


Introduction
The Tengchong and Baoshan blocks, in the western Yunnan of Southwest China, are considered to be Gondwana derived, mainly on the basis of their Permo-Carboniferous Gondwana-affinity sediments (diamictites, pebbly mudstones) and fossils (cool- to cold-water faunas; Wang, Reference Wang1983; Jin, Reference Jin1994, Reference Jin1996, Reference Jin2002; Wopfner, Reference Wopfner1996; Fig. 1). Both blocks have been suggested to have been at the northern margin of Gondwana during the Permo-Carboniferous glaciation and detach thereafter (e.g., Wopfner, Reference Wopfner1996; Shi and Archbold, Reference Shi and Archbold1998; Ueno et al., Reference Ueno, Wang and Wang2003; Wang et al., Reference Wang, Lin, Shen, Chaodumrong, Shi, Wang and Wang2013). The carbonate deposition starting in the late early Permian marks the rapid climate amelioration after the glaciation (Jin et al., Reference Jin, Huang, Shi and Zhan2011). The continuation of the carbonate environment resulted in the formation of Permian (to Triassic) carbonate successions.
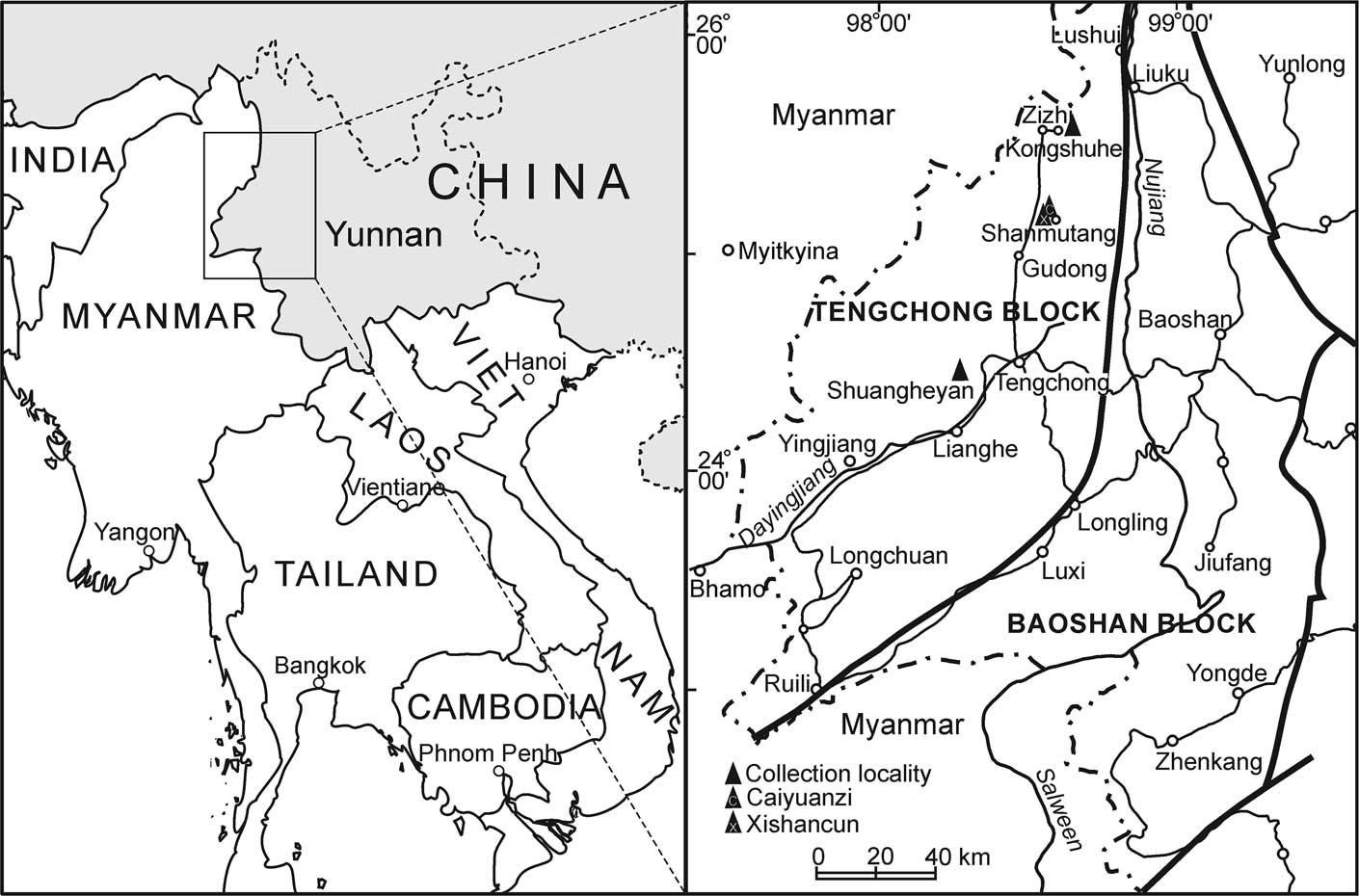
Figure 1 Major tectono-stratigraphic units of western Yunnan and localities of collections.
The understanding of the remarkable climate transition in the Tengchong Block is mainly attributed to the work since the 1980s. Geological mapping and a few studies on Carboniferous and Permian strata in the Tengchong Block have discovered a number of fossils and accumulated important geological data (e.g., Geological Survey of Yunnan, 1979, 1985, 1986; Sheng and He, Reference Sheng and He1983; Fan, Reference Fan1993; Nie et al., Reference Nie, Song, Jiang and Liang1993; Yang, Reference Yang1998; Jin, Reference Jin2002). The brachiopods, bryozoans, crinoids, and small corals reported from the late Carboniferous–early Permian clastic sediments indicated a cool climate, whereas the fusulinids discovered from the Permian carbonates suggested somewhat warmer water (Nie et al., Reference Nie, Song, Jiang and Liang1993; Jin, Reference Jin2002; Duan et al., personal communication, 2005; Shi et al., Reference Shi, Jin, Huang and Yang2008; Jin et al., Reference Jin, Huang, Shi and Zhan2011). Nevertheless, among these fossils, only limited materials have been illustrated and described (e.g., Sheng and He, Reference Sheng and He1983; Fan, Reference Fan1993; Nie et al., Reference Nie, Song, Jiang and Liang1993; Yang, Reference Yang1998; Shi et al., Reference Shi, Jin, Huang and Yang2008); thus most of these reported data are unexaminable for further discussions concerning paleoecology and paleobiogeography. In particular, although fusulinid foraminifera have long been recognized as an environment-sensitive indicator (e.g., Thompson, Reference Thompson1948; Ross, Reference Ross1967, Reference Ross1995; Rui, Reference Rui1981; Leven, Reference Leven1993), the fusulinids in the Tengchong Block were not systematically studied until several years ago (Shi et al., Reference Shi, Jin, Huang and Yang2008).
During recent years, we have conducted several field trips in the Tengchong Block and discovered a considerable number of fusulinid fossils. According to the results of our studies, the early Permian fusulinids are dominated by Eoparafusulina, and the middle Permian fusulinids are dominated by Chusenella and Monodiexodina. These genera have also been found in many other Gondwana-derived blocks although with specific distinctions (Shi et al., Reference Shi, Jin, Huang and Yang2008). In 2010 and 2013, more middle Permian fusulinid specimens were found in both northern and southern regions of the Tengchong Block. This paper describes these newly discovered fossils and discusses their particular taxonomic features. In conjunction with previously reported data, these new materials are then used to unravel the paleogeographic and paleoenvironmental details of the Tengchong Block.
Geologic setting
The Tengchong Block is bounded by the Nujiang fault zone on the east. Its west boundary is hard to define due to the lack of available data, although it has been suggested to lie roughly along the line from Bahmo to Myitkyina in Myanmar (Jin, Reference Jin1996; Fig. 1). The Permo-Carboniferous succession in the Tengchong Block is generally composed of siliciclastics, namely the Menghong Group, in the lower part and carbonates in the upper part. In the northern region, the Menghong Group consists of, in ascending order, the Zizhi Formation and the Kongshuhe Formation and is overlain by carbonates of the Dadongchang Formation (Jin, Reference Jin2002; Jin et al., Reference Jin, Huang, Shi and Zhan2011; Fig. 2). The 600–700 m thick Zizhi Formation is composed of mainly sandstones, whereas the Kongshuhe Formation is composed of diamictites, pebbly mudstones, black mudstones, and siltstones, indicating probably a glacial-deglacial-postglacial sequence (Jin, Reference Jin2002; Jin et al., Reference Jin, Huang, Shi and Zhan2011). The Dadongchang Formation consists of up to 600 m thick limestones and dolomitic limestones. The early and middle Permian fusulinids reported previously were collected from the lower part of the limestones (Jin, Reference Jin2002; Shi et al., Reference Shi, Jin, Huang and Yang2008). The fusulinid materials that we newly collected from the northern Tengchong Block are also from the lower part of the Dadongchang Formation (Fig. 2). Samples are from a section about 500 m south of the Xishancun Village of the Shanmutang area (hereafter the Xishancun fauna). The locality is about 2 km south to the Caiyuanzi section where the middle Permian Chusenella-Monodiexodina fauna was reported by Shi et al. (Reference Shi, Jin, Huang and Yang2008) in the same area (hereafter the Caiyuanzi fauna; Fig. 1).
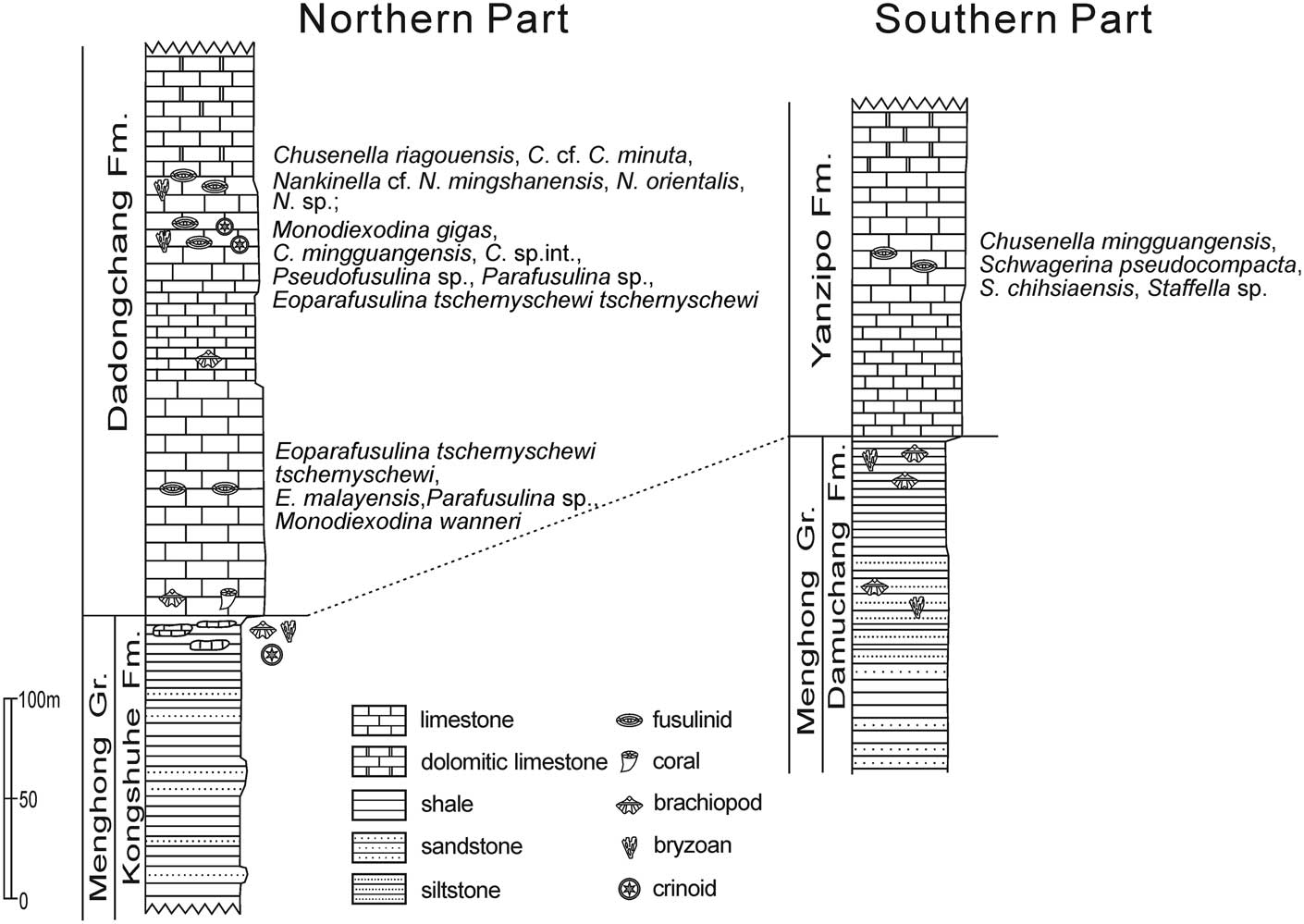
Figure 2 The composite Permian stratigraphic successions in the northern and southern Tengchong Block showing occurrence of fusulinids studied here and reported by Shi et al. (Reference Shi, Jin, Huang and Yang2008). The succession in the northern part is based on sections near the Shanmutang village, and in the southern part is based on sections in the Damuchang and Shuangheyan areas (Jin, Reference Jin1994).
The Menghong Group in the southern Tengchong Block includes, in ascending order, the Bangdu Formation, Luogengdi Formation, Siguaping Formation, and Damuchang Formation. It has a thickness of about 1,500 m and exhibits a similar lithological sequence to that in the northern region, except for lateral variation of thicknesses of main lithological components. Above the siliciclastics rest the carbonates of the Yanzipo Formation, composed of massive fossiliferous limestones in the lower part, where fusulinids have been reported (Geological Survey of Yunnan, 1986; Fan, Reference Fan1993), and dolomitic limestones and dolomites in the upper part, which was once named ‘Shuangheyan Formation’ (Geological Survey of Yunnan, 1986; Jin, Reference Jin1994; Fig. 2). The fusulinid samples we newly collected in the southern Tengchong Block are from loose stones on the carbonate hillside of the Shuangheyan valley (hereafter the Shuangheyan fauna; Fig. 1).
Materials and methods
About 20 kg of limestones containing fusulinids were collected from the Xishancun Section of Shanmutang area and the Shuangheyan valley. The axial thin sections of fusulinids were prepared by grinding the specimens parallel to the coiling axial until the middle of the proloculus was exposed. Altogether, 186 axial thin sections have been prepared and then carefully examined for the identification. Digital images of each specimen were collected using cameras attached to a transmitted light microscope with digital resolutions of 72 dots per inch (dpi).
Repositories and institutional abbreviations
All illustrated specimens are housed in the School of Earth Sciences and Engineering, Nanjing University, China. Specimen catalog numbers indicate either collecting localities or dates.
Systematic paleontology
Classification follows the scheme of Sheng et al. (Reference Sheng, Zhang and Wang1988).
Superfamily Fusulinoidea Möller, Reference Möller1878
Family Schwagerinidae Dunbar and Henbest, Reference Dunbar and Henbest1930
Subfamily Pseudoschwagerininae Zhang, Reference Zhang1963
Genus Chusenella Hsu, Reference Hsu1942
Type species
Chusenella ishaensis Hsu, Reference Hsu1942 from the Chungkuh Limestone of Yishan area in Kuangxi province, China, by original designation.
Chusenella riagouensis Chen, 1985 in Zhang, Chen, and Yu, Reference Zhang, Chen and Yu1985
Figure 3.17, 3.18, 3.20, 3.21, 3.23–3.26

Figure 3 Fusulinids from the Xishancun section of Shanmutang area. (1–4, 10) Nankinella orientalis, cat. nos. 37-6-1, 37t-1-4, 130317-4-18, 37t-1-5-2, 37-6-2; (5, 6, 11) Nankinella mingshanensis, cat. nos. 37-12-2, 37-9, 37t-1-11; (7–9) Nankinella sp., cat. nos. 130317-4-10, 130317-4-22, 130317-4-11; (12–16, 19, 22, 27) Chusenella cf. C. minuta, cat. nos. 130317-4-7, 37-7, 37t-1-3, 130317-4-15, 37t-2-6, 37t-1-5-1, 37t-2-1, 37-19; (17, 18, 20, 21, 23–26) Chusenella riagouensis, cat. nos. 37-21, 130317-4-3, 37-4, 130317-4-14, 37t-2-7, 130317-4-23, 37-5-2, 37-11. Scale bar=1 mm.
1985Chusenella riagouensis Chen; Reference Zhang, Chen and YuZhang et al., p. 131, pl. 2, fig. 7.
1982Chusenella sinensis minor Chu; Reference ZhuZhu, p. 118, pl. 2, figs. 19, 20.
2010Chusenella sinensis Sheng, Reference Sheng1963; Reference Zhang, Cheng and ShenZhang et al., p. 971, fig. 5.31, 5.32, 5.39–5.48.
Holotype
The specimen (Sz2f2) from the Zhuobu Formation, Xainza area of central Tibet, China (Zhang et al., Reference Zhang, Chen and Yu1985, pl. 2, fig. 7).
Occurrence
The present species only occurred in the Tengchong Block and central Tibet (Zhang et al., Reference Zhang, Chen and Yu1985; Zhang et al., Reference Zhang, Cheng and Shen2010).
Description
Test small, fusiform with bluntly pointed poles. Adult tests commonly contain 5–7.5 whorls, with length in the range of 2.8–3.2 mm and width 1.2–1.5 mm. The first three whorls tightly coiled with sharply pointed poles, and after that the coiling becomes looser. Spirotheca composed of the tectum and coarsely alveolar keriotheca. Septa are nearly straight in the inner whorls and flute strongly but regularly. Septal folds are nearly as high as half of the chamber. Proloculus is moderate in size, commonly 0.1–0.2 mm in outside diameter. Secondary deposits fill the axial regions.
Remarks
The present species is typical in the Xainza area of central Tibet. Zhu (Reference Zhu1982) assigned the specimens from Xainza to Chusenella sinensis minor because they share similarity with Chusenella sinensis Sheng and also have significantly smaller size. Chen (in Zhang et al., Reference Zhang, Chen and Yu1985) renamed the specimen holding same characters in the same area as Chusenella riagouensis and published the holotype. Zhang et al. (Reference Zhang, Cheng and Shen2010) assigned those from Xainza to Chusenella sinensis but didn’t explain the differences of size. The specimens from Tengchong Block are rather similar, with regard to adult test shape and ontogenetic shape changes, to the types of Chusenella sinensis, but they are only half the size. Therefore, it is not appropriate to assign them into C. sinensis until there is enough evidence to demonstrate that the species could cover a quite wider size distribution. Therefore, we assign the present group of specimens to Chusenella riagouensis.
Chusenella cf. C. minuta Skinner, Reference Skinner1969
Occurrence
This species only discovered in the Tengchong Block hitherto.
Description
Test small, inflated fusiform with distinctively bluntly pointed poles. Adult tests have 6.5–8 whorls with 3.4–4.5 mm in length and 1.5–2.3 mm in width, giving axial ratio 1.8–2.6. The first three or four whorls tightly coiled with sharply pointed poles, and after that the coiling becomes looser. Spirotheca composed of the tectum and coarsely alveolar keriotheca. Septa are nearly straight in the inner whorls and flute strongly but regularly. Septal folds are slightly higher than half of the chamber’s height. Proloculus is moderate in size, 0.1–0.2 mm in outside diameter. Secondary deposits fill the axial regions.
Remarks
The present specimens are similar to Chusenella minuta Skinner, Reference Skinner1969 in terms of the adult test shape and general ontogenetic shape change, but differ in the less tightly coiled inner whorls and slightly bigger adult test.
Chusenella mingguangensis Shi et al., Reference Shi, Jin, Huang and Yang2008
Figure 4.1–4.7, 4.10, 4.11, 4.13–4.16

Figure 4 Fusulinids from the Shuangheyan area. (1–7, 10, 11, 13–16) Chusenella mingguangensis, cat. nos. 090303-2-1, 090303-2-2, shy100321-2-7, 090304-2-11, 090303-2-11, shy100320-1-18, 090303-2-6, 090303-2-10, shy100319-6-10, 090304-2-4, shy100321-2-23, shy100320-1-1, 090304-2-9; (8, 9) Staffella sp., cat. nos. shy100321-1-2, shy100321-1-10; (17–23) Schwagerina chihsiaensis, cat. nos. shy100320-1-26, shy100320-1-32, 090304-2-12, shy100320-2-8, shy100320-1-11, shy100320-1-2, shy100320-1-23; (12, 24–26) Schwagerina pseudocompacta, cat. nos. 090303-2-4, shy100320-1-9, shy100321-2-3, shy100320-2-9. (1–7, 10–26) Scale bar=1 mm; (8, 9) scale bar=500 µm.
2008Chusenella mingguangensis Reference Shi, Jin, Huang and YangShi et al., p. 126, fig. 4.1–4.8, 4.10–4.16, 4.20.
Holotype
The specimen (MCC2-b7) from the Dadongchang Formation, Shanmutang area of northern Tengchong, China (Shi et al., Reference Shi, Jin, Huang and Yang2008, p. 126, fig. 4.2)
Occurrence
This species only discovered in the Tengchong Block hitherto.
Description
Test medium, elongated fusiform with bluntly rounded poles. Adult tests have 5.5–7 whorls with 4.5–6 mm in length and 1.2–1.8 mm in width, resulting axial ratio 3.5–4. The first two or three whorls tightly coiled, others expanding gradually. Spirotheca composed of the tectum and keriotheca. Septa folds regularly and widely spaced with height more than half the chambers. Proloculus is moderate in size, 0.1–0.2 mm in outside diameter. Secondary deposits fill the polar regions.
Remarks
This species is rather abundant in the Caiyuanzi area, its type locality. Specimens characteristically exhibit intensively involute whorls and extensively developed axial fillings. Although specimens of Shuangheyan are poorly preserved compared with the contemporaneous ones from the Shanmutang area, identical aforementioned characters could still be recognized.
Subfamily Schwagerininae Dunbar and Henbest, Reference Dunbar and Henbest1930
Genus Schwagerina Möller, Reference Möller1877
Type species
Borelis princeps Ehrenberg, Reference Ehrenberg1842 from Am Onega-See, Russland, figured by Ehrenberg (Reference Ehrenberg1854), reassigned as genotype of Schwagerina by Möller (Reference Möller1877), and restudied by Dunbar and Skinner (Reference Dunbar and Skinner1936).
Schwagerina chihsiaensis (Lee, Reference Lee1931)
1931Schellwienia chihsiaensis Reference LeeLee, p. 287, pl. 1, fig. 2, 2a.
1962Schwagerina chihsiaensis (Lee), Reference ShengSheng, p. 102, pl.15, fig. 5.
1986Schwagerina chihsiaensis Sheng, Reference Sheng1962; Reference Xiao, Wang and ZhangXiao et al., p. 104, pl. 5, figs. 10, 14–17.
Holotype
The specimen (without catalog number) from the uppermost Chihsia Limestone, Nanjing area, China (Lee, Reference Lee1931, pl. 1, fig. 2)
Occurrence
This species was first discovered in the Kungurian Chihsia limestone of South China (Sheng, Reference Sheng1962) and could range up to the Guadalupian Maokou Formation (Zhang et al., Reference Zhang1988). It was widely distributed in South and North China (Fan et al., Reference Fan, Chen, Hou, Miller, Melchin, Shen, Wu, Goldman, Mitchell and Yang2013).
Description
Test median, fusiform with bluntly rounded poles. The inner whorls with more sharply pointed poles. Adult tests grow 6.5–7 whorls, with 7.4–8.4 mm in length and 1.8–2.3 mm in width, giving form ratio 3.0–4.2. Spirotheca composed of the tectum and coarsely alveolar keriotheca. Septa flute strongly but regularly. Septal folds are lower than half of the chamber’s height in the inner whorls but becoming higher in the outer whorls. Proloculus is moderate in size, around 0.2 mm in outside diameter. Axial fillings present.
Schwagerina pseudocompacta Sheng, Reference Sheng1956
1956Schwagerina pseudocompacta Reference ShengSheng, p. 190, pl. 8, figs. 4–6.
1962Schwagerina pseudocompacta; Reference ShengSheng, p. 106, pl. 17, fig.8.
1986Schwagerina pseudocompacta; Reference Xiao, Wang and ZhangXiao et al., p. 103, pl. 10, fig. 13.
2010Schwagerina pseudocompacta; Reference Zhang, Cheng and ShenZhang et al., p. 967, fig. 5.21–5.30.
Holotype
The specimen (No. 8165) from the Maokou Formation, Liangshan area of Shaan’xi, China (Sheng, Reference Sheng1956, pl. 8, fig. 6).
Occurrence
This species is widely distributed in the Guadalupian Maokou Formation in North and South China, and in the Lhasa Block (Fan et al., Reference Fan, Chen, Hou, Miller, Melchin, Shen, Wu, Goldman, Mitchell and Yang2013).
Description
Test small, fusiform with bluntly rounded poles. Spirotheca composed of the tectum and coarsely alveolar keriotheca. Adult tests commonly have 5–6.5 whorls, with 4.0–5.4 mm in length and 1.7–2.3 mm in width, giving axial ratio 2.3–2.7. Septa are fluted strongly and irregularly. Septal folds are nearly as high as half of the chamber’s height. Proloculus is moderate in size, around 0.2 mm in outside diameter. Axial fillings present.
Family Staffellidae Miklukho-Maklay, Reference Miklukho-Maklay1949
Subfamily Staffellinae Miklukho-Maklay, Reference Miklukho-Maklay1949
Genus Nankinella Lee, Reference Lee1934
Type species
Staffella discoides Lee, Reference Lee1931 from the upper Chihsia Limestone of Nanjing in Jiangsu province, China, assigned as the genotype of Nankinella by Lee (Reference Lee1934).
Nankinella mingshanensis Sheng and Rui, Reference Sheng and Rui1984
1984Nankinella mingshanensis Reference Sheng and RuiSheng and Rui, p. 34, pl. 1, figs. 24–26.
Holotype
The specimen (no. 82384) from the Palaeofusulina sinensis zone, Leping village of Jiangxi, China (Sheng and Rui, Reference Sheng and Rui1984, pl. 1, fig. 25).
Occurrence
This species was first discovered in the upper Permian strata in Jiangxi (Sheng and Rui, Reference Sheng and Rui1984) and later discovered in the Maokou Formation in Guizhou (Zhang et al., Reference Zhang1988) and Yunnan (Zhou, Reference Zhou1998).
Description
Test median, subrhomboidal with narrowly pointed periphery, slightly convex lateral sides, and closed umbilici. Adult tests commonly develop 6–8 whorls, with length in the range of 1.2–1.7 mm, width 1.9–2.9 mm, giving axial ratio around 0.6. Spirotheca with three layers, tectum as well as outer and inner tectorium. Septa straight. Proloculi are quite small, less than 0.1 mm in diameter, therefore difficult to recognize and measure in most thin sections.
Nankinella orientalis Miklukho-Maklay, Reference Miklukho-Maklay1954
1954Nankinella orientalis Reference Miklukho-MaklayMiklukho-Maklay, p. 70, pl. 11, figs. 1–4.
1963Nankinella orientalis; p. 31, pl. 2, figs. 8–10.
Holotype
The specimen (no. 226-4-45) from a pebble of foraminiferal limestone from Lower Triassic conglomerates in Epchika area, northern Caucasus (Miklukho-Maklay, Reference Miklukho-Maklay1954).
Occurrence
This species is commonly distributed in the Changhsingian of North and South China, and has been reported from the Guadalupian strata in Azerbaijan associated with Chusenella and Verbeekina (Ruzhentsev and Sarycheva, Reference Ruzhentsev and Sarycheva1965).
Description
Test median, subrhomboidal with rounded periphery, slightly convex lateral sides, and closed umbilici. Test of inner whorls with narrowly pointed periphery. Adult tests commonly develop 6–9 whorls, with 1.5–1.9 mm in length and 2.0–2.5 mm in width, giving axial ratio around 0.7. Spirotheca with three layers, tectum as well as outer and inner tectorium. Septa straight. Proloculi are less than 0.1 mm in diameter.
Nankinella sp.
Description
Test median, elliptical with rounded periphery, slightly convex lateral sides and closed umbilici. Tests of the inner whorls are subrhomboidal with narrowly pointed periphery. Adult tests develop 6–7.5 whorls, with length of 1.3–1.4 mm, width of 1.5–1.9 mm, giving axial ratio around 0.7. Spirotheca are composed of tectum, upper and lower tectorium. Septa straight. Proloculi are less than 0.1 mm in diameter.
Remarks
The present species is similar to N. orientalis except for the much smaller size and the elliptical shape of the adult tests.
Results
The Xishancun fauna from the northern Tengchong Block is composed of Chusenella riagouensis Chen, 1985, C. cf. minuta Skinner, Nankinella cf. mingshanensis Sheng and Rui, N. orientalis Miklukho-Maclay, 1954, and Nankinella sp. (Fig. 3). Among them, Chusenella riagouensis is the dominant species. Until now it was only found in the Xainza area of Tibet (Zhang et al., Reference Zhang, Chen and Yu1985). It is distinct from other Chusenella species in its small size. This species occurs in the Zhaopu (Zhuobu) Formation of the Xainza area and indicates an age of Roadian to Capitanian. Nankinella mingshanensis Sheng was first found in the upper Permian in Jiangxi (Sheng and Rui, Reference Sheng and Rui1984) and later discovered in the Guadalupian Maokou Formation in Guizhou and Yunnan of South China (Zhang et al., Reference Zhang1988; Zhou, Reference Zhou1998). Nankinella orientalis is widely distributed in the Changhsingian strata of North and South China (Geobiodiversity Database, Fan et al., Reference Fan, Chen, Hou, Miller, Melchin, Shen, Wu, Goldman, Mitchell and Yang2013) and ranges down to the Guadalupian strata in Azerbaijan (Ruzhentsev and Sarycheva, Reference Ruzhentsev and Sarycheva1965). These data suggest an age of Roadian–Capitanian for the Chusenella-Nankinella fauna.
The Shuangheyan fauna from the southern Tengchong Block consists of four species: Chusenella mingguangensis Shi et al., Reference Shi, Jin, Huang and Yang2008, Schwagerina chihsiaensis (Lee, Reference Lee1931), Schwagerina pseudocompacta Sheng, Reference Sheng1956, and Staffella sp. (Fig. 4). Chusenella mingguangensis is the dominant element here, and is also prolific in the previously reported Wordian–Capitanian Caiyuanzi fauna (Shi et al., Reference Shi, Jin, Huang and Yang2008). This species is rather unique for its small size and intensely coiled inner whorls. So far it has only been discovered in the Tengchong Block. Schwagerina chihsiaensis was first discovered in the Kungurian Chihsia limestone in South China (Lee, Reference Lee1931). It ranges up to the Guadalupian Maokou Formation (Zhang et al., Reference Zhang1988). Schwagerina pseudocompacta occurs widely in the Maokou Formation in South China (Fan et al., Reference Fan, Chen, Hou, Miller, Melchin, Shen, Wu, Goldman, Mitchell and Yang2013). These elements altogether suggest a Wordian–Capitanian age.
In 2008, we documented two other fusulinid faunas from the northern Tengchong Block, the early Permian Eoparafusulina fauna from the Kongshuhe area and the middle Permian Caiyuanzi fauna from the Shanmutang area (Shi et al., Reference Shi, Jin, Huang and Yang2008). The Eoparafusulina fauna includes four species belonging to Eoparafusulina, Parafusulina, and Monodiexodina, and the Chusenella-Monodiexodina (Caiyuanzi) fauna is composed of six species in five genera, including Pseudofusulina, Eoparafusulina, and Parafusulina besides the eponymous genera. The Caiyuanzi fauna is dominated by Chusenella mingguangensis and Monodiexodina gigas Shi et al., Reference Shi, Jin, Huang and Yang2008.
The common dominant element in the three middle Permian faunas of the Tengchong Block is Chusenella, and the species discovered, i.e., C. riagouensis, C. mingguangensis, C. cf. minuta, are all small or medium in size. Chusenella is widely distributed in southern Europe, North America, Japan, South China, and Central and Southeast Asia, but more than two-thirds of the species are medium to large in size, which means over 6 mm in test length. Most Chusenella species from the Tengchong Block are less than 5 mm in length, and even the largest species, Chusenella mingguangensis, is only around 6 mm long. The small Chusenella species are also significant in the Lhasa Block, and those reported from the Xainza area are hardly over 4 mm in length (Huang et al., Reference Huang, Jin, Shi and Yang2007; Zhang et al., Reference Zhang, Cheng and Shen2010; Zhang et al., Reference Zhang, Shi and Shen2013).
Discussion
Depauperate faunas and paleoenvironmental inference
The common feature for all four faunas of the Tengchong Block is low generic and specific diversity with abundant specimens. Although fusulinid individuals discovered in all four locations were plentiful, no more than five genera and six species have been recovered in any given fauna, and nearly half the individuals of each fauna belong to the predominant species.
Modern examples of depauperate foraminiferal assemblages are diagnostic of inhospitable environments. For example, Amphistegina lobifera Larsen, Reference Larsen1976 is a larger benthic foraminiferal species with a tolerance for low winter temperatures and mixotrophic feeding strategy. It is found in the Mediterranean and dominates the benthic foraminiferal assemblages there (Triantaphyllou et al., Reference Triantaphyllou, Dimiza, Koukousioura and Hallock2012). Androsina lucasi Lévy, Reference Lévy1977 thrives in open, dwarf-mangrove flats areas in exceptional abundance, owing to its euryhaline adaptation (Hallock and Peebles, Reference Hallock and Peebles1993). The Frierfjord of Norway was heavily polluted by waste products for centuries and contains anoxic deep bottom water. An opportunistic species, Stainforthia fusiformis (Williamson, Reference Williamson1858) thrives at > 50 m water depth in this area with a large number of individuals (Alve, Reference Alve1994). These species have developed the ability to survive and flourish in stressful environments. Similarly, the depauperate fusulinids of the Tengchong Block very likely suggest a somewhat inhospitable shallow sea for fusulinids during the early and middle Permian.
Most fusulinids favored open shallow marine environments with warm and clear water (Ross, Reference Ross1995), probably or partly due to the likelihood that photosymbionts within them required sunlight (Vachard et al., Reference Vachard, Munnecke and Servais2004; Groves et al., Reference Groves, Pike and Westley2012). Accordingly, the inhospitable environmental factors for fusulinids likely include low temperature and specific carbonate facies related with hydrodynamics, depth, and sunlight.
Since there is only one early Permian fusulinid fauna from the Tengchong Block, it is difficult to discern the relationship between carbonate facies and fusulinids during that time. By contrast, middle Permian carbonates containing depauperate faunas include a variety of sedimentary facies (Fig. 5). One is skeletal grainstone in the Shuangheyan and Caiyuanzi areas, indicating high-energy open shallow marine environments (Fig. 5.5, 5.6). Within this facies, carbonate grains are usually moderately sorted and rounded. Skeletal grains are diverse with fusulinids, bryozoans, echinoderms, smaller foraminifera, green algae, etc., and fusulinid tests are more or less abraded. In the Shuangheyan area, the grainstone shows two distinct generations of spar cementation. It is also noteworthy that Monodiexodina in the Caiyuanzi area is particularly confined to well-sorted and rounded grainstone with mostly fine-grained bioclasts. This special facies type suggests a littoral marine setting with high water energy. The second facies is poorly sorted and rounded skeletal packstone with fusulinids, smaller foraminifera, bryozoans, and intraclasts in the Shuangheyan and Caiyuanzi areas (Fig. 5.1, 5.2). The matrix was originally micritic, but it has undergone neomorphism to become pseudospar. These characteristics indicate open shallow platform with moderate water agitation. It is interesting to note that fusulinids are broken and infilled with black micrite in the packstone of the Shuangheyan area. This probably indicates that fusulinids experienced a certain degree of transportation or turbulence. Another facies type is partially dolomitized wackestone in the Shuangheyan and Xishancun areas (Fig. 5.3, 5.4). Chusenella and Nankinella/Staffella are characteristically dominant, and algae are abundant in this facies, whereas other fusulinid taxa and other fossil groups are rather impoverished. In the Xishancun section, the micritic matrix of this facies shows a clotted structure. A more restricted platform with stagnant water could well generate such wackestones.

Figure 5 Diverse lithological facies in fusulinid-containing sections/areas of Tengchong. (1) Packstone, from Caiyuanzi Section of Shanmutang area, cat. no. NMC+3-2; (2) packstone, Shuangheyan area, cat. no. 100320-2-1; (3) dolomitized wackestone with (3a) exhibiting dolomites, from Shuangheyan area, cat. no. 100321-1-3; (4) wackestone, from Xishancun section of Shanmutang area, 130317-4-7; (5) grainstone, from Shuangheyan, cat. no. 090304-2-1; (6) grainstone with fine-grained bioclasts and Monodiexodina gigas, from Caiyuanzi Section, cat. no. MCC8-11. (1–3, 4–6) Scale bars=1 mm; (3a) scale bar=200 µm.
These varied carbonate facies reflect various hydrodynamic conditions inhabited by the depauperate faunas and suggest water turbulence is not responsible for the low diversities. The development of green algae in these faunas denotes adequate sunlight in these shallow marine environments. As a result, the probable controlling factor of these impoverished faunas could be unfavorable temperature. Therefore, we infer that Permian depauperate fusulinids in the Tengchong Block probably inhabited seas with relatively cool temperate temperatures.
The Gondwana-affinity feature and paleogeographic discussion
We compiled all the fusulinid data hitherto documented from the Tengchong Block in order to fully understand the fusulinid composition and their paleogeographic indications. There are several reports of Permian fusulinids from the Tengchong Block prior to 2008, although without illustrations or descriptions (Geological Survey of Yunnan, 1985, 1986; Fan, Reference Fan1993; Nie et al., Reference Nie, Song, Jiang and Liang1993). According to our recent study, specimens previously reported as Hemifusulina from the Kongshuhe area (Fan, Reference Fan1993; Nie et al., Reference Nie, Song, Jiang and Liang1993) should be assigned to Eoparafusulina (Shi et al., Reference Shi, Jin, Huang and Yang2008). All other reported fusulinid genera from the block are valid and considered in the following discussion.
The generic diversities of fusulinids through the early and middle Permian of the major Gondwana-derived blocks, including the Tengchong Block, are listed in Table 1. Integrated generic lists from peninsular Thailand and northwest peninsular Malaysia (PTNPM), Shan State of Burma, Lhasa, Baoshan, and Tengchong blocks are presented because fusulinid data from these blocks have been updated recently.
Table 1 Permian fusulinid generic diversity in the major Gondwana-derived blocks.

Numbers in square brackets indicate genera number of Pseudoschwagerininae; starred genera and numbers within parentheses refer to those of Verbeekinidae and Neoschwagerinidae.
The early Permian fusulinids of the Cathaysian (tropical Tethyan) region are characterized by many species in the subfamily Pseudoschwagerininae. By contrast, the Gondwana-derived blocks contain generally less diverse pseudoschwagerine. For example, the common genera Pseudoschwagerina, Robustoschwagerina, Sphaeroschwagerina, and Zellia are abundant in most sections of South China (Zhu et al., Reference Zhu, Yang and Yin2002; Shi et al., Reference Shi, Yang and Huang2009), but they are sparsely distributed in the Gondwana-derived blocks (Ueno, Reference Ueno2003; Table 1). Moreover, the overall diversities of early Permian fusulinids in the Gondwana-derived blocks are commonly lower than those in South China. For example, the Asselian–Sakmarian fusulinids typically include over 15 genera in South China, whereas most Gondwana-derived blocks have fewer than 10 genera. In the Tengchong Block, early Permian fusulinids are only composed of three genera, Eoparafusulina, Pseudofusulina, and Monodiexodina, and the association is devoid of Pseudoschwagerininae.
The middle Permian fusulinids of the Gondwana-derived blocks are characterized by: (1) the relatively poor development of the tropical/subtropical Verbeekinidae and Neoschwagerinidae taxa, and (2) the association of certain Schwagerinidae genera, such as Chusenella, Rogososchwagerina, and Eopolydiexodina (Ozawa, Reference Ozawa1987). In the Tengchong Block, only one genus of Neoschwagerinidae, i.e., Cancellina, has been reported from the middle Permian strata of the northern region. Instead, Chusenella is the principal element. It occurs abundantly and is accompanied by Nankinella or Monodiexodina in the Shanmutang area and by Schwagerina in the Shuangheyan valley. In this respect, the fusulinids from the Tengchong Block bear strong Gondwana affinity.
With regard to the generic diversity through the early and middle Permian, the Gondwana-derived blocks could be further divided into two groups. Group 1 contains Turkey, Iran, and Transcaucasia, south Afghanistan, south Pamir; Group 2 includes the Lhasa Block, PTNPM, Shan State, Baoshan, and the Tengchong Block. Fusulinids from the former have evidently higher diversities than those from the latter in general, especially during Kungurian and the succeeding middle Permian.
Specifically, in the Asselian to Artinskian (early Permian) times, the blocks of Group 2, except the Tengchong Block, hosted fewer fusulinid genera than those of Group 1. The generic number from Tengchong Block might have been overestimated because some of the previously reported genera cannot be checked due to the lack of illustration or description. In the Kungurian, fusulinids in Group 1 areas were diverse, characterized by the proliferation of newly developed Verbeekinidae and Neoschwagerinidae genera containing a new inner skeletal structure named parachomata. However in Group 2 areas, fusulinids scarcely occurred, and the only recorded genera, i.e., Monodiexodina and Parafusulina, developed another new structure called cuniculi.
During the middle Permian, fusulinid in Group 1 areas were more diverse, including certain newly evolved taxa, i.e., Neoschwagerinidae and Verbeekinidae. Group 2 areas still hosted less diversified fusulinids, especially within the Neoschwagerindae and Verbeekinidae, than Group 1. The diversification of the families Verbeekinidae and Neoschwagerinidae is believed to be diagnostic for the middle Permian Tethyan region (Leven, Reference Leven1993; Ross, Reference Ross1995). Representatives of these families are thought to have been thermally stenotopic and typify tropical or subtropical warm-water environment (Gobbett, Reference Gobbett1967; Ozawa, Reference Ozawa1970; Ross, Reference Ross1982; Stevens, Reference Stevens1985; Ueno, Reference Ueno2003). They are abundant in the Cathaysian Tethyan region and well developed in Group 1 areas, but obviously less so in Group 2. In most blocks of Group 2, Schwagerinidae fusulinids, such as Chusenella, play the major role. In the Tengchong Block, Chusenella co-occurs with either Schwagerina or Nankinella, and in the PTNPM it is accompanied by Schwagerina, Nankinella, and Staffella in several sites (Ingavat-Helmcke, Reference Ingavat-Helmcke1993; Hassan et al., Reference Hassan, Aung, Becker, Rahman, Ng, Ghani and Shuib2014). A Chusenella-Nankinella association was reported from the Xainza area of the Lhasa Block, although the dominant species are different from those of the Tengchong Block (Huang et al., Reference Huang, Jin, Shi and Yang2007; Zhang et al., Reference Zhang, Cheng and Shen2010). It is noteworthy that the Chusenella species in the Xainza area are rather small, displaying a certain similarity to those of the Tengchong Block as mentioned before. In the Baoshan Block, Chusenella was found in various locations with Schwagerina, Rugosofusulina, or Nankinella (Huang et al., Reference Huang, Shi and Jin2015b). The genus has not been reported yet from the Shan State.
In summary, by comparison with Group 1, the regions of Group 2 demonstrate overall fewer fusulinid taxa in both early and middle Permian and impoverished Neoschwagerinidae and Verbeekinidae. The Gondwana-derived blocks were geographically reconstructed to lie in a southern transitional region between the Paleo-equator and temperate/cool Gondwanan Pangea during most of the Permian (Shi et al., Reference Shi, Archbold and Zhan1995; Grunt and Shi, Reference Grunt and Shi1997; Shi and Archbold, Reference Shi and Archbold1998; Ueno, Reference Ueno2003). Most Group 2 members, i.e., PTNPM, Shan State, Baoshan Block, are interpreted to occur in the higher latitude portion of the southern transition region (Ueno, Reference Ueno2003; Huang et al., Reference Huang, Jin and Shi2015a, Reference Huang, Shi and Jin2015b, personal communication, 2016). In this regard, the Tengchong Block shares more similarities with these temperate marine blocks. This is consistent with the aforementioned paleoenvironment inference.
There are still several notable distinctions between the fusulinids of Tengchong Block and those of other Gondwana-derived blocks, revealing a somewhat endemic nature of the block. Whereas there are several Neoschwagerinidae and Verbeekinidae genera developed in the Shan State, Baoshan Block, and Lhasa Block of Group 2, they are rather scarce in the PTNPM and Tengchong Block. In the Tengchong Block, only the genus Cancellina of Neoschwagerinidae was reported in the Guanyinshan area (Fan, Reference Fan1993). We searched that area during the fieldwork in 2010, but failed to verify its presence.
Monodiexodina was suggested to be an antitropical genus and confined paleogeographically to the northern and southern middle latitudinal areas (Ueno, Reference Ueno2006). However, there is no Monodiexodina reported from the Lhasa Block, Shan State, or Baoshan Block, but Monodiexodina gigas is a dominant species of the middle Permian of the Tengchong Block. Moreover, this species is significantly distinct from the Monodiexodina species of PTNPM (Fontaine et al., Reference Fontaine, Chonglakmani, Bin and Piyasin1994; Hassan et al., Reference Hassan, Aung, Becker, Rahman, Ng, Ghani and Shuib2014), and any other reported Monodiexodina species, for evolving a huge test with the diameter of 2 cm (Shi et al., Reference Shi, Jin, Huang and Yang2008).
Among the fusulinids absent from the Tengchong Block, Eopolydiexodina is thought to have been restricted to the Laurentian and peri-Gondwanan borders of the Tethys (Ueno, Reference Ueno2003; Colpaert et al., Reference Colpaert, Monnet and Vachard2014). It is an endemic genus in the Western Tethys Province and constitutes a typical combo associated with Rugososchwagerina or Xiaoxinzhaiella (Ueno, Reference Ueno2003; Shi et al., Reference Shi, Yang and Jin2005). This combo is absent in the Tengchong Block, but thrived in both Baoshan and Lhasa blocks.
Conclusions
-
1. This paper describes two newly collected fusulinid faunas from the Tengchong Block, involving Chusenella riagouensis, C. cf. minuta, Nankinella cf. mingshanensis, N. orientalis, and Nankinella sp. from the Shanmutang area, and Chusenella mingguangensis, Schwagerina chihsiaensis, Schwagerina pseudocompacta, and Staffella sp. from the Shuangheyan area. The former indicates an age of Roadian–Capitanian (middle Permian), and the latter suggests a Wordian–Capitanian (middle Permian) age.
-
2. Middle Permian fusulinid faunas discovered in the Tengchong Block display the depauperate characteristic of low generic and specific diversities with abundant specimens. This may suggest an inhospitable temperate zone of shallow sea.
-
3. Hitherto published fusulinid data from the Tengchong Block include eight genera in the early Permian and 10 genera in the middle Permian. The fusulinids exhibit Gondwana affinity of low diversity and impoverished Pseudoschwagerininae, Verbeekininae, and Neoschwagerininae. They further indicate that the block was probably located at the higher latitude area of the southern transitional zone.
Acknowledgments
X. H. An and P. Zong participated in the fieldwork. We thank P. H. Muller for offering information. We are grateful to J. Groves and another anonymous reviewer for their valuable suggestions and comments. This research is supported by the State Program of Overseas Study for Outstanding Young Scholars of Jiangsu, the National Natural Science Foundation of China (No. 41272043, 41372008), State Key Laboratory of Palaeobiology and Stratigraphy (Nanjing Institute of Geology and Palaeontology, CAS) (No. 133110), and the Fundamental Research Funds for the Central Universities (No. 20620140390).






