



Introduction
Length–weight relationships (LWRs) of fish are important for fisheries science as they allow the estimation of biomass from commercial length-frequency data (Anderson and Gutreuter, Reference Anderson, Gutreuter, Nielsen and Johnson1983), are relevant input parameters for length-based assessments of data-limited stocks (Froese, Reference Froese2004; Chong et al., Reference Chong, Mildenberger, Rudd, Taylor, Cope, Branch, Wolff and Stäbler2019) and facilitate studies of spatio-temporal variation in fish condition, including inter-regional comparisons of life histories (Petrakis and Stergiou, Reference Petrakis and Stergiou1995). Therefore, the use of accurate LWR parameters is critical to obtain reliable stock assessment results leading to scientifically robust management recommendations (Gerritsen and McGrath, Reference Gerritsen and McGrath2007).
Atlantic Moroccan waters (Exclusive Economic Zones from 27°40′00″ N to 36°00′18.4″ N, 02°03′27.1″ W to 13°39′53.1″ W, including the Mediterranean sector) are strongly influenced by one of the major upwelling systems located in the south, with fisheries including both national artisanal and international industrial fleets mainly targeting demersal species (FAO, 2020; RCG LDF, 2022). Regular monitoring activities are carried out by the ‘Institut National de Recherche Halieutique’ (INRH); however, there are data gaps relating to the life history of some target and bycatch species caught by fisheries operating in the Fishery Committee for the Eastern Central Atlantic (CECAF) area (FAO, 2020).
In the 2000's, the Spanish Oceanographic Institute (IEO, CSIC) launched a partnership programme with the INRH to study the resources and ecosystems in deep waters. As part of this programme, several scientific surveys were planned to study the biodiversity of demersal species along the continental shelf off Morocco and Western Sahara, in order to increase the knowledge on the species caught and their biology, including length–weight data.
Materials and methods
The present study analysed data obtained onboard the R/V Vizconde de Eza during two different scientific surveys carried from mid-November to mid-December in both 2005 and 2006, in deep waters along the edge of the continental shelf of Morocco and Western Sahara (Figure 1). Fish were caught by a Lofoten type otter trawl with 17.70 m wing spread, 5.5 m vertical net opening and 35 mm cod-end mesh. The trawl had 250 m sweeps and the groundgear comprised 14″ bobbins, for trawling over rough grounds. The headline had high-strength floats (240 mm in diameter) to work at great depths. The trawl doors were oval type, and weighed 850 kg. The sampling design was random stratified based on depth and latitude (Figure 1). The planned hauls duration was 1 h, but varied from 45 to 60 min. In total, 95 hauls were conducted in 2005, and a further 99 in 2006. The mean depth of hauls (based on the average depth at the shoot and haul positions) ranged from 500–1860.5 m in 2005, and from 229–1845 m in 2006.

Figure 1. Map of the study area off Northwest Africa. Represented points indicate the hauls performed during both scientific surveys in 2005 and 2006 from North to South, respectively (created using Google Earth Pro© v. 7.3.6.9345 and Adobe Photoshop CS6 v. 13.0).
After the taxonomic identification, individuals of target and bycatch species were selected by length classes to complete the length range available. All specimens were measured for total length (TL, to 0.1 cm) and total weight (TW, to 1 g) and, when possible, they were sexed macroscopically.
LWRs were estimated by using the log-transformation (TW = log10 (a) + b log10 (TL)) of the power formula: TW = a × TLb, where a is the condition factor (related to body shape) and b is the allometry coefficient (related to the growth pattern) (Pauly, Reference Pauly1984). The 95% confidence interval and the standard error were also estimated for the LWR coefficients. The calculations were performed using the R-package FSA 0.9.4 (Ogle et al., Reference Ogle, Doll, Wheeler and Dinno2023; R-Core Team, 2023).
When LWRs were estimated by sex, the existence of statistically significant differences between the b-values was performed an ANCOVA test using GraphPad Prism® 10.1.2 software.
Results and discussion
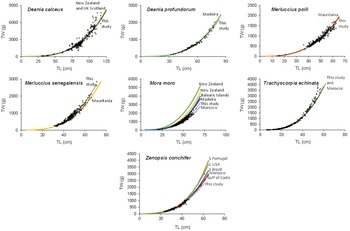
The species reported on here have no, or limited, LWRs in the FishBase database (Froese and Pauly, Reference Froese and Pauly2023) for either the eastern Atlantic (D. calceus and M. polli) or for Northwest Africa (other species). Results for the LWR for the species in this study are presented in Table 1 and Figure 2. Linear regressions provided suitable adjustments (r 2 ≥ 0.93), with b-values ranging from 2.638 and 3.172 for Z. conchifer and D. calceus, respectively.
Table 1. Length–weight relationships (LWR) for seven fish species caught off Atlantic Morocco and Western Sahara (NW Africa) by sex (F = females; M = males); N, sample size; TL, total length (cm); TW, Total Weight (g); a and b, LWR parameters; SE, standard error; CI, 95% confidence interval; r 2, determination coefficient; * no LWR references in FishBase (Froese and Pauly, Reference Froese and Pauly2023); ‡ Species with commercial interest; ¥ included in the IUCN red list


Figure 2. LWRs estimated for demersal species in Morocco and Western Sahara. Dots correspond to the length (TL)/total weight (TW) data pairs registered. Black line represents the LWRs estimated for the species in the present study. Other lines represent LWRs available in FishBase (Froese and Pauly, Reference Froese and Pauly2003), except for M. polli from Mauritania (Rey et al., Reference Rey, Fernández-Peralta, Quintanilla, Hidalgo, Presas, Salmeron and Puerto2015).
The present study provides new LWR data for seven data-limited demersal species inhabiting Atlantic Moroccan and Wester Saharan waters. Based on the Actinopterygii's information shared in FishBase (Froese and Pauly, Reference Froese and Pauly2023), this is the first LWR available for M. polli. However, previous studies provided LWR parameters (Rey et al., Reference Rey, Fernández-Peralta, Quintanilla, Hidalgo, Presas, Salmeron and Puerto2015) for both M. polli and M. senegalensis inhabiting Mauritanian waters, where these species support important fisheries and are assessed together in the FAO/CECAF framework (FAO, 2020). The TL ranges were wider in the study based on Mauritanian hakes (13.6–71.1 cm and 19.0–82.9 cm for M. polli and M. senegalensis, respectively), and the b-values are slightly different compared to the ones obtained in the present analyses. These differences could be explained by the very different environmental conditions present along the Mauritanian coast, where the water enrichment driven by the upwelling is much more noticeable. The b-values obtained for each sex in M. senegalensis resulted statistically similar (F = 0.57, P = 0.452). Regarding M. moro, T. echinata and Z. conchifer, the references available for the study area were only obtained for unsexed individuals (Meiners-Mandujano et al., Reference Meiners-Mandujano, Fernández-Peralta, Faraj and García-Cancela2018). In the present study, similar b-values were obtained for all the individuals analysed and the females of Z. conchifer (F = 0.006, P < 0.94) and between sexes in the case of T. echinata (F = 1.24, P = 0.266); however, for M. moro, significant differences exist between LWRs estimated by sex (F = 7.74, P < 0.01).
With regards the Elasmobranchii, although available LWRs for unsexed D. calceus correspond to long distance areas (i.e. Scotland and New Zealand), the very similar estimations could be explained because it is a cosmopolitan species inhabiting deep waters (usually between 400 and 1400 m depth) (Froese and Pauly, Reference Froese and Pauly2023), where the conditions are not so variable as in shallower and/or coastal waters. The same explanation could be used for D. profundorum, with similar LWR parameters estimated in Madeira (Ferreira et al., Reference Ferreira, Sousa, Delgado, Carvalho and Chada2008). Regarding the b-values estimated by sexes, significant differences exist in D. calceus (F = 37.88, P < 0.0001), but these values are statistically equal in D. profundorum (F = 0.0026, P = 0.96).
In the present study, two main constraints prevented from describing the growth patterns based on allometry in this study, such as the size range of fish caught and sample collection only occurring between November and December. Samples included the most common lengths observed in commercial and bycatch landings thus, LWR would be useful to assessment models; however, it should be noted the absence of smaller specimens, probably due to juveniles tending to occupy shallower waters than those surveyed. Furthermore, the limitation of sampling only to one season meant that no changes of seasonal energy investment between somatic growth and gonad maturation could be investigated (Rey et al., Reference Rey, Fernández-Peralta, Quintanilla, Hidalgo, Presas, Salmeron and Puerto2015). Additionally, other factors that could influence LWRs parameters, such as health or condition of the fish, and geographic and/or seasonal variations in the environmental conditions (Froese, Reference Froese2006), were not possible to assess in these analyses.
To conclude, it is well known that life history traits are used to delineate population geographical structures of species, due to they reflect genetic differences or environmental forced adaptations, and the LWRs determined here will be useful for future assessments of these fish species (Begg, Reference Begg, Cadrin, Friedland and Waldman2005; Gerritsen and McGrath, Reference Gerritsen and McGrath2007). However, it seems necessary to insist in the necessity to establish standardized methodologies agreed among the institutions involved in fishery research in the region, to be able to update and to monitor potential spatial and/or temporal variations, especially in a region greatly influenced by numerous anthropogenic pressures (i.e. fishing, industry, tourism, etc.) and environmental changes (major upwelling systems, and global warming, among others).
Acknowledgements
Authors thank all the scientific staff and the vessels' crews involved in the scientific surveys. Likewise, we thank Pablo Martín-Sosa Rodríguez for his help with R-packages.
Authors' Contributions
Both authors have participated in the data analysis and the manuscript writing and revision.
Financial Support
The present study was performed in the framework of a partnership programme between the Spanish Oceanographic Institute (IEO, CSIC) and the Moroccan ‘Institut National de Recherche Halieutique’ (INRH).
Competing interests
None.
Ethical Standards
This research was not covered by any regulation and formal ethical approval was not required. The scientific design was approved by the Spanish Fishery Office belonging to the National Government of Spain and the Kingdom of Morocco.
Availability of data and materials
Data available on request from the authors.



 Open access
Open access