



Non-technical Summary
Some trilobite species lived in deeper-water environments that are not well preserved in the fossil record. There are also intervals of time that are not well represented in some areas. This is the case for the earliest part of the Ordovician in Morocco. Because of this, any well-preserved trilobite specimens from these rocks are important for understanding how they relate to other parts of the world. They are also important for our understanding of migration and evolution of trilobites during this time. Here we present the first well-preserved trilobites of the family Olenidae from Morocco. Species from this family were often restricted to deeper or low-oxygen environments. This is the first discovery of olenid trilobites from Africa that are well preserved enough to be able to identify the species.
Introduction
The early Paleozoic rock record of Morocco is predominantly marine and highly fossiliferous. Among Ordovician deposits, the Fezouata Shale Formation is notable for its diverse fossil fauna, including two intervals (upper Tremadocian and mid-Floian interval) with exceptional preservation (Lefebvre et al., Reference Lefebvre, Gutiérrez-Marco, Lehnert, Martin, Nowak, Akodad, El Hariri and Servais2018). In contrast, the lowermost Fezouata Shale Formation (lower Tremadocian) is relatively unfossiliferous: in the last 60−70 years, only a handful of marine invertebrate specimens have been recovered from the lowest 100 meters of the more than 800 m that the formation reaches in the depocenter of the central Anti-Atlas Mountains (Lefebvre et al., Reference Lefebvre, Allaire, Guensburg, Hunter, Kouraïss, Martin, Nardin, Noailles, Pittet and Sumrall2016a, Reference Lefebvre, Gutiérrez-Marco, Lehnert, Martin, Nowak, Akodad, El Hariri and Servais2018). The fossil material consists primarily of graptolites and rare trilobite specimens referred to as Beltella sp. and Pharostomina sp. (Destombes, Reference Destombes1967, Reference Destombes2006c; Destombes et al., Reference Destombes, Hollard, Willefert and Holland1985; Lefebvre et al., Reference Lefebvre, Allaire, Guensburg, Hunter, Kouraïss, Martin, Nardin, Noailles, Pittet and Sumrall2016a). Further, across Morocco, the early Tremadocian is represented only by the lowermost Fezouata Shale Formation in the Anti-Atlas area (Destombes et al., Reference Destombes, Hollard, Willefert and Holland1985). Thus, any new discoveries of well-preserved specimens from this unit can have particular value on our understanding of the earliest Ordovician faunas (e.g., Lefebvre et al., Reference Lefebvre, Allaire, Guensburg, Hunter, Kouraïss, Martin, Nardin, Noailles, Pittet and Sumrall2016a). In this paper, we describe the first well-preserved olenid trilobite specimens recovered from this unit. These are also the first olenid trilobite specimens well preserved enough to be identified to species level from the African continent.
Geologic setting
The specimens were found by locals near N'kob village (Fig. 1.1) and acquired in 2019. In the N'kob area, the Fezouata Shale Formation crops out extensively across the southern flank of the Precambrian antiform of the Jbel Saghro (also known as the Saghro Massif), but there are no complete stratigraphic sections because the unit is regularly covered by Quaternary sediments. Partial exposures exist on hillsides, where they are topped with Pleistocene terrace deposits (Fig. 1.2). In the plains, Holocene deposits are thin enough (1 meter or less) that any small excavation has the potential to expose Fezouata Shale Formation layers. Although the exact location where these specimens were found is unknown, outcrops of the Fezouata Shale Formation around N'kob preserve the graptolites Rhabdinopora canadensis (Lapworth, Reference Lapworth1898) and R. socialis (Salter, Reference Salter1858), among other graptolites, in the lowermost 6-15 m, followed by sporadic occurrences of R. flabelliformis (Eichwald, Reference Eichwald1840) in the succeeding 50 meters of the Fezouata Shale Formation (Destombes and Willefert, Reference Destombes and Willefert1959; reviewed by Gutiérrez-Marco and Martin, Reference Gutiérrez-Marco and Martin2016; with taxonomy updated according to Maletz et al., Reference Maletz, Wang and Wang2023).
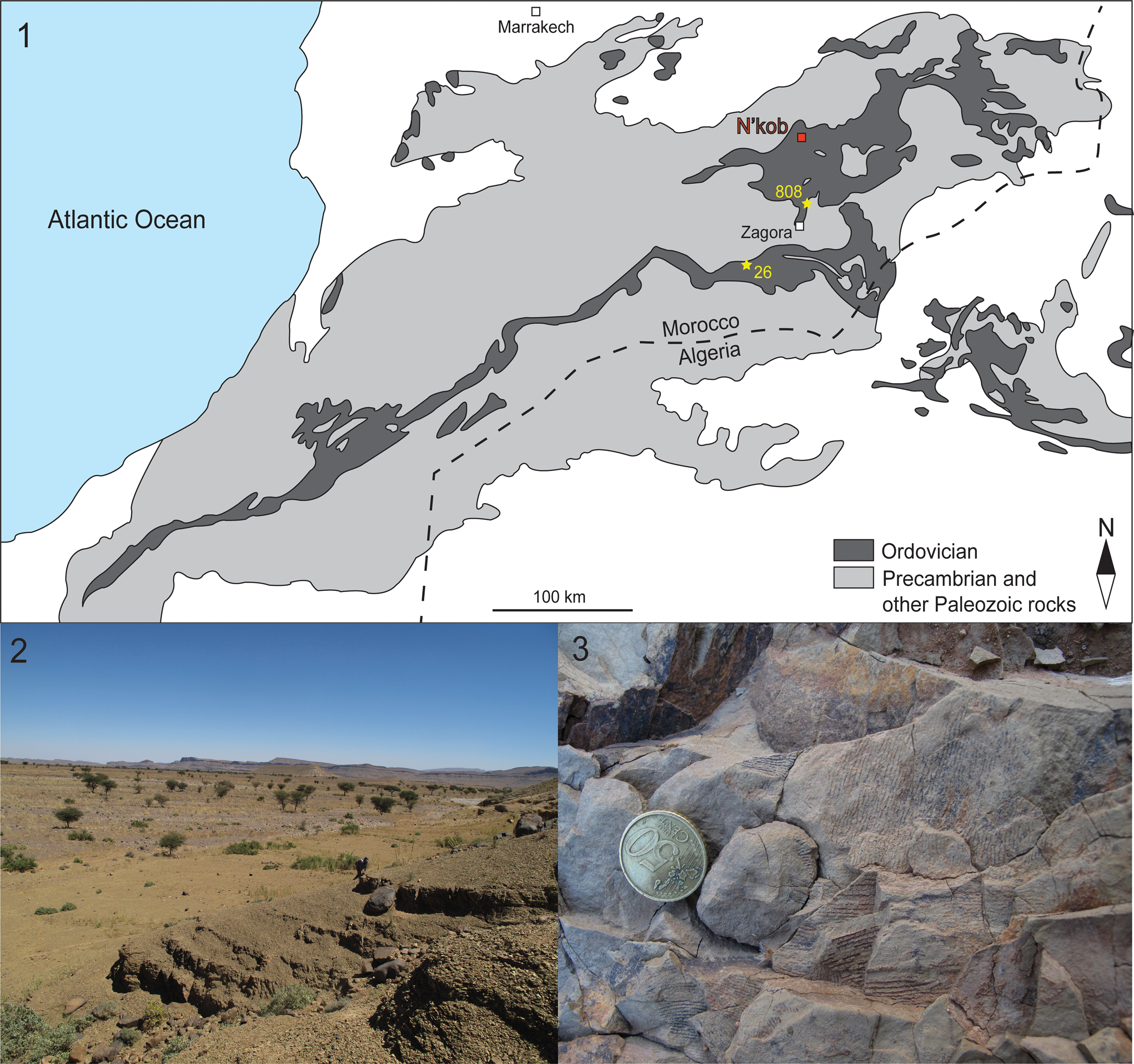
Figure 1. Geographic setting. (1) Simplified geologic map of the Anti-Atlas Mountains and localities of recovered olenid trilobites in Morocco. Previously reported specimens identified as Beltella sp. were found at localities 26 and 808 (yellow stars) in the Zagora area (Destombes, Reference Destombes2006c). The specimens described in this paper were found in the N'kob area (red square). Modified from Álvaro et al. (Reference Álvaro, Benharref, Destombes, Gutiérrez-Marco, Hunter, Lefebvre, van Roy, Zamora, Hunter, Álvaro, Lefebvre, van Roy and Zamora2022). (2) Lower Tremadocian exposures of the Fezouata Shale Formation in the N'kob area. Mountainous area in background represents the First Bani Group. (3) Bedding-plane accumulations of the graptolite Rhabdinopora cf. R. socialis (Salter) in the Akka n'Ousdidene locality east of N'kob, ~12 m above the base of the Fezouata Shale Formation. Diameter of coin is 24 mm.
Materials and methods
The specimens are preserved in a light tan, non-calcareous, micaceous argillite. The fossil material is likely recrystallized, brown calcite, that strongly stands out visually against the matrix. While the exoskeletal surface preserves small details of ornamentation, it is also characterized by a network of fine fractures and the exoskeleton fragments easily. The specimens were prepared using air abrasion with a dolomite–calcium-carbonate powder blend. Four exoskeletons of varying completeness were recovered, three of which are clustered on top of one another (Fig. 2). Although mostly articulated, displacement of librigenae indicates that these specimens are molts of roughly the same size (the length of the cranidia varies from 6.8 to 7.7 mm).
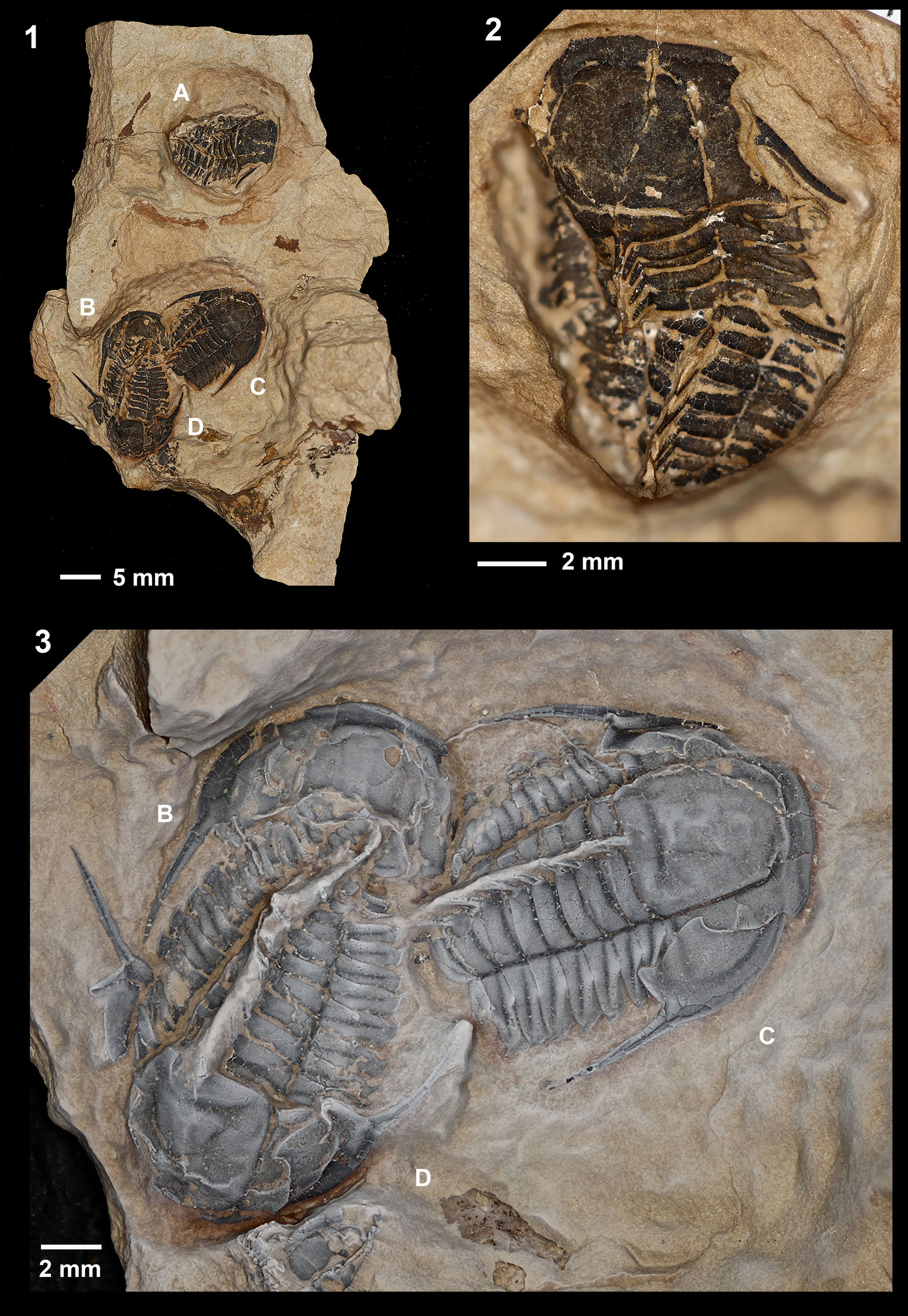
Figure 2. Leptoplastides salteri (Callaway, Reference Callaway1877) specimens from N'kob, Morocco. (1) Entire rock sample showing all four specimens labeled A–D to correspond to (2, 3). (2) Part of rock sample showing AMNH-FI-139288 (labeled “A”). (3) Part of rock sample showing AMNH-FI-139289 (labeled “B”), AMNH-FI-139290 (labeled “C”), and AMNH-FI-139291 (labeled “D”); (1, 3) whitened with ammonium chloride. Scale bars below associated images.
Repository and institutional abbreviation
Specimens examined and figured in this study are deposited in the American Museum of Natural History (AMNH), New York, USA, under catalog numbers AMNH-FI-139288 to AMNH-FI-139291.
Results
The Moroccan material fits the diagnosis of the genus Leptoplastides Raw, Reference Raw1908 (Nikolaisen and Henningsmoen, Reference Nikolaisen and Henningsmoen1985, p. 12) and, among the species defined for the genus, it most closely resembles Leptoplastides salteri (Callaway, Reference Callaway1877). Articulated specimens, including meraspides, of L. salteri (Callaway, Reference Callaway1877) have been figured and described by Lake (Reference Lake1919, p. 89−93, pl. 11, figs. 2−5), Raw (Reference Raw1925, p. 229−247, pls. 16−18), Fortey and Owens (Reference Fortey and Owens1991, p. 449−451, fig. 8c−j), and Månsson and Clarkson (Reference Månsson and Clarkson2020, p. 2−15, figs. 1−11). Features that align the newly recovered Moroccan specimens with L. salteri include the mid-axial spines on the occipital ring and thoracic tergites (Figs. 2.3, 3.1) but not the pygidium (Fig. 3.3), the small anteriorly placed palpebral lobes (Fig. 2.2), the narrow, relatively long genal spines emerging from the lateral margin of the cephalon (Figs. 2.3, 3.2), the spinose thoracic pleura (Fig. 3.2), and the small spinose projection at the antero-lateral angle of pygidium (Fig. 3.3).
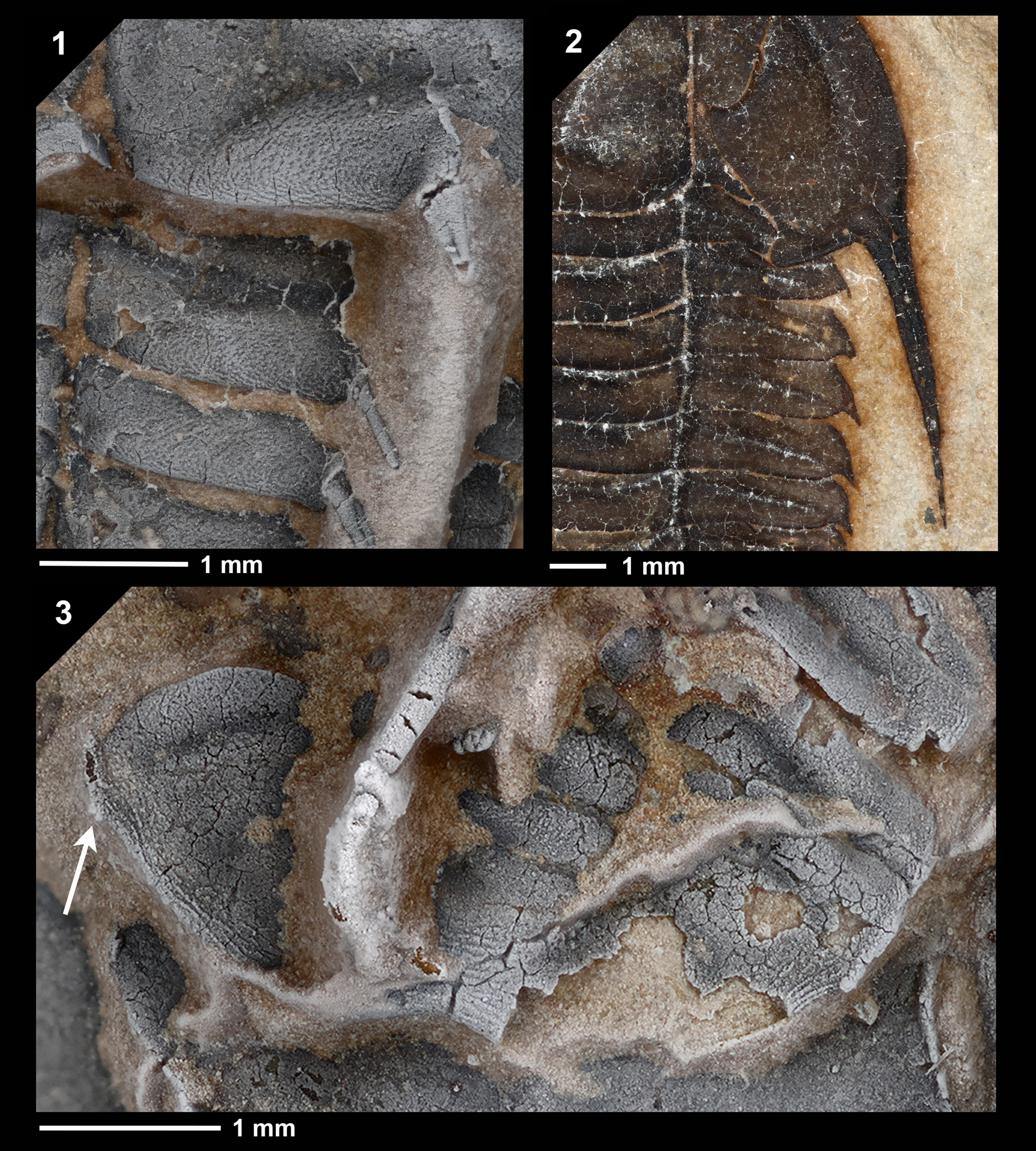
Figure 3. Close-up images of Leptoplastides salteri (Callaway, Reference Callaway1877) specimens, whitened in (1, 3) with ammonium chloride to show sculpture. (1) Close-up of occipital ring and two anteriormost axial rings of the thorax, showing pitting sculpture and axial spines, AMNH-FI-139291 (labeled “D” in Fig. 2A). (2) Close-up of genal spine and pleural region of thorax, AMNH-FI-139288 (labeled “A” in Fig. 2A). (3) Close-up of pygidium, AMNH-FI-139291 (labeled “D” in Fig. 2A). Arrow points to a small spinose projection at the antero–lateral angle. Scale bars below associated images.
The specimens presented here have two axial rings and a terminal piece (Fig. 3.3). This would seem in contrast to the description of L. salteri provided by Lake (Reference Lake1919), which stated that the axis is divided into three distinct rings and a terminal portion. However, Lake's illustrations clearly show specimens with two rings and a terminal piece (e.g., Lake, Reference Lake1919, pl. 11, fig. 3), as do the specimens figured in Raw (Reference Raw1925) and Fortey and Owens (Reference Fortey and Owens1991), including the lectotype, which is housed in the Birmingham University Collections (BU691, see Fortey and Owens, Reference Fortey and Owens1991, fig. 8h).
Raw (Reference Raw1925) noted that L. salteri transitioned to the holaspid stage at a body size of about 5.75 mm and that disarticulated material indicated a maximum size of 60 mm. He reported ongoing ontogenetic changes during the holaspid period, including (1) increasing width-to-length ratio of the glabella; (2) increasing effacement of glabellar furrows; (3) decrease in prominence of the ocular ridge; (4) increasingly posterior position of the genal spine; (5) increase in the number of thoracic segments with “unguiculate” (= spiny or pointed) pleural terminations from anterior to posterior; (6) diminishing size of the pleural projection at the antero–lateral angle of the pygidium; (7) increasing emargination of the posterior border of the terminal piece; and (8) increasingly concave posterior border of the pygidium. The Moroccan specimens are more than twice as large as the previously figured articulated specimens. For example, AMNH-FI-139291 (Fig. 2.3, specimen D) is 16.58 mm in length, while the largest articulated specimens figured by Månsson and Clarkson, Reference Månsson and Clarkson2020, fig. 8e, and Fortey and Owens, Reference Fortey and Owens1991, fig. 8j, are 7.1 mm and 6.8 mm, respectively, and the differences between them and previously figured specimens are consistent with the ontogenetic changes described by Raw (Reference Raw1925).
Although displacement complicates comparisons, the genal spines in the Moroccan specimens appear to be relatively longer than those of smaller specimens figured elsewhere. The occipital furrows are more deeply expressed medially than laterally in the Moroccan specimens. It is not clear if this is taphonomic (for example, this is seen in some of the more compressed specimens from England [e.g., Fortey and Owens, Reference Fortey and Owens1991, figs. 8c–e]), ontogenetic, or both.
Additional morphological details that are expressed on the Moroccan specimens include fine pitting on the axial lobes of the cephalon and thorax (Fig. 3.1) and terrace lines along the margin of the pygidium (Fig. 3.3).
Combinations of these features distinguish the UK and Moroccan specimens from other Leptoplastides species (see also Table 1).
Table 1. Summary of species assigned to the genus Leptoplastides, including references supporting current designations, geographic occurrence, and stratigraphic age. The last four rows list species that are not currently assigned to Leptoplastides but at one point were designated as Beltella.
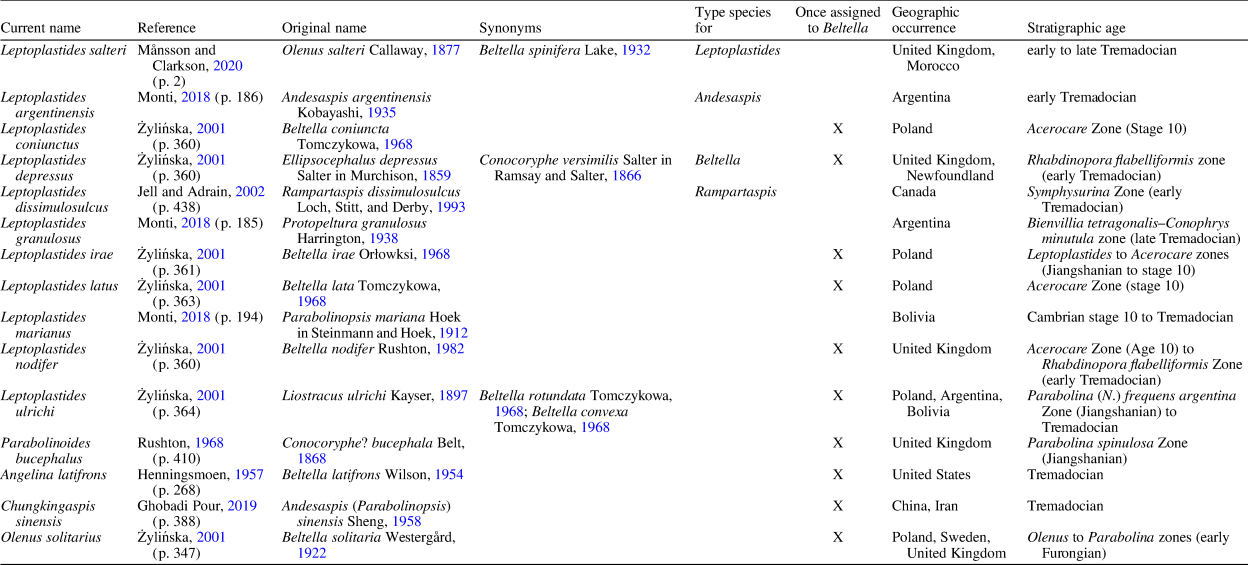
Leptoplastides argentinensis (Kobayashi, Reference Kobayashi1935) has a longer (sag.) preglabellar field, smaller palpebral lobes, no intergenal angle, and more developed pygidial marginal spines (see Monti, Reference Monti2018).
Leptoplastides coniunctus (Tomczykowa, Reference Tomczykowa1968) has transglabellar furrows, no (apparent) axial spines or nodes on the thorax, shallower intergenal angle, and shorter genal spine (see Żylińska, Reference Żylińska2001).
Leptoplastides depressus (Salter in Murchison, Reference Murchison1859) has no axial spines or nodes and blunt pleural ends on the thorax (see Fortey and Owens, Reference Fortey and Owens1989).
Leptoplastides dissimulosulcus (Loch et al., Reference Loch, Stitt and Derby1993) has a longer (sag.) preglabellar field and wider librigenal field. The thorax is unknown, and the pygidium is too incomplete for adequate comparison, but the occipital ring bears what may be the base of a robust spine (see Loch et al., Reference Loch, Stitt and Derby1993).
Leptoplastides granulosus (Harrington, Reference Harrington1938) has a narrower (sag. and tr.) anterior border, shorter, more anteriorly placed palpebral lobes, shallower genal angle, thoracic axial nodes not developed into spines, and granular ornamentation (see Waisfeld and Vaccari, Reference Waisfeld, Vaccari and Benedetto2003; Monti, Reference Monti2018).
Leptoplastides irae (Orłowski, Reference Orłowski1968) has no intergenal angle, shorter genal spines, no axial spine or nodes, and two short marginal pygidial spines (see Żylińska, Reference Żylińska2001).
Leptoplastides latus (Tomczykowa, Reference Tomczykowa1968) has an axial node (instead of long spine) on the occipital ring and at least two short marginal pygidial spines; the thorax is unknown although comparable specimens from Norway show nodes only on the anteriormost thoracic tergites (see Nikolaisen and Henningsmoen, Reference Nikolaisen and Henningsmoen1985; Żylińska, Reference Żylińska2001).
Leptoplastides marianus (Hoek in Steinmann and Hoek, Reference Steinmann and Hoek1912) was recently restricted to the type specimens (Monti, Reference Monti2018), which have no apparent axial spines or nodes and a shallower genal angle. Comparison is hampered by extreme distortion of the specimens, which has resulted in a complicated taxonomic history for this and other Argentinian species (see Waisfeld and Vaccari, Reference Waisfeld, Vaccari and Benedetto2003; Monti, Reference Monti2018).
Leptoplastides nodifer (Rushton, Reference Rushton, Bassett and Dean1982) has a longer (sag.) preglabellar field, shallower intergenal angle, and axial nodes along the thorax but they are not developed into spines (see Rushton, Reference Rushton, Bassett and Dean1982).
Leptoplastides ulrichi (Kayser, Reference Kayser1897) has no apparent axial nodes or spines, short spines on anteriormost thoracic tergites but blunt pleural ends otherwise, and a shallower intergenal angle (see Harrington and Leanza, Reference Harrington and Leanza1957; Żylińska, Reference Żylińska2001).
Discussion
Comparison with other Moroccan olenid specimens
Previously, the only olenid trilobite to have been reported from Morocco was Beltella sp., which has been mentioned frequently in recent descriptions of the lowermost (lower Tremadocian) Fezouata Shale Formation (e.g., Destombes et al., Reference Destombes, Hollard, Willefert and Holland1985; Destombes and Feist, Reference Destombes and Feist1987; Lefebvre et al., Reference Lefebvre, Allaire, Guensburg, Hunter, Kouraïss, Martin, Nardin, Noailles, Pittet and Sumrall2016a, Reference Lefebvre, El Hariri, Lerosey-Aubril, Servais and Van Royb). However, this occurrence is based on the only two specimens known from extensive collections made by Destombes (Lefebvre et al., Reference Lefebvre, El Hariri, Lerosey-Aubril, Servais and Van Roy2016b). These specimens include an internal mold of a cranidium with four partial thoracic tergites and an internal mold of a pygidium and partial thorax (Destombes, Reference Destombes2006c, pl. 1, figs. 1, 2). These two specimens were collected from localities 808 and 26 in the Zagora area (Destombes, Reference Destombes2006b) in 1959 and 1961, respectively (Destombes, personal communication, 2009) (Fig. 1.1), about 50−100 km south of the N'kob area. Although one of us (JCG-M) has been to locality 808, no new olenid trilobites were recovered.
Although the specimens figured by Destombes are not complete, there is no aspect of the preserved exoskeleton that provides conclusive evidence that these are not L. salteri. Destombes may have assigned these two specimens to Beltella because neither clearly show axial spines. However, previous workers noted that the spines are not well preserved on molds, especially on convex internal molds, and the axial rings can appear smooth (Lake, Reference Lake1919); the spines also break easily (Månsson and Clarkson, Reference Månsson and Clarkson2020). There is at least one axial ring with what could be the base of a spine on the cranidium with partial thoracic tergites (Destombes, Reference Destombes2006c, pl. 1, fig. 1).
Because he did not find any “Beltella” specimens or acritarchs in areas farther north of Zagora, Destombes (Reference Destombes2006a, p. 17) suggested that the lowermost Fezouata Shale Formation was missing from those areas, including the region around N'kob. Recent work on graptolites refutes this suggestion (Gutiérrez-Marco and Martin, Reference Gutiérrez-Marco and Martin2016); furthermore, it is not surprising that Destombes did not find olenid trilobites near N'kob given how rarely they appear in the Zagora area.
Taxonomic history of the genus Leptoplastides.—There are currently 11 species assigned to the genus Leptoplastides (Table 1). Perhaps the most controversially assigned species are those formerly assigned to the genus Beltella. In contrast to L. salteri, Beltella depressa (Salter in Murchison, Reference Murchison1859), the type species of Beltella, was known only from deformed, disarticulated specimens until the end of the twentieth century when well-preserved specimens were described by Fortey and Owens (Reference Fortey and Owens1989) from material recovered from temporary excavations in Gloucestershire, UK. Fortey and Owens (Reference Fortey and Owens1989) argued that each genus should be retained, contrary to previous opinions to synonymize the two (Henningsmoen, Reference Henningsmoen1957; Nikolaisen and Henningsmoen, Reference Nikolaisen and Henningsmoen1985; Morris, Reference Morris1988). The primary distinctions made were the presence of axial spines or nodes and posterolateral pleural spines on Leptoplastides and the absence of both on Beltella. However, Żylińska (Reference Żylińska2001) noted that the presence of these features is not restricted to species of Leptoplastides and synonymized them on this basis as well as the observation that pleural spines were lost in some olenid trilobite species during ontogeny (Whittington, Reference Whittington1992, Reference Whittington1996), a view that we share.
Opinion has varied over whether Andesaspis argentinensis Kobayashi, Reference Kobayashi1935, should be synonymized with Leptoplastides marianus (Hoek in Steinmann and Hoek, Reference Steinmann and Hoek1912) or treated as a separate species of Leptoplastides (e.g., Henningsmoen, Reference Henningsmoen1957; Waisfeld and Vaccari, Reference Waisfeld, Vaccari and Benedetto2003; Tortello and Esteban, Reference Tortello and Esteban2016), with the most recent treatment providing morphometric evidence for their separation with L. marianus restricted to the type specimens (Monti, Reference Monti2018; see also Waisfeld and Vaccari, Reference Waisfeld, Vaccari and Benedetto2003). Jell and Adrain (Reference Jell and Adrain2002) synonymized Rampartaspis Loch in Loch et al., Reference Loch, Stitt and Derby1993, with Leptoplastides without explanation.
Monti and Confalonieri (Reference Monti and Confalonieri2018) published the first attempt at a family-wide phylogenetic analysis of olenid trilobites. The analysis included 58 species, five of which belong to Leptoplastides sensu Żylińska (Reference Żylińska2001), and the resulting maximum parsimony tree indicated that those species comprise a polyphyletic group. However, recent explorations of the same matrix modified for analysis using a Bayesian approach frequently recovered trees where included Leptoplastides species formed a monophyletic clade (Wright and Hopkins, Reference Wright and Hopkins2021). Nonetheless, different treatments also identified different distributions of trees that vary considerably in their topologies (Wright and Hopkins, Reference Wright and Hopkins2021), so our understanding of genera and subfamilies within Olenidae remains in flux (see also Monti et al., Reference Monti, Tortello and Confalonieri2022).
Spatial and temporal extent of Leptoplastides.—The 11 species currently assigned to Leptoplastides range from the Jiangshanian (e.g., Leptoplastides irae [Orłowski, Reference Orłowski1968] from the Holy Cross Mountains in Poland; see Żylińska, Reference Żylińska2001) into the Tremadocian. Species occur in deeper-subtidal to offshore deposits of Gondwana (Argentina, Bolivia, southern Mexico, Morocco, Poland, Spain?), Baltica (Norway), British Avalonia, and Laurentia (Canada, Texas) (Fig. 4). The occurrences of Leptoplastides in Laurentia appear to belie previous compilations indicating provincialization of trilobite faunas in the Early Ordovician (Fortey and Cocks Reference Fortey and Cocks2003; Torsvik and Cocks, Reference Torsvik and Cocks2017, p. 113). Further, the occurrence of L. argentinensis in deeper-water basin sequences of Texas is reported alongside at least one other olenid genus (Angelina) known from Argentina (Wilson, Reference Wilson1954), implying some connection between South American Gondwana and the southern coast of Laurentia. However, the Mexican occurrences are from the Tiñú Formation (Robison and Pantoja-Alor, Reference Robison and Pantoja-Alor1968), which is considered to belong to a terrane (Oaxaquia) that, although accreted to North America, originally was either an insular area in temperate latitudes or was situated in a Gondwanan or peri-Gondwanan position in pre-Silurian times (Landing et al., Reference Landing, Westrop and Keppie2007; Torsvik and Cocks, Reference Torsvik and Cocks2017, p. 104; Cuen-Romero et al., Reference Cuen-Romero, Buitrón-Sánchez, Beresi, Palafox-Reyes, Monreal, Servais, Harper, Lefebvre and Percival2023), even though Mexico is reconstructed adjacent to Laurentia in the PALEOMAP models (Scotese, Reference Scotese2023). Notably the only other trilobite genus reported from the lowermost Fezouata Shale Formation, the phacopide Pharostomina, also co-occurs with Leptoplastides in the Tiñú Formation (Robison and Pantoja-Alor, Reference Robison and Pantoja-Alor1968), various formations of Argentina (Balseiro et al., Reference Balseiro, Waisfeld and Vaccari2011), and Wales (Owens et al., Reference Owens, Fortey, Cope, Rushton and Bassett1982).
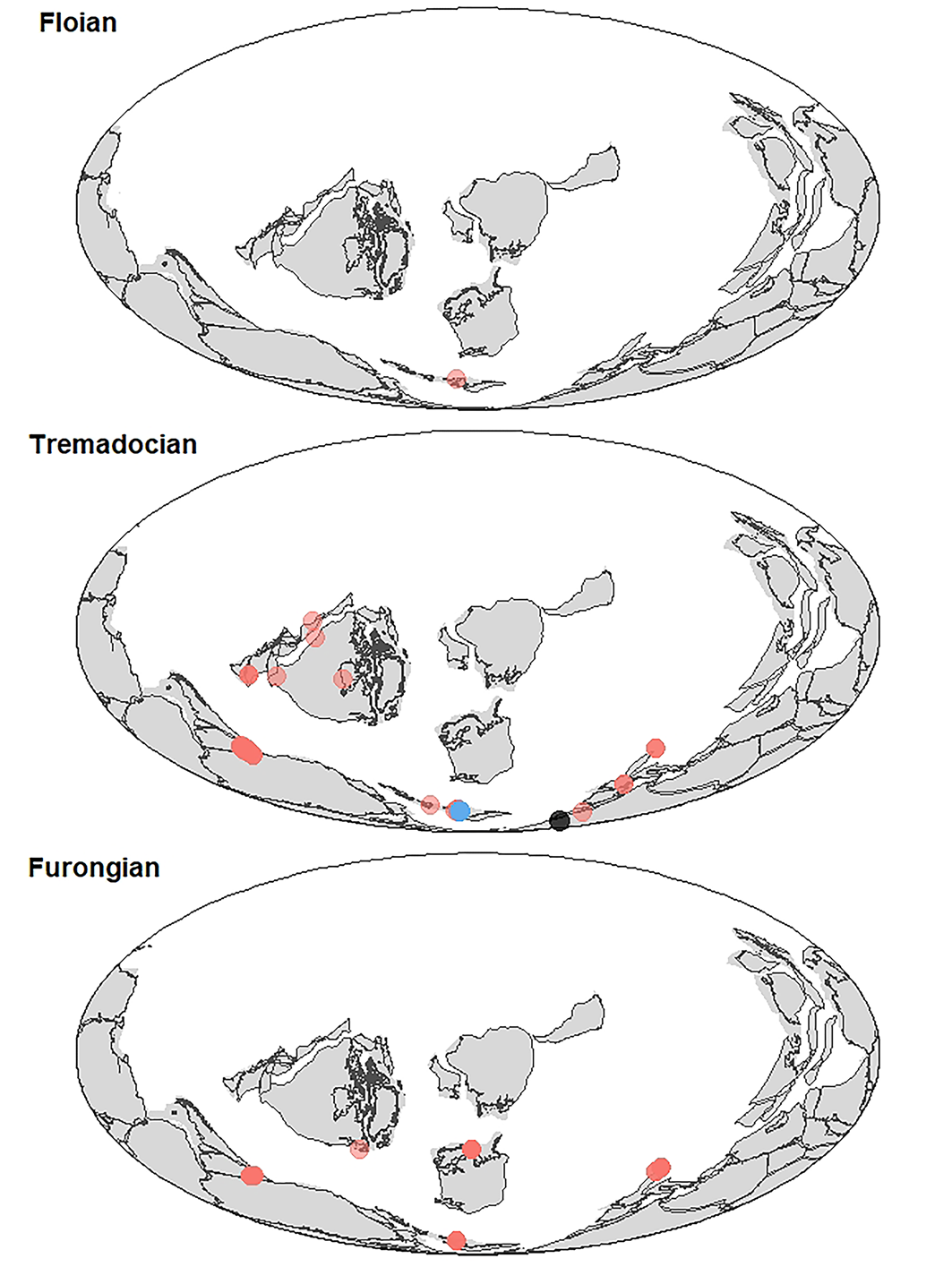
Figure 4. Paleogeographic maps with Leptoplastides occurrences in the Furongian (lower panel), Tremadocian (middle panel), and Floian (upper panel). Increased opacity of locality dots indicates greater number of occurrences in those areas. Blue = Leptoplastides salteri, UK; Black = Leptoplastides salteri, Morocco; Red = all other Leptoplastides species, including those previously assigned to Beltella and Rampartaspis. Paleoreconstructions are based on the PALEOMAP reconstruction model (Scotese and Wright, Reference Scotese and Wright2018) and plotted using a Mollweide projection. Paleogeographic coordinates were downloaded directly from the Paleobiology Database (PBDB) (7 April 2023) using the URL “https://paleobiodb.org/data1.2/occs/list.csv?base_name=Leptoplastides, Rampartaspis&pgm=Scotese&show=coords,paleoloc”. Eighty-six new occurrences and 53 taxonomic names and opinions were entered into the PBDB specifically for this project. Coastlines and polygons for each timeline were reconstructed using GPlates Webservice (gwsdoc.gplates.org) and the R packages rgdal v 1.6-5 (Bivand et al., Reference Bivand, Keitt and Rowlingson2023), ggplot2 v. 3.3.5 (Wickham, Reference Wickham2016), broom v. 1.0.3 (Robinson et al., Reference Robinson, Hayes and Couch2023), ggthemes v. 4.2.4 (Arnold, Reference Arnold2021), and cowplot v. 1.1.1 (Wilke, Reference Wilke2020). See also https://bcmoon.uk/palaeomap/02-gplates-web-service/ for useful guidelines on using the GPlates Webservice.
The genus Leptoplastides has been cited by Hammann et al. (Reference Hammann, Álvaro, Vizcaïno, Rábano and García-Bellido2008, p. 154) from the Lower Ordovician sequence of Celtiberia (northwestern Spain), which also yielded other olenid genera (Angelina and Hypermecaspis). Most probably they come from the lower Tremadocian shales of the Borrachón Formation, from which Hammann et al. (Reference Hammann, Robardet and Romano1982, p. 24) previously reported a trilobite assemblage very closely related to early Tremadocian faunas of South America. Several of the genera cited in the last paper (but with no mention of Leptoplastides) were illustrated for the Borrachón Formation by Álvaro and Martínez-Benítez (Reference Álvaro and Martínez-Benítez2023), who considered the record of the olenid Angelina aff. A. hyeromimi (Kayser, Reference Kayser and Stelzner1876) as evidence of direct correlation of these beds with Oaxaca and the Argentinean margin of Gondwana at the Furongian–Tremadocian boundary interval. The premature death of Wolfgang Hammann prevented publication of a monograph, by then almost finished, describing, among other trilobites, the Iberian material of Leptoplastides and other olenids. The manuscript is presently in the hands of J.J. Álvaro (personal communication, 2022) with the intent to publish posthumously.
Leptoplastides salteri (Callaway, Reference Callaway1877) is one of the youngest species in the genus. In Morocco “Beltella sp.” is known from the Anisograptus matanensis to Rhabdinopora anglica graptolite zones of the early Tremadocian (Lefebvre et al., Reference Lefebvre, Allaire, Guensburg, Hunter, Kouraïss, Martin, Nardin, Noailles, Pittet and Sumrall2016a); in the UK Leptoplastides salteri occurs somewhat above the Adelograptus tenellus graptolite Zone, which places it in the later Tremadocian (Fortey and Owens, Reference Fortey and Owens1991). A single occurrence of Leptoplastides sp. is from either the very latest Tremadocian or the very earliest Floian (Paratetragraptus approximatus graptolite Zone) of Wales (Fortey and Owens, Reference Fortey and Owens1992). The occurrence of L. salteri in the early Tremadocian of Morocco followed by the later Tremadocian occurrence in the UK indicates a possible equatorward immigration of the species during the Early Ordovician, at a time when other closely related clades were dispersing equatorward from other areas of Gondwana (Monti et al., Reference Monti, Confalonieri and Tortello2023). These occurrences are also consistent with previous paleobiogeographic analyses indicating a close association of faunas from western Gondwana and Avalonia (e.g., Álvaro et al., Reference Álvaro, Elicki, Geyer, Rushton and Shergold2003, Reference Álvaro, Ferretti, González-Gómez, Serpagli, Tortello, Vecoli and Vizcaïno2007; Gutiérrez-Marco et al., Reference Gutiérrez-Marco, Sá, García-Bellido and Rábano2017), and similarly, support post-Tremadocian rifting of Avalonia from Gondwana (Murphy et al., Reference Murphy, Gutiérrez-Alonso, Nance, Fernández-Suárez, Keppie, Quesada, Strachan and Dostal2006; Cocks and Fortey, Reference Cocks, Fortey and Basset2009).
Olenid trilobites are frequently abundant and diverse where they occur (e.g., Spitsbergen, Fortey, Reference Fortey1974; Argentina, Balseiro et al., Reference Balseiro, Waisfeld and Vaccari2011; Scandinavia, Terfelt et al., Reference Terfelt, Ahlberg and Eriksson2011), but because they are usually restricted to deeper-water sediments, their fossil record is geographically and temporally discontinuous (see for example the distribution of Cloacaspis species; Hopkins, Reference Hopkins2019). The rarity of olenid trilobites in Morocco is likely due to the relative rarity of deeper-water sediments in an already sparse Furongian and early Tremadocian record (Álvaro et al., Reference Álvaro, Ferretti, González-Gómez, Serpagli, Tortello, Vecoli and Vizcaïno2007; Álvaro and Vizcaïno, Reference Álvaro and Vizcaïno2018). The occurrence of well-preserved Leptoplastides salteri in the N'kob area, however, provides a promising precedent for further fossil discoveries from the earliest Tremadocian of Morocco.
Acknowledgments
Thank you to B. Hunda for inviting us to submit this manuscript to the special issue and to S. Pereira and K. Månsson for thoughtful reviews that improved the quality and relevance of this work. Thank you also to A. Rashkova for minor additional preparation work and to B. Moon for making guidelines for using the GPlates Web Service available online (https://bcmoon.uk/palaeomap/02-gplates-web-service/). This paper is a contribution to projects IGCP 735 (Rocks n'ROL) of the IUGS–UNESCO and PDI2021-125585NB-100 of the Spanish Ministry of Science and Innovation (to JCG-M). This is Paleobiology Database publication 476.
Declaration of competing interest
The authors declare none.
Data availability statement
There are no supplementary data associated with this paper.






 Open access
Open access