction methods (Cagnato and Ponce Reference Cagnato and Ponce2017). Before analysis, all the laboratory consumables were sterilized using bleach. We concentrated 1 g of the gut remains from each sample by centrifugation. To dissolve any limescale in the sample, we added 10 ml of 0.1% ethylenediaminetetraacetic acid (EDTA) and then rinsed it with distilled water. We then added hydrogen peroxide (5.75%) to ensure that nonstarch organics would be destroyed and to aid in breaking up the sediment. We performed heavy-liquid flotation using sodium polytungstate (SPT), which enables a centrifuge to separate the starch from the rest of the sediment by using a specific gravity that ensures that the starch floats or remains suspended. Fresh starch typically has a density of 1.5g/ml, and therefore the specific gravity of the SPT was set at 1.7g/ml. We did supernatant extraction two more times. Once the solution was clear, we transferred it to a clean slide. To mount the samples, we used a 50% water and glycerin solution, which slows down the drying of the sample and makes it easier to rotate the starch grains. Finally, we placed a coverslip to seal the sample.
We observed the starch grains under a cross-polarized microscope (Nikon Eclipse E600 Pol) and took photographs and measurements using the software NIS-Elements. Starch grains were identified to genus and when possible to species, according to qualitative and quantitative criteria, which included the presence of the distinct extinction cross (a birefringent pattern visible under polarized light); the size of the grain; the angularity of the facets; and whether lamellae, fissures, or the hilum was visible. We took photographs of every starch grain at 600× in both cross-polarized light and transmitted light. We then compared the starch grains with a reference collection (Supplemental Table 1), located at MSH Mondes, that includes wild plants found along the coast and domesticated species previously reported in archaeobotanical contexts in the Andean region.
Of the 11 camelids we sampled, eight had already been sampled for SIA (Dufour et al. Reference Dufour, Goepfert, Neün, Prieto and Verano2020). For the other three (CA-151, CA-171, CA-200; Supplemental Table 2), we prepared a fragment of bone (phalanx) and extracted collagen (gelatin) at the sclerochronology lab of the Muséum National d'Histoire Naturelle (MNHN), Paris, as described by Dufour and colleagues (Reference Dufour, Goepfert, Neün, Prieto and Verano2020). For δ13C and δ15N and C and N content analysis, between 300 and 500 μg of gelatin were combusted using an Elemental Analyser Flash 2000 coupled with a Delta V Advantage (Thermo Scientific) isotope ratio mass spectrometer, at the Service de Spectrométrie de Masse Isotopique du Muséum (SSMM) of the MNHN. We estimated the mass spectrometer analytical error to be 0.164‰ for δ13C and 0.083‰ for δ15N, based on replicate analysis of an internal standard (alanine; accepted δ13C = −22.16‰ and δ15N = +0.59‰) calibrated against the international standard IAEA 600. Analysis of sample CA-200 was repeated three times to test for intraindividual variability and analytical reproducibility and provided standard deviations of 0.060‰ and 0.032‰ for δ13C and δ15N, respectively. We used well-defined criteria to test for the quality of preservation of collagen: yield (wt %), C and N content (%), and the atomic C:N ratio (Ambrose Reference Ambrose1990; DeNiro Reference DeNiro1985; Dobberstein et al. Reference Dobberstein, Collins, Craig, Taylor, Penkman and Ritz-Timme2009).
We estimated the relative contributions of C3 and C4 plants in the diet of the 11 camelids using a single Bayesian mixing model in the SIAR package (Parnell and Jackson Reference Parnell and Jackson2013). Values of plants sampled in northern Peru by Szpak and colleagues (Reference Szpak, Millaire, White and Longstaffe2014) were used as source values. We used a correction of ~1.5‰ for modern source values to account for the Suess effect (Marino and McElroy Reference Marino and McElroy1991) and a collagen–diet spacing value of +3.6 ± 1.7‰ for bone collagen (Szpak et al. Reference Szpak, Millaire, White and Longstaffe2014).
Results
In total, we recovered 51 starch grains from the 12 starch grain samples studied (Table 1). We also included in the final count starch grains (n = 14, plus a large cluster) that we found in 10 of the 33 gut samples tested for phytoliths (ongoing study). These latter grains could not be viewed in three dimensions, and therefore their identification remains tentative, but the breakdown is as follows: cf. maize (n = 5), cf. Prosopis sp. (algarrobo; n = 1), a cluster of lenticular starch grains (not individually counted), and unidentified/damaged grains (n = 8).
The most numerous grains identifiable to genus or species belong to maize (n = 14) and were present in 7 of the 11 camelids studied. The grains are simple, with an open hilum, ranging from round to more polyhedral, and they have a continuous double border (Figure 3a–d). On average, the grains in the camelid samples measure 16.5 μm in width.

Figure 3. Starch grains viewed under transmitted and cross-polarized light. Maize starch grains from individuals CA-200 (a, b) and CA-05 (c, d). Lenticular starch grains from individuals CA-200 (e, g) and CA-171 (h, i). (Color online)
A total of seven grains closely resembling those produced by chili peppers (Capsicum sp.), locally known as “aji,” were identified from four camelids (Figure 3e–g). Moreover, in the sample of camelid CA-171, we found a large mass of fused lenticular grains (Figure 3h, i). It remains unclear whether these grains belong to chili peppers or to a member of the Triticeae tribe, which produces similar grains to those of the chili pepper. Notably, starch grains produced by chili peppers are lenticular when viewed in plan view (see Figure 3a, b); in side view, they have a line that extends parallel to the long axis of the grain (see Figure 3g). They measure between 13 and 45 μm in length (Perry et al. Reference Perry, Dickau, Zarrillo, Holst, Pearsall, Piperno, Berman, Cooke, Rademaker, Ranere, Raymond, Sandweiss, Scaramelli, Tarble and Zeidler2007). Only two genera in the Triticeae tribe are known to grow wild in the region: Hordeum and Elymus (Mostacero León et al. Reference Mostacero León, Coico and Torres2009). For the former genus, H. muticum is found in western South America, and H. pubiflorum is found in Peru (Bothmer et al. Reference Bothmer, Sato, Komatsuda, Yasuda, Fischbeck, von Bothmer, van Hintum, Knüpffer and Sato2003); for the latter genus, three species are known from our region of interest: E. angulatus, E. cordilleranus, and E. hitchcockii. We do know that species in the genus Elymus tend to produce smaller starch grains (see Yang and Perry Reference Yang and Perry2013), and in Peru, they tend to grow at higher altitudes (Mostacero León et al. Reference Mostacero León, Coico and Torres2009). Therefore, we can eliminate Elymus as a potential candidate. The size range of the starch grains in H. muticum and H. pubiflorum is currently unknown. The non-native wild Hordeum species (H. spontaneum, H. marinum) produce starch grains that measure between 6 and 26 μm (Piperno et al. Reference Piperno, Weiss, Holst and Nadel2004), whereas domesticated barley (H. vulgare) produces grains that measure between 8 and 25 μm (Henry et al. Reference Henry, Hudson and Piperno2009). However, an identification as Hordeum is problematic because the grains from HLL seem to lack one characteristic that is well known for Hordeum starch grains: the presence of crater-like depressions on the surface.
We found additional taxa in smaller quantities. Five grains resembling those produced by algarrobo (Figure 4a, b), measuring on average 25 μm, were found in four of the camelids (CA-134, 136, 151, 170). Three grains of manioc were recovered from the gut sample of one camelid (CA-68; Figure 4c, d). The grains are bell shaped, with one to three basal facets, and measure between 6 and 12.50 μm in length. They are on the small end of the spectrum but still fall within published ranges (see Piperno and Holst Reference Piperno and Holst1998). We found one starch grain measuring 20 μm in length belonging to the Fabaceae family, in camelid CA-136 (Figure 4e, f). Due to damage, it was not possible to identify it further, but it could belong to the domesticated bean (Phaseolus vulgaris) or the lima bean (P. lunatus).
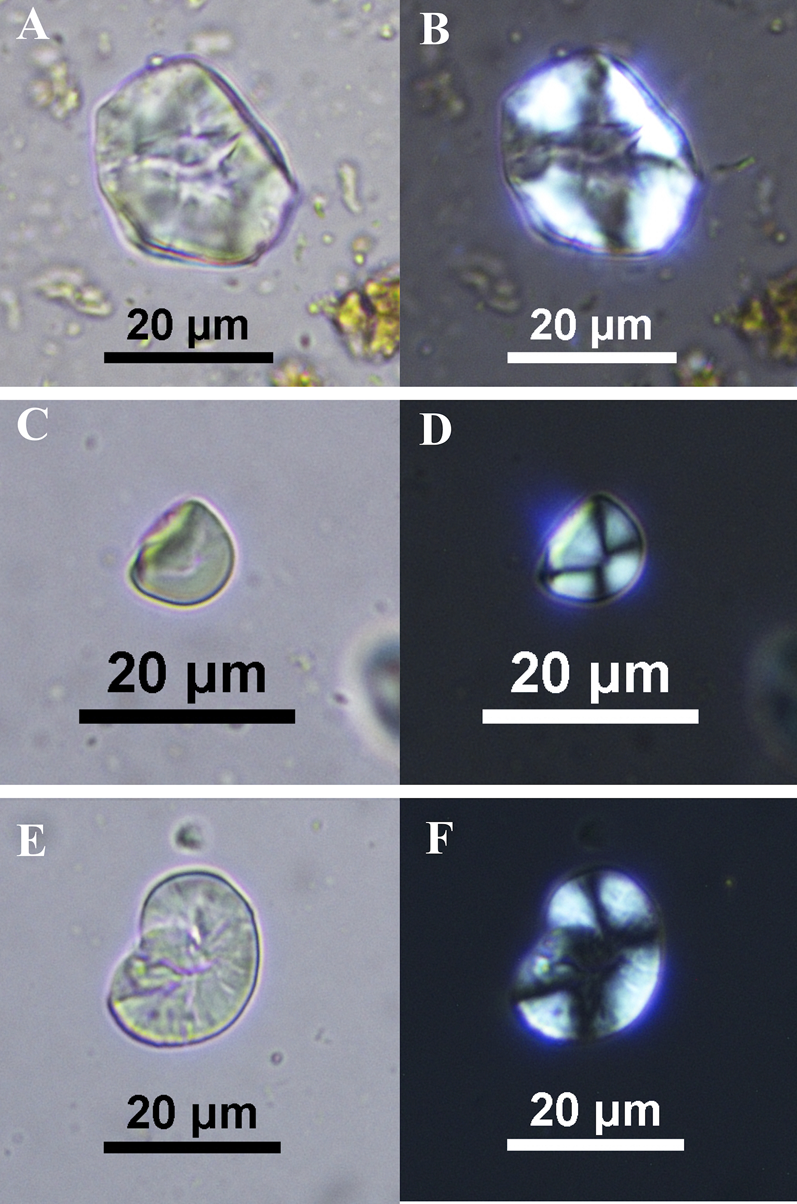
Figure 4. Various starch grains recovered from the gut samples, viewed under transmitted and cross-polarized light: (a, b) algarrobo (CA-170); (c, d) manioc (CA-68); and (e, f) bean family (CA-136). (Color online)
We also found a total of 21 grains that could not be identified further, mainly due to damage, perhaps caused by the effects of mastication and digestion in the camelid gut. Some elements, such as the exploded hila seen on three starch grains (Figure 5a–c; Babot Reference Babot, Hart and Wallis2003; Cagnato Reference Cagnato2019), suggest that the camelids may have been fed cooked foods. In another camelid, three fused starch grains clearly are in the process of gelatinization (Figure 5d, e). One mass of starch grains recovered from CA-151 resembles the elements commonly seen when starchy foods are exposed to heat (Figure 5f). In some cases, the extinction cross is less clear or absent (Figure 6a–h). In two camelids, the starch grains have small pits on their surface (Figure 6i–l). The cause of this damage is unclear.

Figure 5. Damaged starch grains: (a) CA-134; (b) CA-178; (c) CA-11; (d, e) CA-200; and (f) CA-151. (Color online)

Figure 6. Damaged starch grains, viewed under transmitted and cross-polarized light: (a, b) CA-194; (c, d) CA-136; (e, f) CA-05f; (g, h) CA-151; (i, j) CA-151; and (k, l) CA-136. (Color online)
In addition, 50% of the samples provided round pollen grains having a single pore and ring (annulus) and measuring on average 65 μm wide. They belong to the Poaceae family, and we believe these are good candidates for maize pollen (Figure 7). The presence of maize pollen would be consistent with the predominance of starch grains that are likely also of this taxon in the final meal. Pollen grains of other morphotypes were present but in smaller quantities. They were not identified further in this study.

Figure 7. Probable maize pollen grain (CA-05). (Color online)
CA-151, CA-171, and CA-200, as did all other individuals selected for SIA, met the criteria defined for assessing the quality of preservation of bone collagen (Supplemental Table 2), confirming the very good state of preservation at HLL. Their δ13C and δ15N values varied from −13.9‰ to −11.1‰ and from +6.9‰ to +8.6‰, thus falling within the range of values measured previously for 82 other individuals from this site (Dufour et al. Reference Dufour, Goepfert, Neün, Prieto and Verano2020). C4 plants dominate the diet of the 11 individuals sampled for starch grain analysis, with a mean contribution of C4 and C3 plants estimated to be 80.1% (95% of the Bayesian credible interval: 72.3%–88.3%) and 19.9% (95% of the Bayesian credible interval: 11.7%–27.7%), respectively (Figure 8). There is no correlation between the mean contribution of C4 plants to the diet and the presence of maize starch grains in the feces or gut contents.

Figure 8. Plot of bone collagen δ13C values (‰, VPDB) and estimated proportion of C4 in the diet for the 11 domestic camelids from Huanchaquito-Las Llamas. The proportion of C4 in the diet was estimated using bone collagen δ13C values as defined in the text. The study of gut contents and feces provided maize starch grains for seven camelids (full circles) and no starch grains for four camelids (open circles).
Discussion
Long-Term Camelid Diet at Huanchaquito-Las Llamas
The gut contents of 11 HLL camelids and the feces of one camelid yielded 51 starch grains belonging to five taxa. Of these grains, 41% were not identifiable, and therefore it is possible that additional taxa may have been consumed. Making sense of these data is not straightforward because of the lack of comparative data. Today, camelids do not live on the coast of Peru, except for a modern herd raised at a low altitude at the Tinajones reservoir in the province of Lambayeque, where two of the authors (ED and NG) were able to observe some of its feeding habits. However, these animals have been subject to significant ecological adaptations. Historical data concerning the diet of camelids are not abundant, but those that do exist provide some interesting insights. Moreover, there are archaeobotanical data available, as well as SIA records that specifically targeted camelid dietary reconstruction.
In the present study, maize was the most ubiquitous taxon, its starch grains being present in 7 of 11 camelids. This is in accordance with the diet reconstructed through SIA, which indicates that these individuals consumed a large proportion of C4 plants over their lifetime (see Supplemental Table 2 and Figure 8). Maize has been widely reported in various forms in Andean archaeology (Bonavia Reference Bonavia2008b; Haas et al. Reference Haas, Creamer, Mesía, Goldstein, Reinhard and Rodríguez2013). It was present as early as 6700 BP on the northern coast of Peru (Grobman et al. Reference Grobman, Bonavia, Dillehay, Piperno, Iriarte and Holst2012) and was already a staple in this part of Peru by the fifteenth century AD (Masur et al. Reference Masur, Millaire and Blake2018). The Chimú developed intensive agriculture using an extensive hydraulic system (Farrington and Park Reference Farrington and Park1978; Ortloff Reference Ortloff, Lechtman and Soldi1981), and maize was an essential component of their diet and culture. Historical sources indicate that camelids ate maize when it was given to them, but that otherwise they would graze on whatever they could find. A drawing by chronicler Guamán Poma de Ayala (Reference Guamán Poma de Ayala2005 [1615]:919) shows a llama in a field eating maize, and Squier (Reference Squier1877) witnessed corralled llamas being fed maize kernels in the highlands. This practice seemingly continues, being observed among current llama breeders (Shimada and Shimada Reference Shimada and Shimada1985). Archaeologically, maize has been reported in various forms from camelid coprolites: pollen from llama excrements at PV35-4, a Middle Horizon site (Jones Reference Jones1990), and maize stomata and other diagnostic features from feces at Huaca 1, a LIP site (Bonavia Reference Bonavia2008a). Our starch grain data clearly indicate that maize kernels were consumed by the camelids, because it was their guts that contained the starch grains.
The SIA of archaeological remains has suggested that there was long-term foddering of animals with maize in the Lowlands during the Mochica period at the Uhle Platform (Dufour et al. Reference Dufour, Goepfert, Léon, Chauchat, Jordan and Sánchez2014) and at sites dated from the Early Intermediate period (EIP) to the LIP in the Virú and Chicama Valleys (Santana-Sagredo et al. Reference Santana-Sagredo, Dufour, Goepfert, Zazzo, Jordán and Sánchez2020; Szpak et al. Reference Szpak, Millaire, Chapdelaine, White and Longstaffe2020). Foddering could have been a method used by the lowland herders to compensate for the limited productivity of coastal habitats, and, more widely, it may have been the usual camelid management strategy in the Andes. It was observed at the highland site of Conchopata (2,700 m asl), where maize was an important staple in the economy of the Wari polity (AD 550–1000; Finucane et al. Reference Finucane, Agurto and Isbell2006). There are thus some converging indications that maize could have been a common element in the camelid diet, but earlier records were unable to document which part of the plant was consumed. The consumption of maize kernels indicated by our study is significant because of their high caloric value compared with maize byproducts, such as stalks.
C4 plants other than maize may have contributed to the high δ13C values of the HLL camelids, including cultivated Amaranthus sp. and wild grasses and sedges. Grama salada, which is part of the plant community type called Gramadales, has a wide distribution in areas close to the sea and on dunes (Mostacero León et al. Reference Mostacero León, Coico and Torres2009). According to Thornton and colleagues (Reference Thornton, DeFrance, Krigbaum and Williams2011), this salt-tolerant grass, as well as dropseed (Sporobolus rigens), is consumed by both wild and domestic camelids. However, the consumption of these non-maize plants does not appear to be sufficient to support the daily feeding of large herds and to explain a C4 contribution as high as the 70% observed for the sacrificed animals at HLL.
Starch grains of algarrobo were recovered from four of the camelids from HLL and therefore represent the second most frequent plant recovered from the samples. Starch grains are only found in the mesocarp (the fleshy pulp surrounding the seed) of the algarrobo pod; therefore, the HLL camelids clearly consumed its pods. Algarrobo seeds and other tissues were reported from llama feces from archaeological contexts previously (Bermeo Reference Bermeo and Goepfert2017; Bonavia Reference Bonavia2008a). In the province of Lambayeque, endocarps and seeds were found in llama dung dating to the LIP (Bonavia Reference Bonavia2008a; Shimada and Shimada Reference Shimada and Shimada1985). We observed a few modern adult llamas from the Tinajones herd feeding on algarrobo pods and leaves, but because they were only from one herd, it is difficult to know whether this habit could be common. The practice of foddering livestock was mentioned by Capparelli and Lema (Reference Capparelli and Lema2011:83). Algarrobo may have been an important staple, because it is not solely recorded for camelids: it was also found in the gut contents of human mummies (Holden and Núñez Reference Holden and Núñez1993); in human coprolites in the Ica Valley of Peru (Beresford-Jones et al. Reference Beresford-Jones, Whaley, Ledesma and Cadwallader2011); and in preceramic contexts at Buena Vista, notably as residue inside organic containers recovered from feasting contexts dated to about 2200 cal BC (Duncan et al. Reference Duncan, Pearsall and Benfer2009).
The Unexpected Ritual Foods
Although maize and algarrobo may be easily recognized as regular staple foods for camelids, the presence of manioc and chili peppers was not intuitive. Evidence for manioc has been recovered from various places and from different time periods along the coast of Peru in the form of dried roots, starch grains, and phytoliths (Duncan et al. Reference Duncan, Pearsall and Benfer2009; Ikehara et al. Reference Ikehara, Paipay and Shibata2013; Prieto Reference Prieto, Prieto and Sandweiss2020; Ugent et al. Reference Ugent, Pozorski and Pozorski1986), and it is depicted rather ubiquitously on ceramics from the Chimú period. Manioc tubers can be ground into flour to prepare bread and can be made into a fermented beverage (Duncan et al. Reference Duncan, Pearsall and Benfer2009). The bitter and the sweet varieties are the same species; their only difference is their differing concentrations of cyanogenic glucosides (Isendahl Reference Isendahl2011). It remains unclear whether the sweet, the bitter, or both varieties were consumed along the coast of Peru in prehispanic times (Prieto Reference Prieto, Prieto and Sandweiss2020; Ugent et al. Reference Ugent, Pozorski and Pozorski1986). Yet if camelids ingested the bitter variety raw, it could have resulted in cyanide intoxication, which can cause dizziness, vomiting, and even death (Isendahl Reference Isendahl2011). Moreover, because camelids “do not uproot vegetation with their feet, and never destroy stems and roots” (Bonavia Reference Bonavia2008a:399), it seems unlikely they dug up the manioc root while the herd was kept in pens or fields. It is more probable that sweet or bitter manioc was intentionally given to them by Chimú herders before their ritual killing.
Finally, we present the first tentative evidence of the ingestion of chili peppers by camelids. This spicy plant, considered sacred by the Inca, has been found in both domestic and ritual contexts across the Andean region (Beresford-Jones et al. Reference Beresford-Jones, Whaley, Ledesma and Cadwallader2011; Chiou et al. Reference Chiou, Hastorf, Bonavia and Dillehay2014; Pickersgill Reference Pickersgill1969). As with manioc, chili peppers are depicted on Chimú ceramics. However, the identification of their lenticular starch grains is complicated because of the similarities between the starch grains produced by the Triticeae tribe grasses and those produced by chili peppers. Although the little surface craters/pits are absent on the starch grains in the archaeological samples (seemingly indicating that these are not likely to be grass starches), we also need to consider the effects that mastication and digestive processes may have had on these microbotanical remains. Although it would seem more plausible, given the ritual circumstances, that the animals would have been fed or would have naturally grazed on wild Hordeum, it is not impossible to imagine that special foods or unusual food preparations, including chili peppers, may have been given or fed to them before their death.
Beans were of course consumed by humans. In northern Peru, starch grains of the genus Phaseolus are reported from dental calculus recovered from an early and middle Holocene human settlement in the Ñanchoc Valley (Piperno and Dillehay Reference Piperno and Dillehay2008), and bean cotyledons and pods were also documented from the Virú Valley (Masur et al. Reference Masur, Millaire and Blake2018). The only evidence to date for the Fabaceae family is the presence of Vicia sp. pollen in one llama coprolite sample from a preceramic site on the north-central coast of Peru (Jones and Bonavia Reference Jones and Bonavia1992). There are no known historical accounts of camelids consuming beans in Peru and more generally in the Andes.
Consumption of Cooked Food
Another interesting result is evidence for the ingestion of different preparations of food, including possibly cooked foods. The starch grains recovered from the samples generally retained their shape, even after mastication, ingestion, and digestion. It is known that starch digestion is complex and that a range of factors influence this process (Tester et al. Reference Tester, Qi and Karkalas2006). Uncooked starch will break down more slowly than cooked starch (Copeland and Hardy Reference Copeland and Hardy2018), because raw starch resists digestion by the α-amylase enzyme present in the mouth and gut (Tester et al. Reference Tester, Qi and Karkalas2006). The presence of relatively unaltered grains in the samples may suggest that some of the foods eaten by the camelids were raw, although it is also possible that these seemingly intact grains resisted cooking processes (Crowther Reference Crowther2012). The presence of a gelatinized starch mass and other diagnostic damage that is often seen in cooked plant foods (Babot Reference Babot, Hart and Wallis2003; Vinton et al. Reference Vinton, Perry, Reinhard, Santoro and Teixeira-Santos2009) are more secure indicators that cooked foods were also given to the camelids. Providing cooked foods to the camelids could have been a special practice because of the ritual nature of the event. Another issue that should be considered is whether these foods were consumed as solids or liquids. If the animals were fed some of these plants, we can imagine that they were forced to drink, rather than to eat. The discovery of ceramic vessels, together with wooden paddles, at HLL may be related to the consumption of maize beer, or chicha, during the sacrifice (Prieto et al. Reference Prieto, Verano, Goepfert, Kennett, Quilter, Leblanc, Fehren-Schmitz, Forst, Lund, Dement, Dufour, Tombret, Calmon, Gadison and Tschinkel2019), and the presence of maize starch grains in the camelids’ guts suggests that it is possible that the camelids were given this drink. Chicha was important in both the domestic and the ritual sphere (Jennings and Bowser Reference Jennings and Bowser2009). Since prehispanic times, chicha was prepared not only for daily consumption but also as a key element in ritual and political events (Hayashida Reference Hayashida2008). Different types of evidence exist to support the production of chicha (see Hayashida Reference Hayashida2008; Vásquez et al. Reference Vásquez, Alvárez, Rosales, Fraile and Dorado2018), including its flavoring with a variety of fruits (Prieto Reference Prieto, Zori and Johnson2011). The recovery of maize starch grains from feasting vessels from a coastal Peruvian site has been argued to be indicative of events that served to integrate the community through the increased consumption of maize (Ikehara et al. Reference Ikehara, Paipay and Shibata2013). None of the starch grains recovered from the HLL samples exhibit the damage seen on the starches of maize that had been subjected to experimental fermentation by Vinton and colleagues (Reference Vinton, Perry, Reinhard, Santoro and Teixeira-Santos2009). However, some of the starch grains (Figure 6) do exhibit peculiar damage that has not been seen previously by the first author and that could be the result of some special preparation of the maize given to the camelids.
A Final, Special Meal for the Huanchaquito-Las Llamas Camelids
The sacrifice at HLL necessitated the provisioning of a large number of animals. High variability in both δ13C and δ15N values shows that these young, sacrificed camelids were raised locally on the coast or in the middle valleys and that they most likely originated from different herds that were segregated from each other (Dufour et al. Reference Dufour, Goepfert, Neün, Prieto and Verano2020; Prieto et al. Reference Prieto, Verano, Goepfert, Kennett, Quilter, Leblanc, Fehren-Schmitz, Forst, Lund, Dement, Dufour, Tombret, Calmon, Gadison and Tschinkel2019). The ritual practices and possible management by the Chimú officiants of these animals before their sacrifice may have included keeping them together for a short period and feeding them a special diet. Although collagen analysis cannot be used to infer short-term dietary changes, microbotanical analysis can give us a snapshot of the last meal: the remains of the last 48 hours should be represented in the gut. Alpacas, for example, retain their food in their digestive tract for about 50 hours (Sánchez Reyes Reference Sánchez Reyes2004). Based on what we know about their diet throughout their life (by SIA of their bone collagen) and about the last foods they ate (by studying the starch grains present in their gut and feces) we were able to establish that, shortly before their death, the camelids consumed a broader range of plants than expected. The animals may have been corralled some hours or days before being sacrificed, as evidenced by the remnants of special food that consisted of manioc, beans, and possibly chili peppers and some cooked foods, as well as of foods that were part of their usual diet. Shortly before death, most of the young camelids (7 of 11) consumed maize, a food that they had likely fed on regularly, as evidenced by the high estimated C4 plant contribution. A few also ate algarrobo, which again seems to have been a normal practice when we consider the archaeobotanical and historical record. None of the plants identified in the samples appear to have been imported, because all these taxa could have been grown locally. Chimú society expanded throughout the Moche Valley to take advantage of its fertile fields for growing maize, chili peppers, and coca, as well as the naturally growing bosque seco (algarrobo, acacia). The bosque seco is today a highly threatened forest ecosystem, but, as noted by Rostworowski (Reference Rostworowski1981), it was more prevalent in the past. Therefore, these plants were likely accessible by the inhabitants of the coast.
How did the camelids gain access to these foods before their sacrifice? As we note earlier, one option would be for herders to bring maize—including kernels—and other foods directly to the animals selected for sacrifice. Alternatively, the presence of maize pollen grains in the camelid gut contents suggests that these animals were likely present in or in the vicinity of maize fields or that they were left to forage on maize cobs or graze on the stalks and leaves. Regardless of how the pollen arrived in the camelid guts, occasional foraging in the fields is not enough to explain the high C4 proportions as determined by SIA. Our combined approach allows us to propose with greater certainty the possibility that the camelids were foddered during their lifetime and received special foods, even ritually significant foods, before their sacrifice. Similarly, it could contribute much to the knowledge of the ritual sequence accompanying human sacrifices and to that of more common dietary practices.
Conclusions
Because this is the first and, therefore, an exploratory study carried out on starch grains from desiccated camelid gut contents, it was initially unclear whether these microbotanical remains would be preserved and in what condition. Based on our results, we can confirm that starch grain analysis can be successfully carried out on gut contents to determine what foods were part of a camelid's final meal. Combining these data with other types of analyses—the study of phytoliths and pollen—will provide additional important information regarding the identification of consumed taxa.
The very good preservation of the faunal remains and the interdisciplinary study carried out on this particular archaeological collection show the full potential of this type of approach. By combining the identification of starch grains with SIA, we were able to elucidate the diet of the sacrificed camelids over both the long and short term. The major presence of maize starch grains is consistent with the isotopic results and suggests the importance of this plant to the animals’ diet throughout their life and until their final diet. We document both how the Chimús managed their herds and that they also reserved special treatment for those animals destined for sacrifice. Probably gathered a few hours or days before the sacrificial act, these camelids, which came from different herds, benefited from a special diet, with the addition of unusual foods that included manioc, chili peppers, and beans, some of which may have been cooked as part of a prepared meal. Although we do not know the specific reasons why these plants or meals were chosen and given to the camelids, we could argue that the presence of these particular plants suggests their importance in the ritual.
There is no doubt that the analysis of gut remains shows great potential to provide new insights into camelid management and ritual practices. However, such analysis can also be applied to studying human diets in ritual contexts. In the case of HLL, we were able to establish the ritual sequence a few hours or days before the camelid sacrifice. The use of animals in ancient rituals in the Andes and across the American continent is well attested; in some cases, these animals may even have been held in captivity and fed a special diet before the ritual event (Emery Reference Emery, O'Day, Van Neer and Ervynck2004; Sugiyama et al. Reference Sugiyama, Somerville and Schoeninger2015). Future applications of starch grain analysis in the study of well-preserved animal remains may prove fruitful to reconstruct management practices and ancient rituals and to demonstrate whether standardized practices existed in the past. Similarly, such analysis could contribute much to our knowledge about the ritual sequence accompanying human sacrifices or, indeed, of more mundane dietary practices.
Acknowledgments
This work was supported by the ANR CAMELANDES under Grant ANR-15-CE27-0002 and AASPE (UMR 7209 MNHN-CNRS). We thank the ArchéoScopie department at the MSH Mondes in Nanterre (France) for providing access to the imaging equipment. We are grateful to Olivier Tombret (UMR 7209) for his help in the laboratory during sample preparation and to Denis Fiorillo (UMR 7209, SSMIM) for the stable isotope analysis. We also thank the Municipalidad Distrital de Huanchaco, the Department of Anthropology at Yale University, the National Geographic Society, INNOVATE PERU, Fondecyt-Concytec Perú–Banco Mundial, and Universidad Nacional de Trujillo for providing funds for the excavations at HLL. We are grateful to Belkys Gutiérrez Léon (BGL Arqueología, Peru) for her help with administrative issues, Jean-François Cuénot (UMR 8096, CNRS, France) for processing the map on ArcGIS, and Juan Plengue from the Reserva Chaparri for his assistance during the visit to Tinajones. We would also like to thank the anonymous reviewers and the editor for their constructive remarks.
Data Availability Statement
The processed starch grain samples are stored and available at the MSH Mondes. The original samples of the gut contents are available for some of the camelids only and are also stored at the MSH Mondes. The isotopic values are available in this article and in Dufour and colleagues (Reference Dufour, Goepfert, Neün, Prieto and Verano2020).
Supplemental Materials
For supplemental material accompanying this article, visit https://doi.org/10.1017/laq.2021.19.
Supplemental Table 1. List of the South American Plants in the MSH Mondes Reference Collection that Produce Starch Grains.
Supplemental Table 2. Isotopic Values of Camelids from Huanchaquito-Las Llamas.












 Open access
Open access