



No CrossRef data available.
Article contents
Diet and food assimilation of the Plata pompano Trachinotus marginatus Cuvier, 1832 in a subtropical sandy beach inferred by stomach content and stable isotope analyzes
Published online by Cambridge University Press: 31 October 2023
Abstract
Investigating trophic linkages sustaining consumers is crucial to understanding their functional ecological role in communities and ecosystems. In this work, we combined stomach content (SCA) and stable isotope (SIA) analyses to investigate the trophic ecology of the Plata pompano Trachinotus marginatus during a critical phase of its life cycle along a subtropical sandy beach. This species is a conspicuous component of the southwestern Atlantic coast and commonly targeted by coastal fisheries. The diet was described using SCA, whereas the relative importance of food sources sustaining juveniles, as well their food niche structure and trophic position (TP), were evaluated using SIA. Juveniles consumed mainly crustaceans like the hippid crab Emerita brasiliensis, although other invertebrates (annelids, insects, molluscs) and fishes (including cannibalism) were also recorded. Although microcrustaceans dominated the diet, SIA showed that more palatable preys without carapaces or shells, like fishes and annelids, were the most assimilated preys in the muscle tissue of juvenile T. marginatus. There were marked changes in their isotopic niches (SEAc) and TP across ontogeny. SEAc ranged from 1.41‰2 for smaller individuals (0–40 TL mm) to 0.3‰2 for larger individuals (>80 TL mm). TP ranged from 3.1 (95% CI 2.7–3.6) for smaller to 4.5 (95% CI 3.9–5.1) for larger individuals. SIA suggest that juvenile T. marginatus derived most of their primary nutrients from a planktonic food web and, to a lesser extent, from a benthic pathway. Future studies are needed to better understand its functional role in food web of surf-zone ecosystems.
- Type
- Research Article
- Information
- Copyright
- Copyright © The Author(s), 2023. Published by Cambridge University Press on behalf of Marine Biological Association of the United Kingdom
References
Bastos, RF, Corrêa, F, Winemiller, KO and Garcia, AM (2017) Are you what you eat? Effects of trophic discrimination factors on estimates of food assimilation and trophic position with a new estimation method. Ecological Indicators 75, 234–241.CrossRefGoogle Scholar
Bastos, RF, Lippi, DL, Gaspar, ALB, Yogui, GT, Frédou, T, Garcia, AM and Ferreira, BP (2022) Ontogeny drives allochthonous trophic support of snappers: seascape connectivity along the mangrove-seagrass-coral reef continuum of a tropical marine protected area. Estuarine, Coastal and Shelf Science 264, 107591.CrossRefGoogle Scholar
Begon, M, Townsend, CR and Harper, JL (2006) Ecology: From Individuals to Ecosystems. Malden, USA: Blackwell.Google Scholar
Bergamino, L, Lercari, D and Defeo, O (2011) Food web structure of sandy beaches: temporal and spatial variation using stable isotope analysis. Estuarine, Coastal and Shelf Science 91, 536–543.CrossRefGoogle Scholar
Borcard, D, Gillet, F and Legendre, P (2018) Numerical Ecology with R, 2nd Edn. New York, NY: Springer.CrossRefGoogle Scholar
Calliari, LJ, Speranski, N, Torronteguy, M and Oliveira, MB (2001) The mud banks of Cassino Beach, Southern Brazil: characteristics, processes and effects. Journal of Coastal Research 34, 318–325.Google Scholar
Cansi, ER (2007) Comportamento de escape de Emerita brasiliensis (Crustacea, Anomura, Hippidae); Schmitt, 1935 (PhD thesis). Universidade de Brasília, Brasília, BR.Google Scholar
Condini, MV, Hoeinghaus, DJ and Garcia, AM (2015) Trophic ecology of dusky grouper Epinephelus marginatus (Actinopterygii, Epinephelidae) in littoral and neritic habitats of southern Brazil as elucidated by stomach contents and stable isotope analyses. Hydrobiologia 743, 109–125.CrossRefGoogle Scholar
Estes, JA, Terborgh, J, Brashares, JS, Power, ME, Berger, J, Bond, WJ, Carpenter, SR, Essington, TE, Holt, RD, Jackson, JBC, Marquis, RJ, Oksanen, T, Paine, RT, Pikitch, EK, Ripple, WJ, Sandin, SA, Scheffer, M, Schoener, TW, Shurin, JB, Sinclair, ARE, Soulé, ME, Virtanen, R and Wardle, DA (2011) Trophic downgrading of planet Earth. Science 333, 301–306.CrossRefGoogle ScholarPubMed
Fischer, LG, Pereira, LED and Vieira, JP (2011) Peixes Estuarinos e Costeiros, 2nd Edn. Rio Grande, RS: Gráfica Pallotti.Google Scholar
Garcia, AM, Claudino, MC, Mont'Alverne, R, Pereyra, PER, Copertino, M and Vieira, JP (2017a) Temporal variability in assimilation of basal food sources by an omnivorous fish at Patos Lagoon Estuary revealed by stable isotopes (2010–2014). Marine Biology Research 13, 98–107.CrossRefGoogle Scholar
Garcia, AM, Oliveira, MCLM, Odebrecht, C, Colling, JLA, Vieira, JP, Rodrigues, FL and Bastos, RF (2019b) Allochthonous vs autochthonous organic matter sustaining macroconsumers in a subtropical sandy beach revealed by stable isotopes. Marine Biology Research 15, 241–258.CrossRefGoogle Scholar
Garcia, AFS, Pasquaud, S, Cabral, H, Garcia, AM, dos Santos, ML and Vieira, JP (2019c) Assimilation of allochthonous matter by estuarine consumers during the 2015 El Niño event. Estuaries and Coasts 42, 1281–1296.CrossRefGoogle Scholar
Garcia, AF, Santos, ML, Garcia, AM and Vieira, JP (2019a) Changes in food web structure of fish assemblages along a river-to-ocean transect of a coastal subtropical system. Marine and Freshwater Research 70, 402–416.CrossRefGoogle Scholar
Garcia, AM, Winemiller, KO, Hoeinghaus, DJ, Claudino, MC, Bastos, R, Correa, , Huckembeck, FS, Vieira, JP, Loebmann, D, Abreu, PC and Ducatti, C (2017b). Hydrologic pulsing promotes spatial connectivity and food web subsidies in a subtropical coastal ecosystem. Marine Ecology Progress Series, 567, 17–28.CrossRefGoogle Scholar
Gianuca, NM (1983) A preliminary account of the ecology of sandy beaches in southern Brazil. In Mclachlan, A and Erasmus, T (eds), Sandy Beaches as Ecosystems. New York, NY: Springer, Dordrecht, pp. 413–419.CrossRefGoogle Scholar
Giménez, L and Yannicelli, B (2000) Longshore patterns of distribution of macroinfauna on a Uruguayan sandy beach: an analysis at different spatial scales and of their potential causes. Marine Ecology Progress Series 199, 111–125.CrossRefGoogle Scholar
Hairston, NG, Smith, FE and Slobodkin, SL (1960) Community structure, population control, and competition. The American Naturalist 94, 421–425.CrossRefGoogle Scholar
Hasenack, H and Ferraro, L (1989) Considerações Sobre o Clima da Região de Tramandaí, RS. Pesquisas em Geociências 22, 53–70.CrossRefGoogle Scholar
Helfman, G, Collette, BB, Facey, DE and Bowen, BW (2009) The Diversity of Fishes: Biology, Evolution, and Ecology. Chichester, UK: Wiley-Blackwell.Google Scholar
Hoffman, JC, Sierszen, ME and Cotter, AM (2015) Fish tissue lipid-C:N relationships for correcting δ13C values and estimating lipid content in aquatic food-web studies. Rapid Communications in Mass Spectrometry 29, 2069–2077.CrossRefGoogle Scholar
Huckembeck, S, Winemiller, KO, Loebmann, D and Garcia, AM (2020) Trophic structure of frog assemblages in coastal habitats in southern Brazil. Austral Ecology 45, 977–989.CrossRefGoogle Scholar
Hyslop, EJ (1980) Stomach contents analysis: a review of methods and their application. Journal of Fish Biology 17, 411–429.CrossRefGoogle Scholar
Jackson, AL, Inger, R, Parnell, AC and Bearhop, S (2011) Comparing isotopic niche widths among and within communities: SIBER – Stable Isotope Bayesian Ellipses in R. Journal of Animal Ecology 80, 595–602.CrossRefGoogle ScholarPubMed
Jepsen, DB and Winemiller, KO (2002) Structure of tropical river food webs revealed by stable isotope ratios. Oikos 96, 46–55.CrossRefGoogle Scholar
Kawakami, E and Vazzoler, G (1980) Método gráfico e estimativa de índice alimentar aplicado no estudo de alimentação de peixes. Boletim do Instituto Oceanográfico de São Paulo 29, 205–207.CrossRefGoogle Scholar
Layman, CA, Araujo, MS, Boucek, R, Harrison, E, Jud, ZR, Matich, P, Hammerschlag-Peyer, CM, Rosenblatt, AE, Vaudo, JJ, Yeager, LA, Post, DM and Bearhop, S (2012) Applying stable isotopes to examine food web structure: an overview of analytical tools. Biological Reviews 87, 542–562.CrossRefGoogle ScholarPubMed
Lazzari, S, Panizzi, AR and Grazia, J (2008) Insect drift and the case of Mayrinia curvidens (Mayr) (Hemiptera: Pentatomidae) drift on the southern Atlantic coast of Brazil. Neotropical Entomology 37, 109–117.CrossRefGoogle ScholarPubMed
Lemos, VM, Junior, ASV, Velasco, G and Vieira, JP (2011) The reproductive biology of the Plata pompano, Trachinotus marginatus (Teleostei: Carangidae), in southern Brazil. Zoologia 28, 603–609.CrossRefGoogle Scholar
Lima, MSP and Vieira, JP (2009) Variação espaço-temporal da ictiofauna da zona de arrebentação da Praia do Cassino, Rio Grande do Sul, Brasil. Zoologia 26, 499–510.CrossRefGoogle Scholar
Majdi, N, Hette-Tronquart, N, Auclair, E, Bec, A, Chouvelon, T, Cognie, B, Danger, M, Decottignies, P, Dessier, A, Desvilettes, C, Dubois, S, Dupuy, C, Fritsch, C, Gaucherel, C, Hedde, M, Jabot, F, Lefebvre, S, Marzloff, MP, Pey, B, Peyrard, N, Powolny, T, Sabbadin, R, Thebault, E and Perga, M (2018) There's no harm in having too much: a comprehensive toolbox of methods in trophic ecology. Fooweb 17, e00100. doi: 10.1016/j.fooweb.2018.e00100Google Scholar
Marques, WC, Fernandes, EH, Monteiro, IO and Möller, OO (2009) Numerical modeling of the Patos Lagoon coastal plume, Brazil. Continental Shelf Research 29, 556–571.CrossRefGoogle Scholar
McLachlan, A and Defeo, O (2017) The Ecology of Sandy Shores. California, CA: Academic Press.Google Scholar
Mittelbach, GG and McGill, BJ (2019) Community Ecology. New York, NY: Oxford University Press.CrossRefGoogle Scholar
Mont'Alverne, R, Pereyra, PER and Garcia, AM (2016) Trophic segregation of a fish assemblage along lateral depth gradients in a subtropical coastal lagoon revealed by stable isotope analyses. Journal of Fish Biology 89, 770–792.CrossRefGoogle Scholar
Monteiro-Neto, C and Cunha, LPR (1990) Seasonal and ontogenetic variation in food habits of juvenile Trachinotus marginatus (Cuvier, 1832) (Teleostei, Carangidae) in the surf zone of Cassino Beach, Rio Grande do Sul state, Brazil. Atlântica 12, 45–54.Google Scholar
Monteiro-Neto, C, Cunha, LPR and Musick, JA (2003) Community structure of surf-zone fishes at Cassino Beach, Rio Grande do Sul, Brazil. Journal of Coastal Research 35, 492–501.Google Scholar
Neves, LP, Silva, PSR and Bemvenuti, CE (2008) Temporal variability of benthic macrofauna on Cassino beach, southernmost Brazil. Iheringia, Série Zoologia 98, 36–44.CrossRefGoogle Scholar
Newsome, SD, Martinez del Rio, C, Bearhop, S and Phillips, DL (2007) A niche for isotopic ecology. Frontiers in Ecology and the Environment 5, 429–436.CrossRefGoogle Scholar
Niang, TMS, Pessanha, ALM and Araújo, FG (2010) Dieta de juvenis de Trachinotus carolinus (Actinopterygii, Carangidae) em praias arenosas na costa do Rio de Janeiro. Iheringia, Série Zoologia 100, 35–42.CrossRefGoogle Scholar
Nimer, E (1977) Região sul: clima. In IBGE (eds), Geografia do Brasil. Rio de Janeiro, RJ: IBGE, pp. 35–79.Google Scholar
Odebrecht, C, Bergesch, M, Rubi, RL and Abreu, PC (2010) Phytoplankton interannual variability at Cassino beach, Southern Brazil (1992–2007), with emphasis on the surf zone diatom Asterionellopsis glacialis. Estuaries and Coasts 33, 570–583.CrossRefGoogle Scholar
Oliveira, MCLM, Bastos, RF, Claudino, MC, Assumpção, CM and Garcia, AM (2014) Transport of marine-derived nutrients to subtropical freshwater food webs by juvenile mullets: a case study in southern Brazil. Aquatic Biology 20, 91–100.CrossRefGoogle Scholar
Oliveira, MCLM, Mont'Alverne, R, Sampaio, LA, Tesser, MB, Ramos, LRV and Garcia, AM (2017) Elemental turnover rates and trophic discrimination in juvenile Lebranche mullet Mugil liza under experimental conditions. Journal of Fish Biology 91, 1241–1249.CrossRefGoogle ScholarPubMed
Paine, RT (1969) The Pisaster-Tegula interaction: prey patches, predator food preference and intertidal community structure. Ecological Society of America 50, 950–961.Google Scholar
Parnell, A and Inger, R (2016) Stable isotope mixing models in R with simmr (R package version 0.3). Available at https://cran.rproject.org/web/packages/simmr/vignettes/simmr.html (last accessed 20 October 2020).Google Scholar
Paschoal, LRP (2013) Aspectos Gerais: Insetos e Reprodução. In Souza, CR, Lima, T, Paschoal, LRP and Piovezan, R (eds), Biologia da Reprodução de algumas ordens de inseto. Rio Claro, SP: Universidade Estadual Paulista “Júlio de Mesquita Filho”, Instituto de Biociências – Campus de Rio Claro (SP), p. 176.Google Scholar
Pereira, PS, Calliari, LJ and Barletta, RC (2010) Heterogeneity and homogeneity of Southern Brazilian beaches: a morphodynamic and statistical approach. Continental Shelf Research 30, 270–280.CrossRefGoogle Scholar
Peterson, BJ and Fry, B (1987) Stable isotopes in ecosystem studies. Annual Reviews of Ecology Evolution and Systematics 18, 293–320.CrossRefGoogle Scholar
Phillips, DL, Inger, R, Bearhop, S, Jackson, AL, Moore, JW, Parnell, AC, Semmens, BX and Ward, EJ (2014) Best practices for use of stable isotope mixing models in food-web studies. Canadian Journal of Zoology 92, 823–835.CrossRefGoogle Scholar
Pinotti, RM, Minasi, DM, Colling, LA and Bemvenuti, CE (2014) A review on macrobenthic trophic relationships along subtropical sandy shores in southernmost Brazil. Biota Neotropica 14, e20140069. doi: 10.1590/1676-06032014006914CrossRefGoogle Scholar
Polis, GA, Anderson, WB and Holt, RD (1997) Toward an integration of landscape ecology and food web ecology: the dynamics of spatially subsidized food webs. Annual Review of Ecology, Evolution, and Systematics 28, 289–316.CrossRefGoogle Scholar
Post, DM (2002) Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology 83, 703–718.CrossRefGoogle Scholar
Post, DM, Layman, CA, Arrington, DA, Takimoto, G, Quattrochi, J and Montana, CG (2007) Getting to the fat of the matter: models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia 152, 179–189.CrossRefGoogle Scholar
Quezada-Romegialli, C, Jackson, AL, Hayden, B, Kahilainen, KK, Lopes, C and Harrod, C (2018) tRophicPosition, an R package for the Bayesian estimation of trophic position from consumer stable isotope ratios. Methods in Ecology and Evolution 9, 1592–1599.CrossRefGoogle Scholar
R Core Team (2020) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria.Google Scholar
Rodrigues, FL, Cabral, HN and Vieira, JP (2015) Assessing surf-zone fish assemblage variability in southern Brazil. Marine and Freshwater Research 66, 106–119.CrossRefGoogle Scholar
Sampaio, LA, Tesser, MB and Burkert, D (2003) Tolerância de Juvenis do pampo Trachinotus marginatus (Teleostei, Carangidae) ao choque agudo de salinidade em laboratório. Ciência Rural, Santa Maria 33, 757–761.CrossRefGoogle Scholar
Santos, JNS (2010) Relação entre morfologia e dieta e uso da macroinfauna por pampos Trachinotus carolinus e Trachinotus goodei (Actinopterygii, Carangidae) em duas praias arenosas do sudeste do Brasil (PhD thesis). Universidade Federal Rural do Rio de Janeiro, Rio de Janeiro, BR.Google Scholar
Santos, ML and Vieira, JP (2016) A pesca com rede de cabo na Praia do Cassino, RS, Brasil. Boletim do Instituto de Pesca 42, 486–499.CrossRefGoogle Scholar
Savage, C, Thrush, SF, Lohrer, AM and Hewitt, JE (2012) Ecosystem services transcend boundaries: estuaries provide resource subsidies and influence functional diversity in coastal benthic communities. PLoS ONE 7, 1–8.CrossRefGoogle ScholarPubMed
Silva, PSR, Neves, LP and Bemvenuti, CE (2008) Temporal variation of sandy beach macrofauna at two sites with distinct environmental conditions on Cassino Beach, extreme Southern Brazil. Brazilian Journal of Oceanography 56, 257–270.CrossRefGoogle Scholar
Silveira, EL, Semmar, N, Cartes, JE, Tuset, VM, Lombarte, A, Ballester, ELC and Vaz-dos-Santos, AM (2020) Methods for trophic ecology assessment in fishes: a critical review of stomach analyses. Reviews in Fisheries Science & Aquaculture 28, 71–106.CrossRefGoogle Scholar
Toldo, EE Jr, Dillenburg, SR, Almeida, LESB, Tabajara, LL, Ferreira, ER and Borguetti, C (1993) Parâmetros Morfodinâmicos e Deriva Litorânea da Praia de Tramandaí, RS. GEOSUL 15, 75–88.Google Scholar
Tomazelli, LJ and Villwock, JA (1991) Geologia do Sistema Lagunar Holocênico do Litoral Norte do Rio Grande Do Sul, Brasil. Pesquisas 18, 13–24.Google Scholar
Vieira, J, Román-Robles, V, Rodrigues, F, Ramos, L and Santos, ML (2019) Long-term spatiotemporal variation in the juvenile fish assemblage of the Tramandaí River estuary (29° S) and adjacent coast in southern Brazil. Frontiers in Marine Science 6, 269.CrossRefGoogle Scholar
Vollrath, SR, Possamai, B, Schneck, F, Hoeinghaus, DJ, Albertoni, EF and Garcia, AM (2021) Trophic niches and diet shifts of juvenile mullet species coexisting in marine and estuarine habitats. Journal of the Marine Biological Association of the United Kingdom 101, 431–441.CrossRefGoogle Scholar
Winemiller, KO and Layman, CA (2005) Food web science: moving on the path from abstraction to prediction. In Ruiter, PC, Wolters, V and Moore, JC (eds), Dynamic Food Webs: Multispecies Assemblages, Ecosystem Development and Environmental Change. Amsterdam, NL: Elsevier, pp. 10–23.CrossRefGoogle Scholar
Winemiller, KO, Zeug, SC, Robertson, CR, Winemiller, BK and Honeycutt, RL (2011) Food-web structure of coastal streams in Costa Rica revealed by dietary and stable isotope analyses. Journal of Tropical Ecology 27, 463–476.CrossRefGoogle Scholar
Zaret, TM and Paine, RT (1973) Species introduction in a tropical lake. Science 182, 449–455.CrossRefGoogle Scholar

Araújo et al. supplementary material
Araújo et al. supplementary material 1

File
18 KB

Araújo et al. supplementary material
Araújo et al. supplementary material 2
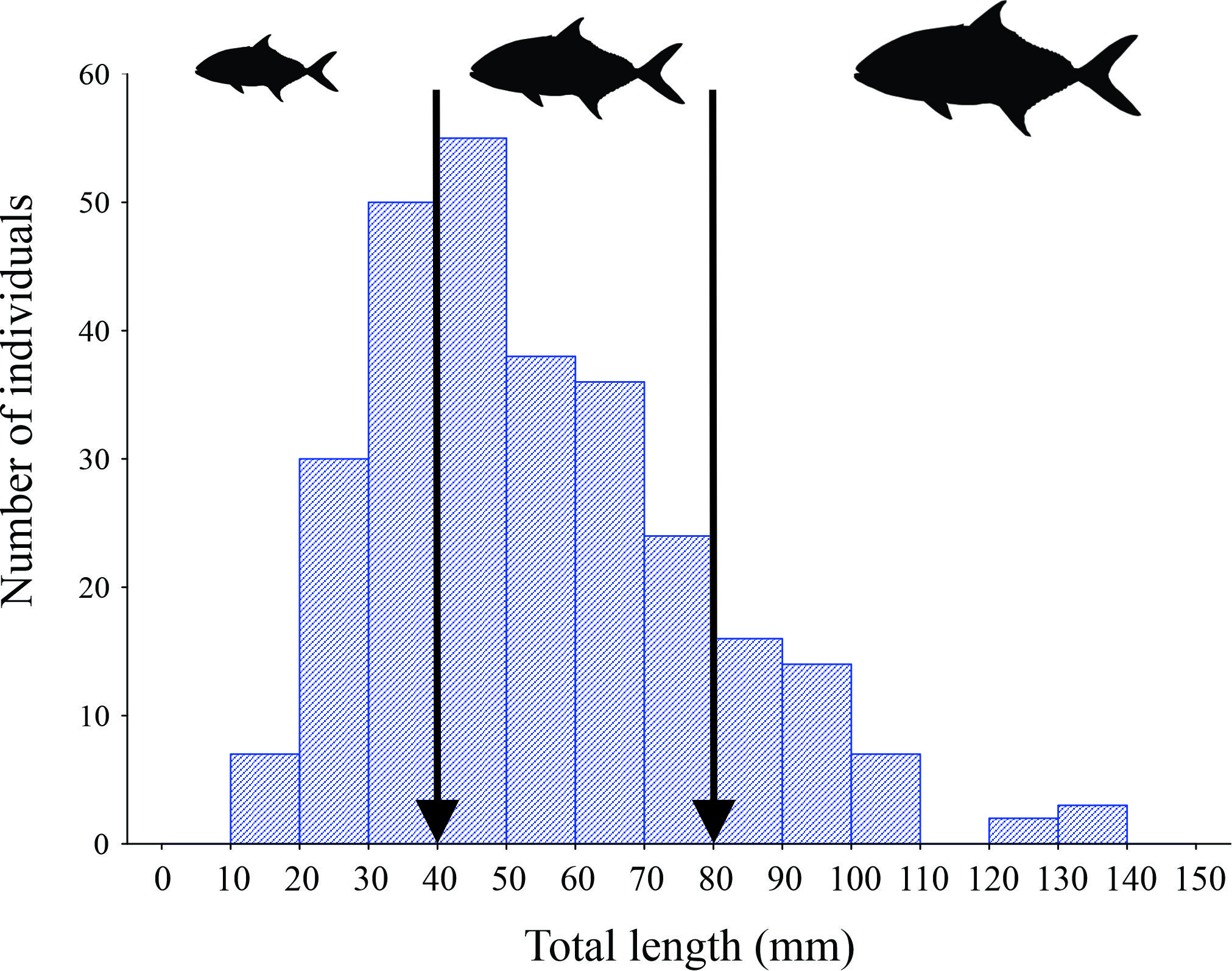
Image
1.8 MB
Araújo et al. supplementary material
Araújo et al. supplementary material 3
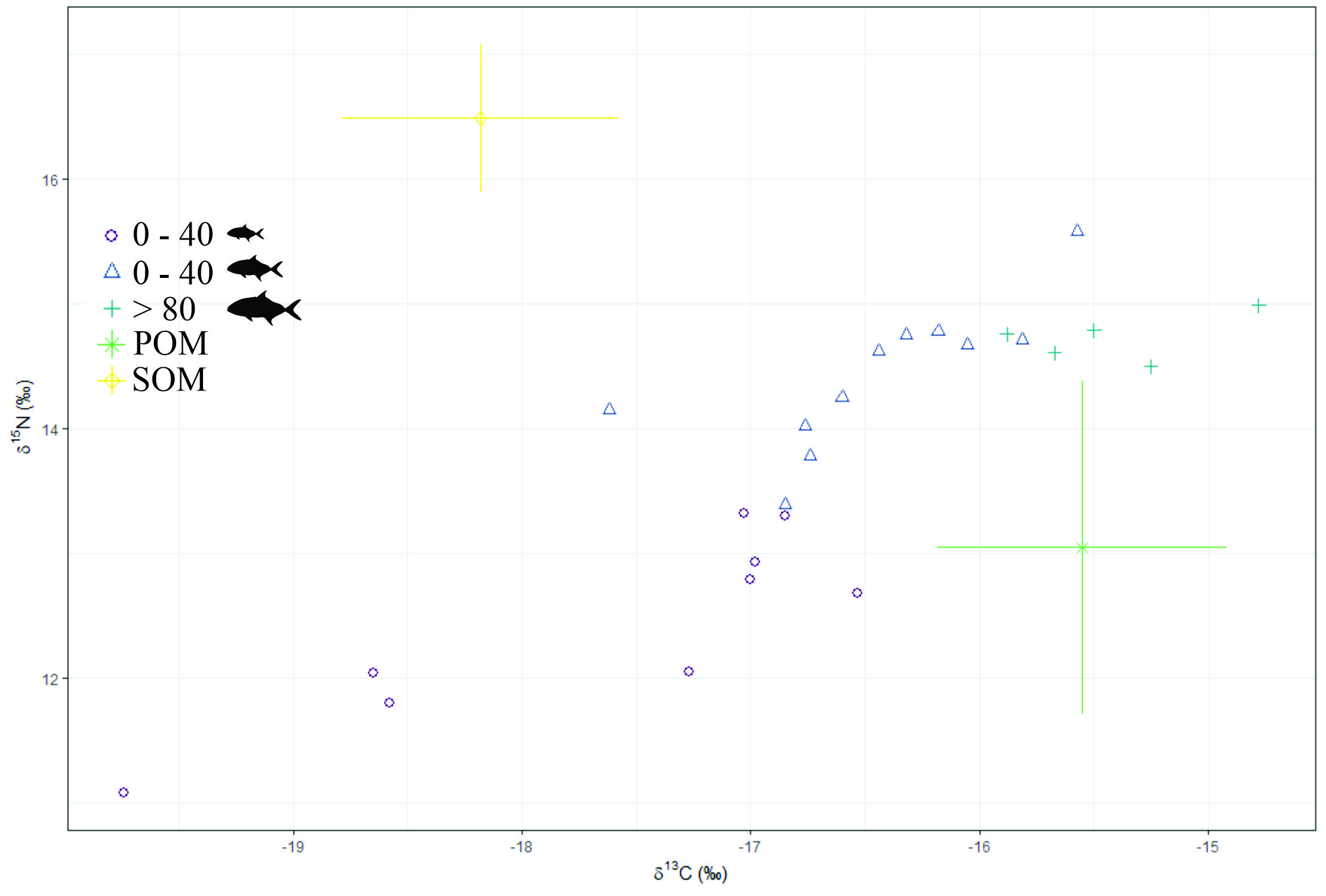
Image
884 KB
Araújo et al. supplementary material
Araújo et al. supplementary material 4
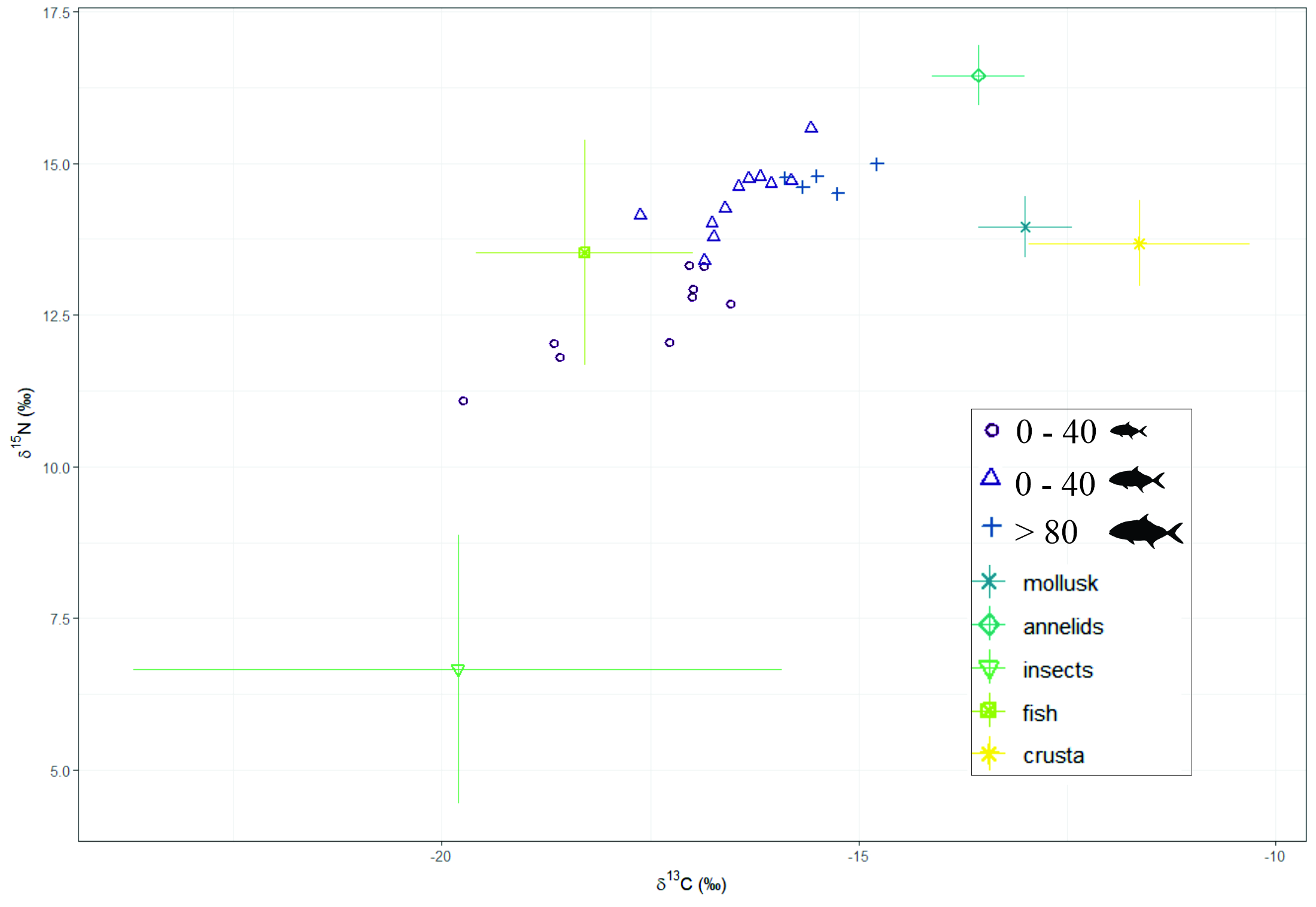
Image
1.2 MB