



Profound changes in indigenous Caribbean foodways have occurred in the last five centuries. The movement of various plant- and animal-based foods between the Old and New Worlds is one of the central themes of the “Columbian Exchange” (Crosby Reference Crosby1972). The quantity and diversity of peoples, pathogens, things, ideas, and foods that comprise the Columbian Exchange and the impacts that they have had on the course of history are well documented in broad strokes (Denevan Reference Denevan1992; Sauer Reference Sauer1966), but our current understanding of this general story is lacking in many critical details. Key research questions that remain unresolved are the dating of the introduction of various foods to different areas, the timing and routes by which they were dispersed, the speed by which new foods and knowledge of food production were incorporated into indigenous foodways, the various factors that contributed to the adoption of certain foods and not others, and how these changing and dynamic culinary traditions varied over space and time both within and between various Amerindian populations.
Patterns of continuity and change in foodways related to cross-cultural encounters in the Caribbean have been previously explored using a broad array of archaeological and historical data (Deagan Reference Deagan2004; Keegan and Hofman Reference Keegan and Hofman2017; Newsom and Wing Reference Newsom and Wing2004; Rouse Reference Rouse1992; Sauer Reference Sauer1966). Although stable isotope methods have been widely applied to human paleodiet studies in the Caribbean for both the precolonial (Chinique de Armas et al. Reference Chinique de Armas, Buhay, Suárez, Bestel, Smith, Mowat and Roksandic2015, Reference Chinique de Armas, Roksandic, Suárez, Smith, Buhay and Roksandic2016, Reference Chinique de Armas, Roksandic, Nikitović, Suárez, Smith, Kanik, Jordá and Buhay2017; Keegan and DeNiro Reference Keegan and DeNiro1988; Krigbaum et al. Reference Krigbaum, Fitzpatrick and Bankaitis2013; Laffoon and de Vos Reference Laffoon, de Vos, Hofman and Duijvenbode2011; Laffoon et al. Reference Laffoon, Rojas and Hofman2013, Reference Laffoon, Hoogland, Davies and Hofman2016, Reference Laffoon, Sonnemann, Shafie, Hofman, Brandes and Davies2017; Mickleburgh and Laffoon Reference Mickleburgh, Laffoon and Reid2018; Norr Reference Norr and Righter2002; Pestle Reference Pestle2010, Reference Pestle, Keegan, Hofman and Ramos2013; Stokes Reference Stokes1998) and late colonial periods (Schroeder et al. Reference Schroeder, O'Connell, Evans, Shuler and Hedges2009; Sparkes et al. Reference Sparkes, Varney, Courtaud, Romon and Watters2012; Varney Reference Varney2003), they have yet to be used to investigate the critical period spanning the historical divide or the early colonial period. This lacuna is unfortunate because isotopic analyses of human remains are particularly well suited for paleodietary studies, being ideal for tracking rapid changes in dietary practices both within and between individuals and for exploring variation in diets within and between different social groups at multiple scales using a bottom-up approach. This type of information concerning indigenous diets is almost entirely neglected in the documentary record of the early colonial period.
In this study we examine patterns in carbon and nitrogen isotopes in dentine and bone collagen, as well as in carbon and oxygen isotopes in enamel and bone apatite, to reconstruct childhood and adult diets at the site of El Chorro de Maíta, Cuba (Figure 1). The analyzed skeletons primarily date to the crucial phase spanning the contact and early colonial periods of the Caribbean that witnessed the arrival of the Spanish, the invasion and colonization of Cuba, and the imposition of the encomienda system of forced labor on the indigenous populations of the island. We use the patterning in the stable isotope results at multiple scales—in conjunction with archaeological, osteoarchaeological, zooarchaeological, and historical evidence—to explore the complex interrelationships between diet and mobility among this indigenous community and how existing patterns were affected after AD 1492. The main research aims of this study are to reconstruct patterns of indigenous foodways at the site of El Chorro de Maíta, to investigate changes in dietary practices in early colonial contexts of the Caribbean, to explore potential intrapopulation differences in responding to changing social and ecological conditions, and to assess possible linkages between dietary and mobility patterns at multiple scales.

Figure 1. Map of Cuba showing the location of the site El Chorro de Maíta.
Investigating Indigenous Diets
Site Background and Context
This study focuses on the burial population of the site of El Chorro de Maíta, on the northern coast of eastern Cuba. The cemetery portion of the site was primarily excavated in the 1980s under the direction of José Guarch Delmonte (Reference Guarch Delmonte1990, Reference Guarch Delmonte1996) and was originally interpreted as an essentially indigenous space of precolonial character with an ephemeral contact period component and without an important link with Europeans. Subsequent archaeological research over the last few decades conducted by Roberto Valcárcel Rojas and colleagues has provided important new insights into many aspects regarding the site. This research at the site identified nonfunerary contexts including domestic spaces and refuse deposits, indicating an indigenous occupation beginning around the thirteenth century AD. Pottery, European metallic artifacts, and pig remains appear in the final layers of occupation mixed with indigenous materials. In the fifteenth and sixteenth centuries AD, the settlement was organized around a central area in which a cemetery was located (Valcárcel Rojas Reference Valcárcel Rojas2016).
Several unique aspects of the site and burial population make it ideally suited for the aims of the current study, including the fact that the skeletal population is one of the largest (MNI = 133; Weston and Valcárcel Rojas Reference Valcárcel Rojas2016) and most well-researched indigenous populations in the insular Caribbean; it is also one of the only indigenous Caribbean populations whose dating can be definitively linked to the early colonial period (ca. AD 1500–1550). A large suite of radiocarbon dates including extensive direct dating of many of the skeletons indicates that most individuals have 2-sigma date ranges spanning the historical divide (Bayliss et al. Reference Bayliss, Rojas, Ramsey, Gulliksen and van der Plicht2012; Valcárcel Rojas Reference Valcárcel Rojas2012, Reference Valcárcel Rojas2016). It is therefore impossible to assign most individuals to either the precontact or postcontact period based on radiocarbon dates alone. A detailed analysis of the calibrated radiocarbon dates, including the use of Bayesian modeling, suggests that the dates are concentrated in the contact or early colonial period (Bayliss et al. Reference Bayliss, Rojas, Ramsey, Gulliksen and van der Plicht2012; Valcarcel Rojas Reference Valcárcel Rojas2016:194). Additional evidence that many of the burials date to this period includes the widespread presence of domestic pig (Sus scrofa) bones, European pottery, and metal artifacts recovered from graves (Valcárcel Rojas Reference Valcárcel Rojas2016), in addition to the extended supine (Christian-style) placement of the body for many of the burials (Valcárcel Rojas Reference Valcárcel Rojas2012; Valcárcel Rojas et al. Reference Valcárcel Rojas, Weston, Mickleburgh, Laffoon, van Duijvenbode, Hofman and Duijvenbode2011, Reference Valcárcel Rojas, Samson and Hoogland2013).
Recent osteoarchaeological investigations (Weston and Valcárcel Rojas Reference Weston, Rojas and Roksandic2016) revealed important new insights into health and demography at the site, including a mortality profile consistent with a catastrophic episode such as an epidemic. Furthermore, large concentrations of burials associated with indigenous (Meillacan) pottery found at this site do not occur at indigenous sites in Cuba, where burial areas generally comprise few individuals predominantly in cave sites, or elsewhere in precolonial Antilles. In addition, the lack of secure chronological indicators of precontact burials, the diverse ethnic and geographical origins observed in the mortuary population (Valcárcel Rojas et al. Reference Valcárcel Rojas, Weston, Mickleburgh, Laffoon, van Duijvenbode, Hofman and Duijvenbode2011), and taphonomic and demographic indicators that point to catastrophic mortality (Weston and Valcárcel Rojas Reference Valcárcel Rojas2016) suggest that the cemetery itself is predominantly a colonial phenomenon resulting from Indigenous–European interactions; however, the possibility that it could include a small nucleus of precolonial burials cannot be ruled out (Valcárcel Rojas Reference Valcárcel Rojas2016; Weston and Valcárcel Rojas Reference Weston, Rojas and Roksandic2016). The indigenous population of the site was rapidly incorporated into the Spanish system (encomienda) of forced labor in the first decades of the sixteenth century AD (Valcárcel Rojas Reference Valcárcel Rojas2012, Reference Valcárcel Rojas2016; Valcárcel Rojas et al. Reference Valcárcel Rojas, Samson and Hoogland2013).
Zooarchaeological research conducted to date has focused on specific components of the site, and in terms of MNI the terrestrial mollusks are the most abundant (Pérez Iglesias Reference Pérez Iglesias2007, Reference Pérez Iglesias2008, Reference Pérez Iglesias2010, Reference Pérez Iglesias2011). Nevertheless, the principal contributions to the biomass came from fishing, followed by hunting hutia, a small native rodent, and collecting mollusks (Pérez Iglesias Reference Pérez Iglesias2008). Thus, the quantitative faunal analyses indicate that marine fish and various small mammals such as hutia (Capromys pilorides, Mysateles melanurus) and cave rats (Boromys offella, Boromys torrei) constituted the core of the animal-based nutrition (Rodríguez Arce Reference Rodríguez Arce1987; Valcárcel Rojas Reference Valcárcel Rojas2016). Importantly the faunal analyses also revealed the presence of domestic pig remains at the site and provided clear evidence for processing (butchery marks) of these animals for consumption (Pérez Iglesias and Valcárcel Rojas Reference Pérez Iglesias and Rojas2014). Because of the biases of the faunal record, questions nevertheless remain concerning how rapidly introduced foods were incorporated into indigenous diets, their importance to caloric intake, and variation in the relative contributions of different foods to individual diets. Although caution is warranted in the use of trace element data from archaeological bone (Burton and Price Reference Burton, Douglas Price, Ambrose and Katzenberg2000), previous analysis of trace element concentrations from El Chorro de Maíta (Taylor Reference Taylor1990) revealed variable patterns consistent with the consumption of both marine and vegetable foods, thereby indicating varied omnivorous diets.
Stable Isotopes and Dietary Reconstruction
Carbon isotope values (δ13C) vary between different types of plants based on the different photosynthetic pathways used to fix atmospheric carbon (Smith and Epstein Reference Smith and Epstein1971). Most plant species use the C3 cycle (Calvin-Benson) and have lower δ13C values, whereas a minority use either the C4 (Hatch-Slack) or CAM (Crassulacean acid metabolism) cycles and possess significantly higher δ13C values (Bender Reference Bender1971; Smith and Epstein Reference Smith and Epstein1971). In the precolonial Caribbean, maize (Zea mays) was one of the only widespread and economically important C4 crops (Newsom and Wing Reference Newsom and Wing2004), although other regionally available C4 (amaranths, chenopods) and CAM (agave, pineapple) plants may have been occasionally consumed (Pestle Reference Pestle2010). Recent microbotanical (starch grain) research has further highlighted the early appearance (ca. 7800 BP) and widespread consumption of maize in the precolonial insular Caribbean (e.g., Mickleburgh and Pagán-Jiménez Reference Mickleburgh and Pagán-Jiménez2012; Pagán-Jiménez Reference Pagán-Jiménez, Keegan, Hofman and Ramos2013; Pagán-Jiménez et al. Reference Pagán-Jiménez, Rodríguez-Ramos, Reid, van den Bel and Hofman2015). Carbon isotopes also vary between terrestrial and marine ecosystems based on differences in their primary carbon sources (DeNiro and Epstein Reference DeNiro and Epstein1978; Schoeninger and DeNiro Reference Schoeninger and DeNiro1984), with the former possessing much lower δ13C values than the latter.
Nitrogen isotope values (δ15N) in plants are generally low compared to animals because δ15N is enriched stepwise between each trophic level in a food chain such that herbivores, omnivores, and carnivores possess increasingly higher δ15N (DeNiro and Epstein Reference DeNiro and Epstein1978, Reference DeNiro and Epstein1981). Marine ecosystems also generally have enriched δ15N values relative to terrestrial ones, given that marine plants possess higher δ15N and that there are usually more trophic levels in marine ecosystems (DeNiro and Epstein Reference DeNiro and Epstein1978; Schoeninger et al. Reference Schoeninger, DeNiro and Tauber1983). Although the combination of higher δ15N and higher δ13C values among marine ecosystems is such that coupled carbon and nitrogen isotopes can often be used to distinguish between marine and terrestrial food sources (Hedges and Reynard Reference Hedges and Reynard2007), the coral reef ecosystems of the Caribbean region include many fish species with lower than expected δ15N values (similar to terrestrial animals). This limits the usefulness of this isotopic proxy for distinguishing between terrestrial and marine protein consumption (Keegan and DeNiro Reference Keegan and DeNiro1988).
The main principle of dietary reconstruction via stable isotope analysis is that the isotopic composition of consumed foods is reflected in the tissues of consumers (DeNiro and Epstein Reference DeNiro and Epstein1976). The earliest applications of stable isotope methods for paleodietary studies focused on inferring long-term changes in diet based on carbon isotopes in human bone collagen (Vogel and van de Merwe Reference Vogel and Van Der Merwe1977). Over the last four decades, stable isotope approaches have become widely used and invaluable tools in archaeological research. Because of preferential routing (Fernandes et al. Reference Fernandes, Nadeau and Grootes2012; Krueger and Sullivan Reference Krueger, Sullivan, Turnlund and Johnson1984), carbon isotopes in collagen primarily reflect dietary protein, whereas carbon isotopes in bone (or enamel) apatite reflect the average of all three macronutrient components (carbohydrates, fats, and proteins) within the diet (Ambrose and Norr Reference Ambrose, Norr, Lambert and Grupe1993; Tieszen and Fagre Reference Tieszen, Fagre, Lambert and Grupe1993).
Previous Isotopic Research
All strontium isotope (87Sr/86Sr) data (n = 88) and a small subset of the enamel carbon (δ13C) and oxygen (δ18O) isotope data (n = 12) from El Chorro de Maíta discussed herein were previously reported (Laffoon Reference Laffoon2012; Laffoon et al. Reference Laffoon, Rojas and Hofman2013; Valcárcel Rojas et al. Reference Valcárcel Rojas, Weston, Mickleburgh, Laffoon, van Duijvenbode, Hofman and Duijvenbode2011). In combination with other lines of evidence, these isotope results revealed a high proportion of nonlocal migrants (25%) with diverse natal origins (Figure 2). Based on the combined evidence, most of these nonlocals probably originated from elsewhere in the Caribbean, possibly even from other locations on Cuba, although two individuals exhibited considerable evidence indicating long-distance mainland origins. For individual CM72B, an adult female, the evidence for nonlocal origins includes deviant burial treatment; tall stature relative to the local population; exotic cranial and dental modification styles; and nonlocal enamel strontium, oxygen, and carbon isotope values. Based on this evidence, it was proposed that this individual probably originated from Mesoamerica (Valcárcel Rojas et al. Reference Valcárcel Rojas, Weston, Mickleburgh, Laffoon, van Duijvenbode, Hofman and Duijvenbode2011). A recent dual isotope provenance approach applied to this individual's combined strontium and oxygen isotope signatures confirms that lowland Mesoamerica is one of the few places in the tropical Americas from where this individual could have originated (Laffoon et al. Reference Laffoon, Sonnemann, Shafie, Hofman, Brandes and Davies2017). For individual CM45, an adult male, the evidence for nonlocal origins includes a burial position that is atypical for indigenous Antilleans; grave inclusions of brass aglets (clothes fasteners) of European origin (Cooper et al. Reference Cooper, Martinón-Torres, Rojas, Hofman, Hoogland and van Gijn2008; Martinón-Torres et al. Reference Martinón-Torres, Rojas, Cooper and Rehren2007; Valcárcel Rojas Reference Valcárcel Rojas2016); cranial metrics indicative of likely African ancestry; and highly elevated enamel isotope values of strontium, oxygen, and carbon that excluded the possibility of Antillean origins. Based on the overall evidence this individual was interpreted as a first-generation migrant from Africa (Valcárcel Rojas et al. Reference Valcárcel Rojas, Weston, Mickleburgh, Laffoon, van Duijvenbode, Hofman and Duijvenbode2011).
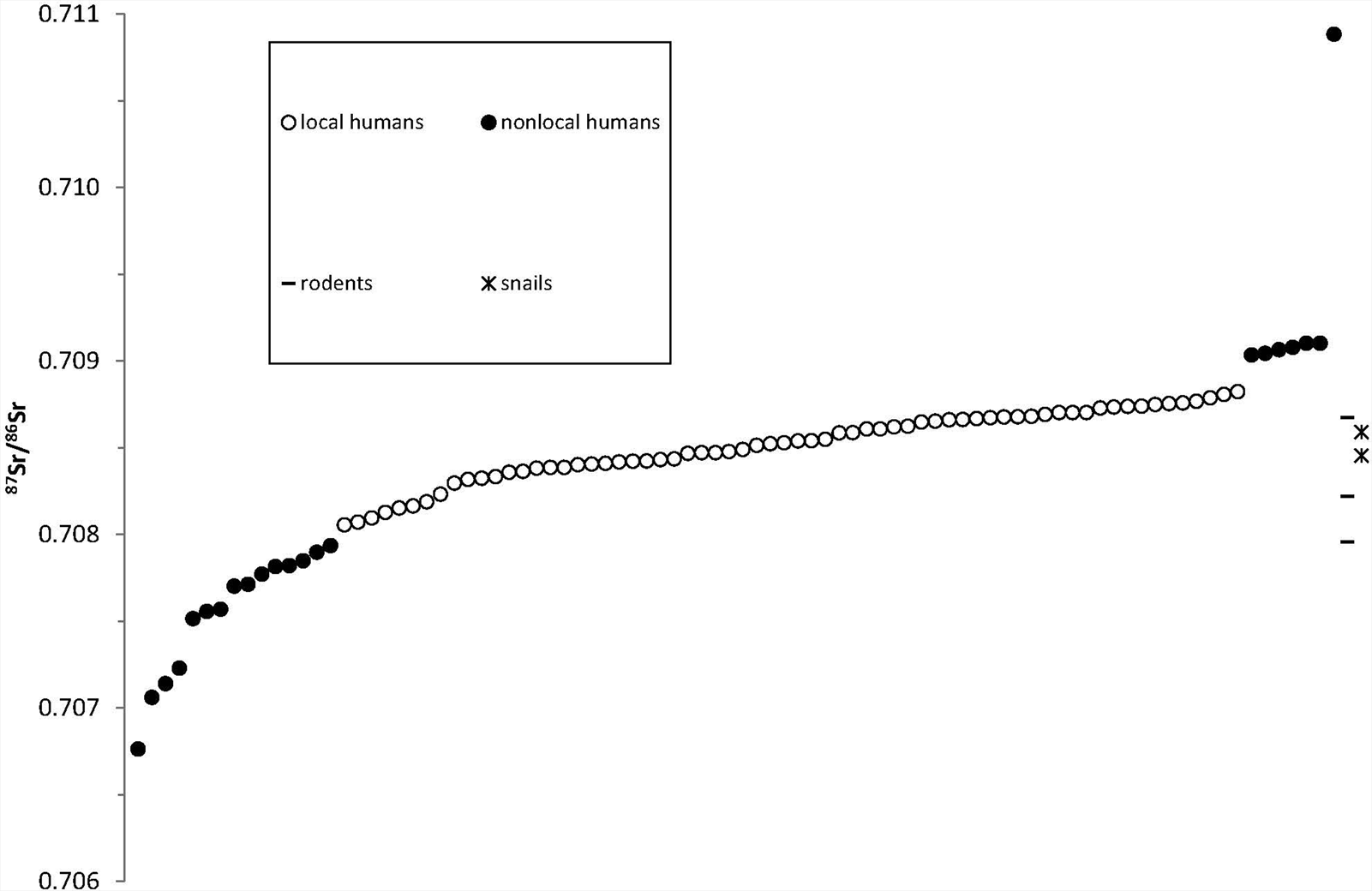
Figure 2. Plot of 87Sr/86Sr ratios from El Chorro de Maíta showing locals (white symbols) and nonlocals (black symbols). (Data sources: Laffoon Reference Laffoon2012; Valcárcel Rojas et al. Reference Valcárcel Rojas, Weston, Mickleburgh, Laffoon, van Duijvenbode, Hofman and Duijvenbode2011.)
In addition to the extensive suite of radiocarbon dates, a preliminary dataset of associated stable isotope results has also been reported for this burial population (Bayliss et al. Reference Bayliss, Rojas, Ramsey, Gulliksen and van der Plicht2012). This preliminary bone collagen carbon and nitrogen isotope data have confirmed both a high degree of dietary variation and tentative links between nonlocal origins and nonlocal dietary practices.
Materials and Methods
Materials
The sampling strategy used for this study focused on both bone and dentine collagen and bone and enamel bioapatite (structural carbonate). This strategy permitted explorations of age-based variation in dietary practices, including changes occurring within the life of a single individual from a life-history perspective (e.g., Laffoon et al. Reference Laffoon, Espersen and Mickleburgh2018; Sealy et al. Reference Sealy, Armstrong and Schrire1995). Sampling of dental elements targeted premolars for consistency and to avoid the complicating effects of breastfeeding inputs; however, in several cases other teeth were also sampled when intact premolars were not available. Enamel and dentine samples were taken from the same dental elements. Bone samples were primarily derived from long bone fragments. All samples were processed at the Stable Isotope Laboratory, Faculty of Science, at Vrije Universiteit Amsterdam.
Methods
Bone and teeth samples were cleaned by mechanically removing the outer surface, which is the most susceptible to diagenetic alteration (Budd et al. Reference Budd, Montgomery, Barreiro and Thomas2000). The extraction of collagen from bone and dentine followed standard protocols for archaeological skeletal materials (Ambrose Reference Ambrose1990; Brown et al. Reference Brown, Erle Nelson, Vogel and Southon1988). Samples were demineralized in 0.6 M HCl at 4°C for several days, rinsed to neutral with ultrapure water (Milli-Q), treated with 0.125 M NaOH for 20 hours, rinsed to neutral again, gelatinized in pH = 3 HCl at 80°C for 48–72 hours, separated with Ezee (Elkay) filters, frozen, and lyophilized. The processing of bone and enamel apatite samples for isotope analyses included a chemical pretreatment procedure (Bocherens et al. Reference Bocherens, Sandrock, Kullmer and Schrenk2011), which involved washing in 2.5% bleach (NaOCl) rinsing, and then leaching in Ca-acetate buffered acetic acid (CH3COOH, pH = 4.75). Carbon and nitrogen isotope measurements were conducted on a ThermoQuest IRMS Delta XP-plus interfaced to a Flash elemental analyzer. International standards (USGS40, USGS41, IAEA-310(A) and IAEA-NO3) were used for sample calibration, with long-term reproducibility of standards of ± 0.1‰ (1SD) for both δ13C and δ15N. Carbon and oxygen isotope measurements were conducted on a Finnigan DeltaPlus IRMS coupled with a Gasbench II; long-term reproducibility of the international reference material (NBS19) is ± 0.1‰ (1SD) for δ13C and ± 0.2‰ (1SD) for δ18O. Stable isotope results are reported in the δ notation, in parts per thousand (‰) relative to the international VPDB (carbon and oxygen) and AIR (nitrogen) standards.
Results
Quality Control Indicators
The sampling information and isotope results are presented in Supplemental Table 1. Not all isotope proxies are available for every individual because some did not possess suitable intact dental elements, whereas others had poor bone preservation (and are not considered further). For all samples reported here, multiple quality control indicators suggest overall good preservation of collagen material with C:N ratios within the range (2.9–3.6) reported for nonaltered bone and with wt.% carbon and wt.% nitrogen higher than 13.0% and 4.8%, respectively (Ambrose Reference Ambrose1990; DeNiro Reference DeNiro1985). Collagen yields were variable for bone collagen, ranging from 2.5% to 26.2% (mean: 7.6%), and for dentine collagen, ranging from 1.8% to 16.5% (mean: 8.5%) but consistent with those reported for precolonial Caribbean populations (Pestle and Colvard Reference Pestle and Colvard2012). Collagen stable isotope values tended to remain consistent for collagen yields higher than 1% (Ambrose Reference Ambrose1990). Unlike for bone collagen, there is no general consensus concerning quality-control indicators for bone apatite (Shin and Hedges Reference Shin and Hedges2012), but given its known susceptibility to multiple diagenetic processes (Koch et al. Reference Koch, Tuross and Fogel1997), we used general and pairwise comparisons of bone and enamel carbon (δ13Capat and δ13Cenam) and oxygen (δ18Oapat and δ18Oenam) to test for potential postmortem alteration of biogenic isotope values. We also considered the bone apatite isotope data as supplementary to the other isotope proxies and focused our interpretations on the more robust collagen and enamel isotope datasets.
Enamel and Bone Apatite Results: Carbon and Oxygen
Summary statistics of isotope results for all samples and for specific subgroups based on sex, age, and origins are presented in Supplemental Table 2. Enamel and bone apatite carbon and oxygen isotope values are plotted in Figure 3. The mean δ13Cenam for all individuals is −11.4 ± 0.9‰ (−15.5‰ to −9.7‰, n = 69), and the mean δ18Oenam is −3.7 ± 0.7‰ (−5.4‰ to −2.0‰). These statistics exclude the two nonlocal individuals (CM45, CM72B) with long-distance (mainland) origins, who possess δ13Cenam (−5.1‰ and −3.7‰) and δ18Oenam values (−5.4‰ and −3.7‰), respectively: these values are highly elevated both relative to the local population and compared to all other Caribbean archaeological populations (Laffoon et al. Reference Laffoon, Rojas and Hofman2013).

Figure 3. Plot of enamel and bone apatite δ13C and δ18O values from El Chorro de Maíta. (Data sources: Laffoon et al. Reference Laffoon, Rojas and Hofman2013 and this study.)
The mean δ13Capat for all samples is −10.5 ± 0.6‰ (−12.3‰ to −8.7‰, n = 41), and the mean δ18Oenam is −6.3 ± 0.5‰ (−7.2‰ to −5.4‰). The mean δ13Cenam and mean δ13Capat are broadly similar, and the corresponding ranges display a high degree of overlap. In contrast, the mean δ18Oenam and mean δ18Oapat differ by 2.6‰ with the latter consistently lower than the former and almost no overlap in the range of values. There is a statistically significant difference between both δ13Cenam and δ13Capat t(102) = −3.81, p < 0.001, and between δ18Oenam and δ18Oapat t(103) = 23.29, p < 0.001.
We identified intra-individual variation in apatite isotope values by comparing enamel and bone apatite isotope values from the same individuals (Figure 4). For the vast majority of individuals, δ13Cenam is more negative than δ13Capat, but the offset (Δ13Cenam-apat) between them is highly variable, with a mean of −1.1 ± 1.2‰ (−5.0‰ to + 1.8‰, range: 6.8‰). Intra-individual offsets in oxygen isotope values (Δ18Oenam-apat) are larger (mean: 2.4 ± 0.8‰) than those observed for Δ13Cenam-apat but with reduced variation (1.0‰ to 4.7‰). The variability in Δ13Cenam-apat, including both negative and positive values, was previously documented for both animals and humans, but its possible causes are not well understood (Laffoon et al. Reference Laffoon, Hoogland, Davies and Hofman2016; Loftus and Sealy Reference Loftus and Sealy2012).

Figure 4. Paired enamel (δ13Cenam and δ18Oenam) and bone apatite (δ13Capat and δ18Oapat) isotope values of the same individuals from El Chorro de Maíta.
Dentine and Bone Collagen Results: Carbon and Nitrogen
Dentine and bone collagen isotope values are plotted in Figure 5. The mean δ13Cdent for all individuals is −17.7 ± 1.3‰ (−19.0‰ to −12.2‰, n = 35), and the mean δ15Ndent is 13.8 ± 1.3‰ (10.2‰ to 15.7‰). The mean δ13Ccoll is −17.6 ± 2.1‰ (−22.8‰ to −9.8‰, n = 44), and the mean δ15Ncoll is 13.2 ± 1.4‰ (8.5‰ to 14.9‰). The two individuals with documented long-distance origins (CM45 and CM72B) were excluded from these summary statistics because their collagen isotope values are extreme outliers and are highly elevated for both dentine, δ13Cdent (−9.1‰ and −8.6‰) and δ15Ndent (9.6‰ and 9.3‰), and bone collagen δ13Ccoll (−7.2‰ and −8.0‰) and δ15Ncoll (10.6‰ and 8.4‰), respectively (Bayliss et al. Reference Bayliss, Rojas, Ramsey, Gulliksen and van der Plicht2012). Overall, the dentine and bone collagen datasets possess similar mean δ13C and δ15N values, differing by less than 1‰, and there is a high degree of overlap in the ranges of values, although the bone collagen dataset possesses a greater range of values for both δ13C (>15‰) and δ15N (>6‰). The collagen isotope dataset exhibits notably extreme variability, which is far greater than that reported for other indigenous populations in the Caribbean.

Figure 5. Plot of dentine collagen (δ13Cdent and δ15Ndent) and bone collagen (δ13Ccoll and δ15Ncoll) isotope values from El Chorro de Maíta.
Intra-individual differences in collagen isotope values based on paired dentine and bone samples (Figure 6) are on average quite small, with mean offsets of 0.3‰ for Δ13Cdent-coll and 0.7‰ for Δ15Ndent-coll. Nevertheless, there is considerable variation among these offsets, with Δ13Cdent-coll varying from −1.8‰ to + 4.8‰ (range: 6.6‰) and Δ15Ndent-coll varying from −2.8‰ to + 7.1‰ (range: 9.9‰). It is noteworthy that the direction of the offset is also quite variable, with roughly equal proportions possessing positive and negative offsets.

Figure 6. Paired dentine (δ13Cdent and δ15Ndent) and bone collagen (δ13Ccoll and δ15Ncoll) isotope values of the same individuals from El Chorro de Maíta.
Discussion
Dietary Reconstructions
Owing to the high variance and extremely large ranges of the stable isotope data overall, and of the collagen isotope data in particular, it is difficult to make broad generalizations about the entire sample population. Nevertheless several trends are apparent. For example, we derived estimates of whole-diet carbon isotope values (δ13Cwhole diet) by subtracting 10.1‰ from the enamel and bone apatite δ13C values (Fernandes et al. Reference Fernandes, Nadeau and Grootes2012), reflecting the average (whole dietary) intake of the childhood and adult periods, respectively. The δ13Cwhole diet values of the majority of the dataset fall within the range of C3 plants, indicating a predominance of C3 resources in the plant component of the diet, but several samples have δ13Cwhole diet values falling at the edge or just outside of the range of values reported for C3 plants in the Caribbean. This likely indicates minor contributions of C4, marine resources, or both (Mickleburgh and Laffoon Reference Mickleburgh, Laffoon and Reid2018). These patterns are consistent with the ecology of Cuba and the broader Caribbean region, which is dominated by C3 plants and where only a limited number of C4 (such as maize) or CAM plants (agave, pineapple) have been documented as human food sources (Newson and Wing Reference Newsom and Wing2004; Pestle Reference Pestle2010). Individuals CM45 (African origin) and CM72B (Mesoamerican origin) have highly elevated δ13Cwhole diet estimates calculated from their δ13Cenam values, indicating a predominance of C4 resources during childhood, likely from their respective consumption of sorghum/millet and maize (Laffoon et al. Reference Laffoon, Rojas and Hofman2013).
Turning to assessments of intra-individual variation in isotope values, a few clear patterns emerge. First, there are at least two possible explanations for the systematic differences between the enamel and bone oxygen isotope values: (1) the δ18Oenam are elevated relative to bone δ18Oapat owing to the influence of breastfeeding, because breastmilk is expected to be elevated in δ18O (Britton et al. Reference Britton, Fuller, Tütken, Mays and Richards2015), and (2) the δ18Oapat values are systematically offset by postmortem alteration or issues of sample treatment and data calibration (Pestle et al. Reference Pestle, Crowley and Weirauch2014). The enamel and bone apatite δ18O values have been used to calculate drinking water oxygen isotope (δ18Odw) values by first converting to the VSMOW scale using the formula of Coplen (Reference Coplen1988) and then applying the calculation of Chenery and colleagues (Reference Chenery, Pashley, Lamb, Sloane and Evans2012). Comparison of these δ18Odw values with the range of precipitation δ18O for the circum-Caribbean (International Atomic Energy Agency/World Meteorological Organization 2016; Laffoon et al. Reference Laffoon, Sonnemann, Shafie, Hofman, Brandes and Davies2017; Terzer et al. Reference Terzer, Wassenaar, Araguás-Araguás and Aggarwal2013) indicates a high degree of overlap with δ18Odw values derived from the El Chorro de Maíta δ18Oenam dataset but not the δ18Oapat dataset. As such, and because enamel is generally considered a more reliable sample substrate for oxygen isotopic analyses (Sponheimer and Lee-Thorp Reference Sponheimer and Lee-Thorp1999), we conclude that the bone oxygen isotope data are unreliable and do not consider them further.
Estimates of the protein component of the diet (Figure 7) are derived by applying the conversion formula from Pestle and colleagues (Reference Pestle, Hubbe, Smith and Stevenson2015) to obtain δ13Cprotein, and by subtracting 3.6‰ from dentine and bone δ15N values to obtain δ15Nprotein, as suggested by various studies (Ambrose Reference Ambrose, Ambrose and Anne Katzenberg2002; DeNiro and Epstein Reference DeNiro and Epstein1981). When compared to the mean (± SD) of plant (C3 and C4) and animal (land and marine) food groups available in the precolonial Caribbean (Pestle Reference Pestle2010), the estimated δ13Cprotein values of the local population overlap considerably with the range of δ13C for both C3 plants and land animals but not with C4 plants or marine animals. In contrast, the δ15Nprotein values of the majority of samples overlap primarily with the range of δ15N in (pelagic) marine animals. Therefore, the estimated protein diet at the site does not match well with any of these four specific native Caribbean food groups. It should be noted that (combined) low carbon and high nitrogen consumer collagen isotope values have been interpreted as reflecting the consumption of freshwater fish in certain archaeological contexts (e.g., Harrison and Katzenberg Reference Harrison and Katzenberg2003), but no archaeological, faunal, or textual evidence indicates that this is a feasible primary food source at El Chorro de Maíta.

Figure 7. Estimated protein carbon isotope values (δ13Cprotein) derived by applying the conversion formula from Pestle and colleagues (Reference Pestle, Hubbe, Smith and Stevenson2015) to combined collagen and apatite carbon isotope values, plotted against estimated protein nitrogen isotope values (δ15Nprotein) derived by subtracting 3.6‰ from collagen nitrogen isotope values. (Faunal isotope data sources: Pestle Reference Pestle2010; Stokes Reference Stokes1998.)
Abundant historical records document both the early arrival and rapid expansion of domestic pig populations within the Spanish colonies of the Caribbean (del Río Moreno Reference del Río Moreno1996). Domestic pigs are expected to have low δ13C owing to a reliance on C3 plants, in addition to moderately enriched δ15N, due to omnivory. A small dataset (n = 9) of pig dentine collagen stable isotope values (mean δ13Cdent = −20.7 ± 1.4‰; mean δ15Ndent = 7.6 ± 1.1‰) from various sites in the Caribbean displays this pattern, with isotope values that are comparable to the main cluster of local El Chorro de Maíta individuals. There is nevertheless considerable variation in the estimated protein isotope values, which indicates substantial diversity in the relative contributions of these different food groups to individual diets. In general terms, these comparative data are consistent with the combined human stable isotope results, indicating mixed diets incorporating plants and marine and terrestrial animals but with a major contribution from the meat of domestic livestock (pigs) to the protein component of the diet for the majority of the local population.
Intrapopulation Variation
The observed diversity in dietary practices evidenced by the stable isotope data is at least partially explained by the large numbers of nonlocals within the population and their highly diverse origins. This diversity is most evident in the isotope values of individuals CM45 and CM72B who appear to have been highly reliant on C4 plants but is also found in several individuals with very low δ13Cprotein and δ15Nprotein values approaching a pure C3 plant diet. Notably, three out of the four most depleted δ13Cprotein values are from juveniles, whereas three out of the four most enriched δ13Cprotein values are from nonlocals. Given that the nonlocals CM45 and CM72B also have deviant bone collagen isotope values, they probably died before their bones could equilibrate to the local dietary pattern. This is consistent with the high rates of mortality among enslaved individuals in colonial times and the potentially catastrophic mortuary profile of the El Chorro de Maíta cemetery population (Weston and Valcárcel Rojas Reference Weston, Rojas and Roksandic2016). Nevertheless, statistical tests based on sex, age, geographic origins, or cranial modifications only identified significant differences in δ15Ndent for two variables. Specifically, there are statistically significant differences in δ15Ndent based on age, with juveniles having significantly higher values (M = 14.6‰, SD = 0.7) than adults (M = 13.1‰, SD = 2.9); t(34) = 3.65, p < 0.001; and based on origins, with locals having significantly higher δ15Ndent (M = 14.2‰, SD = 1.3) than nonlocals (M = 11.8‰, SD = 2.6); t(12) = 4.23, p = 0.001.
The differences in δ15Ndent between juveniles and adults are notable because the values in both groups should in principle reflect roughly the same ages of formation. Given that the sampling of teeth for dentine collagen targeted the later-forming distal tips of the roots to avoid possible influences of breastmilk consumption on measured isotope values, it is not likely that a breastfeeding effect could account for the enriched dentine δ15N values of either group. By contrast, the observed differences in δ15Ndent between adults and juveniles may reflect differential survivorship, with the former representing survivors and the latter nonsurvivors (e.g., Beaumont et al. Reference Beaumont, Gledhill, Lee-Thorp and Montgomery2013). If so, this specific data pattern might indicate a possible link between childhood malnutrition (indicated by elevated δ15Ndent) and mortality (juvenile age at death; Gowland Reference Gowland2015), but more research is required to explore this in greater depth.
Large differences in δ15Ndent relative to different (natal) geographic origins could result if local individuals, particularly local juveniles, born and raised during the imposition of the encomienda system, would have had access to higher trophic level food sources during childhood. Nonlocal individuals, by contrast, could have had quite different dietary habits in their youth, which would be preserved in their dentine stable isotope signals and reflect the food cultures of their various homelands. Yet it should be noted that it is somewhat questionable to treat nonlocals of diverse origins as a coherent statistical group for the sake of comparisons, given the diversity of origins suggested by the isotopic, archaeological, and bioarchaeological evidence (Laffoon Reference Laffoon and Roksandic2016).
Several patterns are evident concerning the observed differences between enamel and bone apatite δ13C values. For the vast majority of individuals, δ13Cenam is lower than δ13Capat. The generally lower δ13Cenam values could result from a breastfeeding effect or simply reflect minor age-related differences in dietary intake over time. Interestingly, the slight offset between δ13Cenam and δ13Capat is also apparent for juvenile individuals, potentially owing to differences in the time spans represented by the two sample types. Importantly, the differences between these two datasets are not statistically significant either at the scale of the entire sample or for paired tooth-bone samples of the same individuals, and the degree of intra-individual offsets is highly variable. Such patterning could also perhaps be explained by differential diagenesis of bone samples if, for example, some bone apatite samples were more isotopically altered than others. Nonetheless, the lack of systematic differences between the enamel and bone apatite carbon isotope values and the fact that all but three of the latter fall within the range of the former would seem to suggest a lack of diagenetic alteration in the δ13Capat dataset, thus indicating a high degree of correspondence between childhood and adult diets.
In fact, although large intra-individual differences were observed for paired enamel-bone and paired dentine-bone samples for several individuals, this large variability was not limited to nonlocals. This pattern could result from a limitation of the strontium isotope method itself, which cannot identify nonlocals originating from isotopically similar regions (false negatives); alternatively it could indicate that large-scale dietary changes also occurred among some locals. This second possibility would not be too surprising, considering (1) that many other regions of Cuba (and the Caribbean in general) possess similar ranges of bioavailable 87Sr/86Sr (Laffoon et al. Reference Laffoon, Davies, Hoogland and Hofman2012) and given (2) the documented diversity of dietary practices on such a large, and ecologically and culturally diverse, island as Cuba (Chinique de Armas et al. Reference Chinique de Armas, Roksandic, Suárez, Smith, Buhay and Roksandic2016). Lastly, the relatively limited range (ca. 4‰) of enamel/apatite δ13C values contrasts sharply with the enormous range (ca. 13‰, excluding nonlocals) of dentine/bone collagen δ13C, possibly indicating a greater diversity of protein diets compared to whole diets.
Interpopulation Variation
Placing the isotopic datasets within a broader regional context, we compared the collagen isotope data with several precolonial and colonial period populations from the Caribbean (Figure 8). Starting with intra-island comparisons, the local El Chorro de Maíta population has higher values for both δ13C and δ15N than most other indigenous Cuban populations, with the notable exception of Canímar Abajo. Although the locals at El Chorro de Maíta and the Canímar Abajo population have overlapping δ13C ranges, the mean δ15N is more than 2‰ higher for the former. At Canímar Abajo, diets are purported to have been heavily reliant on plant and shellfish resources (Chinique de Armas et al. Reference Chinique de Armas, Roksandic, Suárez, Smith, Buhay and Roksandic2016), which may account for their relatively lower δ15N values. Turning to comparisons with precolonial Caribbean populations, the El Chorro de Maíta dataset also possesses a broadly similar range of collagen δ13C values but elevated δ15N values compared to other indigenous populations from across the Antilles. In fact, the combined collagen δ13C and δ15N values for El Chorro de Maíta display little overlap with any of the indigenous populations from Cuba or the insular Caribbean more broadly. The higher δ15N values at El Chorro de Maíta indicate a larger relative contribution of animal protein, the consumption of higher trophic level animal resources compared to other precolonial Caribbean populations, or both.
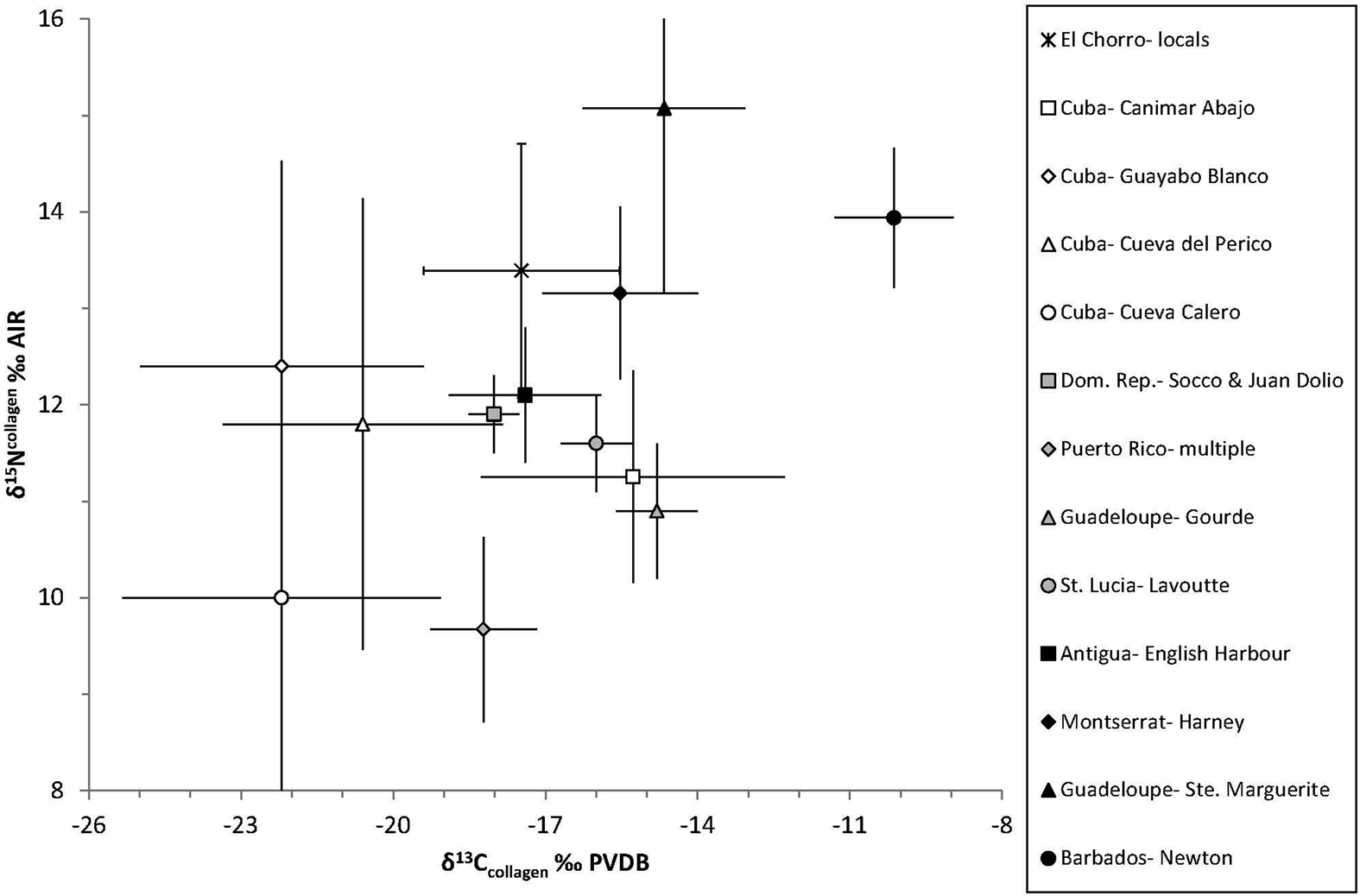
Figure 8. Bone collagen stable isotope values (δ13C and δ15N) from El Chorro de Maíta compared to precolonial (indigenous) populations from Cuba and to precolonial (indigenous) populations from elsewhere in the Antilles, and late colonial period (enslaved) populations from the Antilles (black symbols). (Data sources: Chinique de Armas et al. Reference Chinique de Armas, Roksandic, Suárez, Smith, Buhay and Roksandic2016; Laffoon and de Vos Reference Laffoon, de Vos, Hofman and Duijvenbode2011; Laffoon et al. Reference Laffoon and Roksandic2016; Pestle Reference Pestle2010; Schroeder et al. Reference Schroeder, O'Connell, Evans, Shuler and Hedges2009; Sparkes et al. Reference Sparkes, Varney, Courtaud, Romon and Watters2012; Stokes Reference Stokes1998; Varney Reference Varney2003.)
This conclusion is supported by comparing the El Chorro de Maíta results with several colonial-era populations from the Caribbean (Figure 8). In terms of combined collagen carbon and nitrogen isotope values, the local El Chorro de Maíta population is comparable to some, but not all, colonial period enslaved African populations within the Antilles; for example, at English Harbour, Antigua, and Harney, Monserrat (Sparkes et al. Reference Sparkes, Varney, Courtaud, Romon and Watters2012; Varney Reference Varney2003). In contrast, other enslaved African populations from the colonial-era Caribbean such as Ste. Marguerite, Guadeloupe (Sparkes et al. Reference Sparkes, Varney, Courtaud, Romon and Watters2012), and Newton Plantation, Barbados (Schroeder et al. Reference Schroeder, O'Connell, Evans, Shuler and Hedges2009), possess considerably higher collagen δ13C and δ15N values than El Chorro de Maíta. The combined collagen isotope values at these latter two sites are more consistent with higher rates of marine protein consumption. Interestingly, the enamel and bone apatite δ13C values at El Chorro de Maíta are broadly similar in both precolonial and colonial period Caribbean populations and display much less spatial or temporal variation than collagen δ13C values (Stokes Reference Stokes1998). One plausible explanation for these observations is that there was a pronounced change in protein diets across the historical divide but comparatively less change to the plant component of diets (at least in terms of the relative proportions of C3 and C4 plants).
In summary, the overall patterns in stable isotope results at El Chorro de Maíta and other colonial period sites in the Caribbean reflect greater access to terrestrial animal protein sources during the colonial period, likely as the result of the introduction and dispersal of European domestic livestock within extensive colonial economic systems. The incorporation of Old World domestic livestock such as pigs into dietary regimes that were predominantly indigenous oriented is consistent with the observed patterning of the stable isotope and zooarchaeological data at El Chorro de Maíta. This is evidenced by the general lack of difference in enamel/apatite δ13C values between El Chorro de Maíta and other indigenous Caribbean populations, the intermediate collagen δ13C values indicating mixed diets of plant and animal proteins, and especially the elevated δ15N values at El Chorro de Maíta that correspond well with those observed for domestic pigs in the Antilles. This pattern may also reflect a broader regional trend by which specific Old World animals were fairly rapidly incorporated into New World subsistence economies during the early years of Spanish colonization of the Caribbean (del Río Moreno Reference del Río Moreno1996).
Conclusions
In this article we presented a large corpus of stable isotope data from the burial population of El Chorro de Maita, Cuba, and provided new insights into dynamic patterns of dietary diversity and change during the first generations of the colonization of the New World. The combined isotope results, particularly collagen isotope results, indicate extremely diverse diets at El Chorro de Maíta among not only nonlocal individuals but also within the local population. There are many possible reasons why the local indigenous population may have had highly variable diets in the early colonial period, including but not limited to differences in social status, socioeconomic relations, work activities or occupations, and access to food sources, as well as the possibility that the cemetery includes individuals who did not reside at the site but originated from other places in Cuba, or beyond, with distinct dietary habits and culinary traditions.
A few long-distance immigrants, including one originating from Mesoamerica (CM72B) and one from Africa (CM45), had correspondingly distinct dietary habits. Therefore, the multiple isotope proxies indicate that some nonlocals either maintained the dietary patterns of their homelands after migration or more likely died before their tissues could equilibrate to local dietary patterns. There are nevertheless very few systematic differences between different social groups based on demographic factors (sex and age) or geographic origins (locality). The significant differences in dentine nitrogen isotope values between juveniles and adults and between locals and nonlocals reflect not only the correlation of these variables but also the possibility that locally born juveniles who died in the sixteenth century had distinctly different diets than did other groups within this burial population. This conclusion is supported by the fact that a number of individuals possess pronounced (intra-individual) differences in isotope values from different sample types (paired enamel-bone or dentine-bone samples), including several locally born individuals. These patterns may record individuals who experienced large-scale dietary changes over life courses spanning the historical divide between precolonial and colonial times.
Large-scale chronological changes in dietary patterns are also indicated by the greater similarity of the El Chorro de Maíta isotope results to certain enslaved African populations, rather than to other precolonial populations from Cuba or the Caribbean in general. Thus, differences in dietary practices at multiple scales from the individual to the population level suggest relatively rapid (within ca. 50 years) transitions in indigenous foodways. Although the highly variable human isotope data and general lack of comparative baseline (foodweb) isotopic data for the site make it difficult to generalize about dietary reconstructions, the majority of locals at El Chorro de Maíta seem to have consumed mixed diets dominated by a combination of C3 plants and terrestrial animal proteins. Nevertheless, the moderately elevated apatite and collagen isotope values are not consistent with pure C3 diets and suggest smaller but substantial contributions of C4 plants, seafood, or both. Pork is a likely source of the consumed terrestrial animal protein based on the combined historical, zooarchaeological, and isotopic evidence, but the consumption of meat from other imported domestic livestock cannot be ruled out. The totality of evidence thus suggests that under the encomienda system the indigenous community at this site obtained rapid access to newly introduced foods either via provisioning by Europeans, the hunting of feral pigs, or even the raising of domestic pigs themselves.
Overall, the high diversity in diets documented at El Chorro de Maíta cannot clearly be attributed to any single cause and probably results from the interplay of a multitude of factors including temporal, social, cultural, demographic, and idiosyncratic variables. Several unresolved questions concerning various aspects of dietary and subsistence patterning at El Chorro de Maíta will be addressed in future research. These include expanding the baseline foodweb stable isotope data for both the precolonial and colonial period sites on Cuba, quantifying dietary inputs using Bayesian multiple source mixing models, investigating temporal changes in weaning practices via serial sampling and stable isotope analyses of dentine collagen, and conducting compound-specific isotope analyses (CSIA) of individual amino acids to disentangle the relative influences of terrestrial and marine food sources. Lastly, it is our hope that the research framework developed for this study can be applied to investigate the continuity and change of indigenous lifeways in the face of culture contact, conquest, and colonization at other sites within the Caribbean and the Americas more broadly.
Acknowledgments
The research leading to these results is part of the ERC-Synergy project NEXUS1492, under the direction of Corinne L. Hofman, which has received funding from the European Research Council under the European Union's Seventh Framework Programme (FP7/2007-2013)/ERC grant agreement no. 319209. We are extremely grateful to the Departamento Centro-Oriental de Arqueología, Ministerio de Ciencia, Tecnología y Medio Ambiente (Cuba) and Comisión Nacional de Monumentos de Cuba for providing access and permission (PEA-1/15) to analyze the El Chorro de Maíta skeletal collection, and to the Museo El Chorro de Maíta for facilitating this research. We also wish to acknowledge the contributions of Suzan Verdegaal-Warmerdam and Robin van der Velde at the Free University Amsterdam, who conducted the stable isotope measurements and assisted with the sample preparation and processing, respectively.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.
Supplemental Material
For supplementary material accompanying this article, visit https://doi.org/10.1017/laq.2019.103.
Supplemental Table 1. Sample Information and Isotope Results from El Chorro de Maíta, Cuba.
Supplemental Table 2. Statistical Summary of Stable Isotope Data from El Chorro de Maíta, Cuba.











 Open access
Open access