



Introduction
The Neoproterozoic to lower Cambrian Chilhowee Group is exposed in the western Blue Ridge and the Valley and Ridge provinces of the broader Appalachian Mountains from Alabama to Pennsylvania (Figs. 1, 2), and provides a record of the early evolution of the Iapetan margin of the Laurentian paleocontinent (Thomas, 1977, Reference Thomas2014; Mack, Reference Mack1980; Bond et al., Reference Bond, Nickeson and Kominz1984; Simpson and Sundberg, Reference Simpson and Sundberg1987; Simpson and Eriksson, 1989, Reference Simpson and Eriksson1990). The Chilhowee Group has received much study in terms of sedimentology, facies analysis, and basin analysis (e.g., King and Ferguson, Reference King and Ferguson1960; Whisonant, Reference Whisonant1974; Mack, Reference Mack1980; Cudzil and Driese, Reference Cudzil and Driese1987; Simpson and Eriksson, 1989, Reference Simpson and Eriksson1990; Walker et al., Reference Walker, Simpson and Driese1994; Hageman and Miller, Reference Hageman and Miller2016), and has been used in continental- and global-scale correlations of the Cambrian and of the Precambrian-Cambrian boundary (e.g., Walcott, Reference Walcott1891; Resser, Reference Resser1933; Howell et al., Reference Howell, Bridge, Deiss, Edwards, Lochman, Raasch and Resser1944; Wood, Reference Wood1969). However, metazoan body fossils—including trilobites, which form the primary basis for the biostratigraphic zonation and correlation of lower Cambrian Laurentian strata (e.g., Fritz, Reference Fritz1972; Palmer, Reference Palmer1998; Hollingsworth, Reference Hollingsworth2011; Webster, Reference Webster2011; Webster and Bohach, Reference Webster and Bohach2014; Webster and Landing, Reference Webster and Landing2016)—are rare within the Chilhowee Group. Consequently, correlations are based almost exclusively on lithological similarities (e.g., Palmer, Reference Palmer1971; Mack, Reference Mack1980), and ages of the rock units and the timing of geologic events associated with the rift-to-drift transition along the continental margin are relatively poorly constrained. The discovery of biostratigraphically useful fossils within the Chilhowee Group is therefore important.
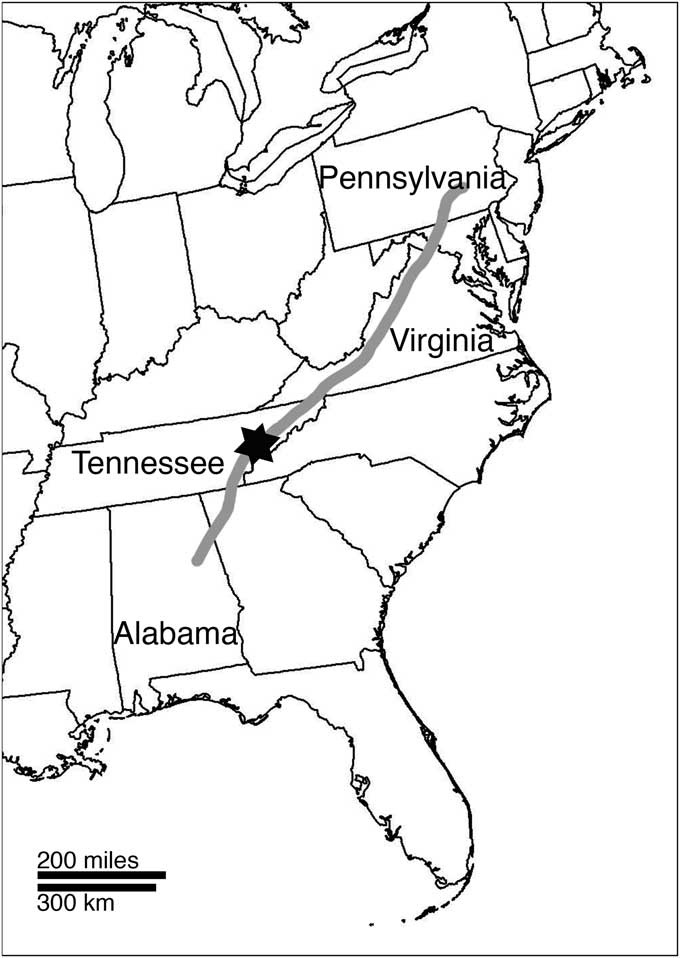
Figure 1 Map of eastern U.S.A. showing trend of Ediacaran to lower Cambrian Chilhowee Group (gray shading) in southern and central Appalachians. Star symbol indicates location of Chilhowee Mountain, Blount County, Tennessee, where the fossils discussed herein were collected.
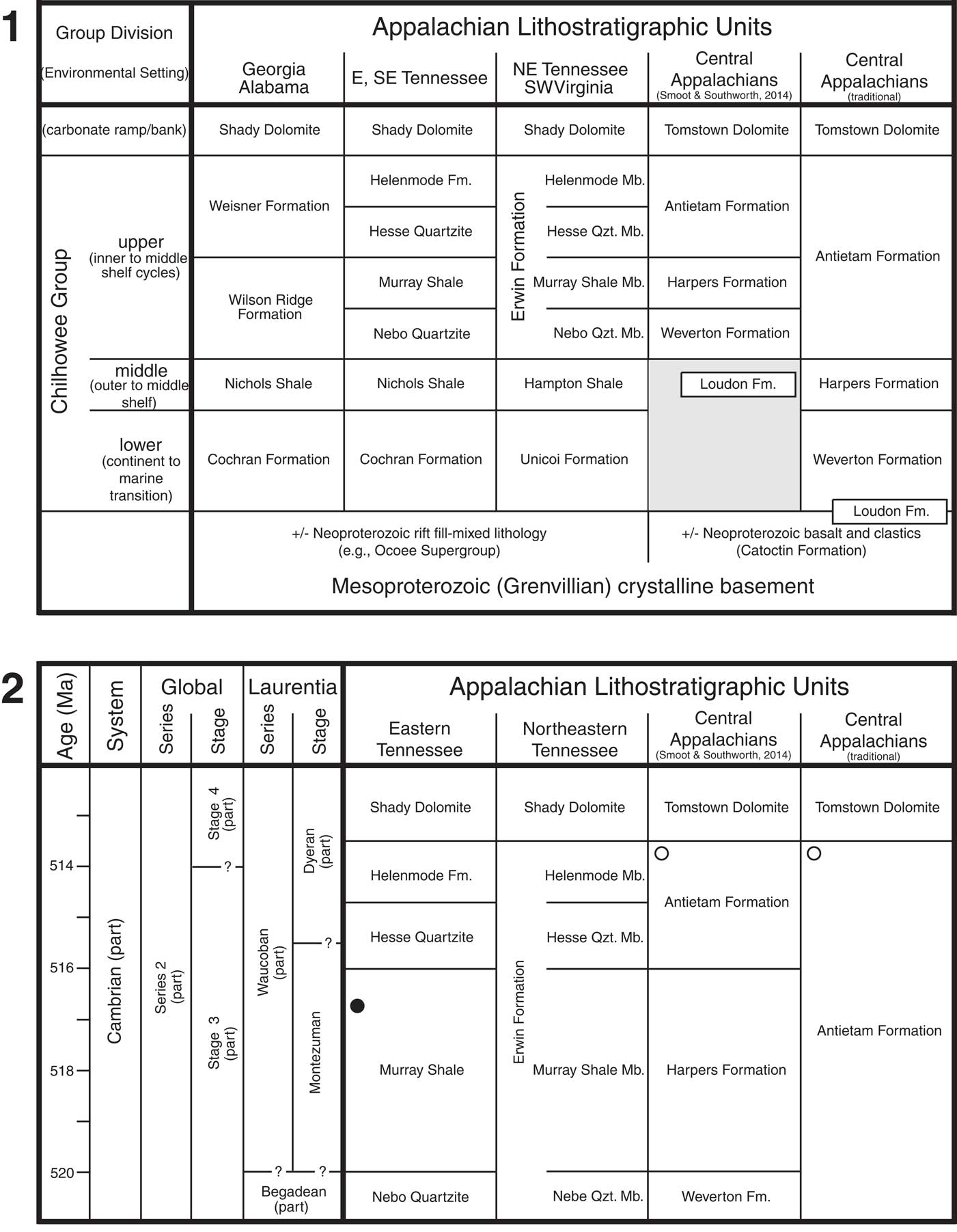
Figure 2 Lithostratigraphic correlations for southern and central Appalachians. Central Appalachians includes central and northern Virginia (approximately north of Roanoke), Maryland, and Pennsylvania. (1) Chilhowee Group plus immediately subjacent and superjacent units. Traditional interpretation of correlation for central Appalachians follows most workers (e.g., King, Reference King1949; King and Ferguson, Reference King and Ferguson1960; Mack, Reference Mack1980; Cudzil and Driese, Reference Cudzil and Driese1987; Walker and Driese, Reference Walker and Driese1991; Walker et al., Reference Walker, Simpson and Driese1994); alternative hypothesis from Smoot and Southworth (Reference Smoot and Southworth2014; gray shaded region indicates marked unconformity). Vertical scale arbitrary and non-uniform. Ediacaran–Cambrian boundary is likely in upper one-third of Unicoi Formation (Walker and Driese, Reference Walker and Driese1991; Hageman and Miller, Reference Hageman and Miller2016), but age of base of Chilhowee Group in Tennessee is poorly constrained. (2) Working hypothesis of correlation and approximate ages of lithostratigraphic units of the upper Chilhowee Group. Circles indicate stratigraphic intervals within the upper Chilhowee Group that have yielded trilobites. Age assignment of Murray Shale in eastern Tennessee based on discovery of Buenellus chilhoweensis n. sp. (black circle), as described in present study. White circles indicate trilobite occurrences in uppermost Chilhowee Group of central Appalachians (see text). Laurentian series and stage subdivisions of Cambrian follow Palmer (Reference Palmer1998); Begadean and Waucoban series together represent the traditional “lower Cambrian” of this paleocontinent. Age in millions of years before present (Ma) and potential placement of global Cambrian Stage 3-Stage 4 boundary taken from provisional Cambrian global correlation charts presented by Peng et al. (Reference Peng, Babcock and Cooper2012). Abbreviations: Fm., Formation; Mb., Member; Qzt., Quartzite.
Lower Cambrian trilobites have been previously reported from two stratigraphic intervals within the Chilhowee Group. The stratigraphically lower occurrence was reported from the Murray Shale on Chilhowee Mountain, Blount County, eastern Tennessee (Figs. 1, 2; Walcott, Reference Walcott1890, Reference Walcott1891; Keith, Reference Keith1895); that unit is the focus of the present paper. The stratigraphically higher occurrence was reported from the upper part of the Antietam Formation at several localities in Virginia, Maryland, and Pennsylvania (Fig. 2.2, white circles; Walcott, 1892, 1896, Reference Walcott1910; Bassler, Reference Bassler1919; Resser, Reference Resser1938; Butts, Reference Butts1940; Stose and Stose, Reference Stose and Stose1944; Amsden, Reference Amsden1951); those younger trilobites will be the focus of a separate study. All trilobites from the Chilhowee Group were initially identified as “Olenellus sp.” (Walcott, Reference Walcott1890, Reference Walcott1891, Reference Walcott1896, Reference Walcott1910; Resser, Reference Resser1938), and later workers have uncritically accepted that generic identification. Historically, the genus name Olenellus Hall in Billings, Reference Billings1861 was applied so broadly that its stratigraphic range spanned the entire Dyeran Stage and even down into the preceding Montezuman Stage (provisional Cambrian Stages 4 and 3, both in part; Peng et al., Reference Peng, Babcock and Cooper2012) (e.g., Walcott, Reference Walcott1910; stratigraphic divisions for the Cambrian of Laurentia follow Palmer, Reference Palmer1998). However, subsequent systematic revisions have greatly restricted the inclusivity of the genus (e.g., Palmer and Repina, Reference Palmer and Repina1993; Palmer and Repina in Whittington et al., Reference Whittington, Chatterton, Speyer, Fortey, Owens, Chang, Dean, Jell, Laurie, Palmer, Repina, Rushton, Shergold, Clarkson, Wilmot and Kelly1997; Lieberman, 1998, Reference Lieberman1999). With the recent reassignment of many species of “Olenellus” sensu lato to other genera, occurrences of Olenellus sensu stricto are apparently restricted to the mid- and upper Dyeran (provisional Cambrian Stage 4; Peng et al., Reference Peng, Babcock and Cooper2012) (Webster, Reference Webster2011 and references therein; Webster and Bohach, Reference Webster and Bohach2014). The historical records of “Olenellus” within the Chilhowee Group must, therefore, be re-evaluated in light of modern systematics in order to exploit their full biostratigraphic potential. Unfortunately, re-evaluation of the Murray Shale record is hampered by: (1) the absence of any description or illustration of specimens; and (2) the failure of subsequent workers to collect any additional trilobite material, despite concerted efforts. The lack of success is due in part to poor and confusing descriptions of field localities (see below) and the apparent rarity of specimens. Indeed, several workers have expressed doubt regarding the supposed stratigraphic provenance of the material reported by Walcott and Keith (see below).
In 2016, SJH discovered an exposure of the Murray Shale on Chilhowee Mountain that yielded a cephalon of an olenelline trilobite. This exposure is located in one of the general areas described by Walcott (Reference Walcott1890) as a source for his initial fossil discoveries, and might even represent a re-discovery of the original fossil-bearing locality (Hageman and Miller, Reference Hageman and Miller2016; see below). Hageman and Miller (Reference Hageman and Miller2016, p. 146, fig. 7d) briefly documented the discovery of the locality and illustrated the new specimen, but no formal description of the taxon was provided. Subsequent visits to the locality yielded several additional specimens. Herein, we provide a formal description of that trilobite—named Buenellus chilhoweensis n. sp.—and review other body fossil occurrences within the Murray Shale. We demonstrate that Buenellus chilhoweensis n. sp. is the oldest known trilobite from the Iapetan margin of Laurentia, and we discuss the significance of the trilobite in terms of the much-needed biostratigraphic constraint it provides on the timing of events during the early evolution of that margin.
Geologic setting, lithostratigraphy, and age of the Chilhowee Group
Following the late Neoproterozoic breakup of Rodinia, the newly formed Iapetan margin of Laurentia evolved from a tectonically active rift margin to a passive, thermally subsiding margin (Rankin, Reference Rankin1976; Thomas, Reference Thomas1977). The Ediacaran through lower Cambrian stratigraphic succession of the southern and central Appalachians records this rift-to-drift transition (Figs. 1, 2; Thomas, 1977, Reference Thomas2014; Mack, Reference Mack1980; Bond et al., Reference Bond, Nickeson and Kominz1984; Simpson and Sundberg, Reference Simpson and Sundberg1987; Simpson and Eriksson, 1989, Reference Simpson and Eriksson1990). The extensional rift phase is represented in Tennessee by the Neoproterozoic Ocoee Supergroup, which is a sequence of turbidites and mass flow deposits that accumulated in a large intracratonic rift basin (Tull et al., Reference Tull, Allison, Whiting and John2010; Thomas, Reference Thomas2014 and references therein). The overlying Chilhowee Group represents the basal siliciclastic portion of the initial transgressive depositional cycle (Sauk Sequence; Sloss, Reference Sloss1963) that blanketed the Iapetan margin during the thermal subsidence phase. Although sedimentary facies are laterally variable in thickness and composition (Walker et al., Reference Walker, Simpson and Driese1994), and stratigraphic nomenclature varies from region to region (Mack Reference Mack1980), the Chilhowee Group can be considered in three successive packages.
The lower Chilhowee Group, 400–1200 m thick, consists of the laterally equivalent Unicoi and Cochran formations in Tennessee and southwestern Virginia (Fig. 2.1). The Weverton Formation of northern Virginia, Maryland, and Pennsylvania has usually been considered to be a northern lateral equivalent of the lower Chilhowee Group (e.g., King, Reference King1949; King and Ferguson, Reference King and Ferguson1960; Cudzil and Driese, Reference Cudzil and Driese1987; Walker and Driese, Reference Walker and Driese1991), but has recently been proposed to correlate to the younger Nebo Quartzite (Smoot and Southworth, Reference Smoot and Southworth2014). The lower Chilhowee Group formed as coalescing alluvial fans, braided stream, and overbank floodplain deposits with local mudflows in fluvial, deltaic, to shallow marginal marine environments (Mack, Reference Mack1980; Simpson and Eriksson, 1989, Reference Simpson and Eriksson1990; Tull et al., Reference Tull, Allison, Whiting and John2010; Smoot and Southworth, Reference Smoot and Southworth2014). Undated amygdaloidal basalt flows are locally present in the braidplain sediments (lower and middle) part of the Unicoi Formation in northeastern Tennessee and southwestern Virginia, but the upper Unicoi Formation probably represents an early phase of transgressive sedimentation on a passive margin (Simpson and Eriksson, Reference Simpson and Eriksson1989; Walker and Driese, Reference Walker and Driese1991; Smoot and Southworth, Reference Smoot and Southworth2014). Synrift volcanics of the Catoctin Formation (underlying the Weverton formations in Virginia, Maryland, and Pennsylvania) have radiometric ages of 572±5 to 564±9 Ma (Aleinikoff et al., Reference Aleinikoff, Zartman, Walter, Rankin, Lyttle and Burton1995), and are therefore late Ediacaran in age. Although speculated upon (King and Ferguson, Reference King and Ferguson1960; Smoot and Southworth, Reference Smoot and Southworth2014), correlative relationships between the Catoctin and Unicoi basalts have not been established.
Compressed carbonaceous tubes within the middle Unicoi Formation are similar to problematic fossils found in Ediacaran assemblages elsewhere (Hageman and Miller, Reference Hageman and Miller2016). Trace fossils suggest that the Ediacaran-Cambrian boundary lies within the upper portion of the Unicoi Formation (Walker and Driese, Reference Walker and Driese1991; Hageman and Miller, Reference Hageman and Miller2016).
The middle Chilhowee Group, a 200–800 m thick succession of sand, silt, and shale, is mapped as the laterally equivalent Nichols Shale and Hampton Shale in Tennessee and southern Virginia (Fig. 2.1). The Harpers Formation of northern Virginia, Maryland, and Pennsylvania has usually been considered to be a northern lateral equivalent of the middle Chilhowee Group (e.g., King, Reference King1949; King and Ferguson, Reference King and Ferguson1960; Cudzil and Driese, Reference Cudzil and Driese1987; Walker and Driese, Reference Walker and Driese1991), but has recently been proposed to correlate to the younger Murray Shale (Smoot and Southworth, Reference Smoot and Southworth2014; see also Bloomer and Werner, Reference Bloomer and Werner1955). (Smoot and Southworth [Reference Smoot and Southworth2014] instead suggested that the Loudon Formation of Maryland and Pennsylvania is age-equivalent to the Nichols and Hampton shales [Fig. 2.1].) The contact between the lower and middle Chilhowee Group appears to be conformable (Mack, Reference Mack1980). The middle Chilhowee Group represents a marine transgression in a prodeltaic to low-energy mud shelf setting that was episodically affected by storms (Walker and Driese, Reference Walker and Driese1991). A thick black mudstone interval within the lower part of the Nichols Shale of Tennessee was deposited during the time of maximum flooding; the rest of the Nichols Shale represents a highstand systems tract (Mack, Reference Mack1980; Simpson and Eriksson, Reference Simpson and Eriksson1990; Tull et al., Reference Tull, Allison, Whiting and John2010). Trace fossil assemblages from the middle Chilhowee Group indicate that the Cambrian Substrate (Agronomic) Revolution had initiated (Hageman and Miller, Reference Hageman and Miller2016). However, searches for body fossils have met with little or no success (Laurence and Palmer, Reference Laurence and Palmer1963; Neuman and Nelson, Reference Neuman and Nelson1965; Appendix): only a single, fragmentary, conical shelly fossil of uncertain affinity has been reported (Simpson and Sundberg, Reference Simpson and Sundberg1987), and the biogenicity of even that specimen has been questioned (Hageman and Miller, Reference Hageman and Miller2016).
The upper Chilhowee Group is a siliciclastic succession that accumulated on a passive margin. Eustatic sea level control on sedimentation is evident in the form of two transgressive sequences (Tull et al., Reference Tull, Allison, Whiting and John2010; Smoot and Southworth, Reference Smoot and Southworth2014; Hageman and Miller, Reference Hageman and Miller2016). In southern and eastern Tennessee, the first of these transgressive sequences is represented by the Nebo Quartzite and overlying Murray Shale (Fig. 2). The Nebo Quartzite contains abundant Skolithos burrows (King, Reference King1949; King and Ferguson, Reference King and Ferguson1960; Neuman and Nelson, Reference Neuman and Nelson1965; Appendix), but nothing of highly refined biostratigraphic utility. The contact between the Nebo Quartzite and the Murray Shale is transitional, with some interbedding of lithologies (Whisonant, Reference Whisonant1974). Laurence and Palmer (Reference Laurence and Palmer1963, p. C53) noted that at Murray Gap on Chilhowee Mountain (see below and Appendix) the Murray Shale is 107 m (350 ft) thick and consists of three units of roughly equal thickness: “a lower unit consisting of bluish-gray noncalcareous shale with scattered quartz grains and muscovite flakes up to about 1 mm across and occasional biotite flakes and glauconite grains; a middle unit which is principally a dark-gray muscovite-bearing fine siltstone and which, when weathered, yields buff chips similar to the weathered shale of the bottom unit; and an upper unit consisting of siltstone, shale, and fine-grained sandstone with many glauconitic layers.” Rb-Sr dating of glauconite grains within the Murray Shale indicates an age of 539±30 Ma (Walker and Driese, Reference Walker and Driese1991 [recalibrating the work of Hurley et al., Reference Hurley, Cormier, Hower, Fairbairn and Pinson1960]; see Holmes,Reference Holmes1959 and Cowie, Reference Cowie1964 for earlier dating efforts). Fossils from the Murray Shale are the main focus of this paper and are discussed in following sections. The interval of maximum transgression is located within the Murray Shale.
The Murray Shale is in sharp but conformable contact with the overlying Hesse Quartzite (Whisonant, Reference Whisonant1974); this transition represents a return to shallow-water wave and tidally influenced conditions. The Hesse Quartzite is a quartz sandstone that contains Skolithos burrows (Neuman and Nelson, Reference Neuman and Nelson1965; Hageman and Miller, Reference Hageman and Miller2016). The second transgressive sequence is represented by the Hesse Quartzite and overlying Helenmode Formation (a quartz siltstone and sandstone with interbedded shale) (Fig. 2).
In northeastern Tennessee these same four successive lithostratigraphic units (Nebo Quartzite, Murray Shale, Hesse Quartzite, and Helenmode) are recognized as members within the Erwin Formation (e.g., King and Ferguson, Reference King and Ferguson1960; Walker and Driese, Reference Walker and Driese1991; Fig. 2). North of central Virginia, the fine-grained sediments of the Murray Shale have typically been considered to be absent and the facies of the Nebo and Hesse quartzites have been interpreted to merge and thicken, so that the entire upper Chilhowee Group is represented primarily by quartz sandstone mapped as the Antietam Formation (e.g., King, Reference King1949; King and Ferguson, Reference King and Ferguson1960; Cudzil and Driese, Reference Cudzil and Driese1987; Walker and Driese, Reference Walker and Driese1991; Fig. 2, right hand column). However, Smoot and Southworth (Reference Smoot and Southworth2014) proposed an alternative model for correlation whereby the Weverton Formation represents the northward lateral equivalent of the Nebo Quartzite Member, the Harpers Formation represents the northward lateral equivalent of the Murray Shale Member, and the Antietam Formation is the northward lateral equivalent of just the Hesse Quartzite plus Helenmode members (see also Bloomer and Werner, Reference Bloomer and Werner1955). Under that alternative model (Smoot and Southworth, Reference Smoot and Southworth2014; Fig. 2, column second from right), localities north of Tennessee record a more obvious sedimentary expression of the two transgressive sequences within the upper Chilhowee Group.
The upper part of the Antietam Formation (Helenmode equivalent) contains trilobite fragments that have historically been identified as “Olenellus sp.” (Walcott, 1892, 1896, Reference Walcott1910; Bassler, Reference Bassler1919; Resser, Reference Resser1938, pl. 2, fig. 23; Butts, Reference Butts1940; Stose and Stose, Reference Stose and Stose1944; Amsden, Reference Amsden1951). However, those specimens must undergo modern systematic revision before their biostratigraphic significance can be determined (MW and SJH, in preparation). The Helenmode Formation and its lateral equivalents also contain hyoliths and brachiopods (Neuman and Nelson, Reference Neuman and Nelson1965; Hageman and Miller, Reference Hageman and Miller2016).
The second transgression ultimately resulted in the development of a carbonate bank that extended along the passive margin from present-day Alabama to Pennsylvania and the northern Appalachians (Landing, Reference Landing2012). Development of the carbonate bank in the southern and central Appalachians is reflected in the conformable transition from the upper Chilhowee Group into the overlying Shady Dolomite and its lateral equivalents the Tomstown Dolomite, Jumbo Dolomite, and Murphy Marble (Fig. 2; Bloomer and Werner, Reference Bloomer and Werner1955; Mack, Reference Mack1980; Simpson and Eriksson, Reference Simpson and Eriksson1990; Tull et al., Reference Tull, Allison, Whiting and John2010). The carbonates have been well studied at localities such as Sleeping Giants, Alabama (Bearce and McKinney, Reference Bearce and McKinney1977), and near Austinville, Virginia (Balsam, Reference Balsam1974; Pfeil and Read, Reference Pfeil and Read1980; Barnaby and Read, Reference Barnaby and Read1990; McMenamin et al., Reference McMenamin, Debrenne and Zhuravlev2000). Locally, the carbonates contain a rich fauna of trilobites, archaeocyathids, brachiopods, Salterella Billings, Reference Billings1861, hyoliths, and echinoderm plates (Resser, Reference Resser1938; Butts, Reference Butts1940; Yochelson, Reference Yochelson1970; McMenamin et al., Reference McMenamin, Debrenne and Zhuravlev2000; Tull et al., Reference Tull, Allison, Whiting and John2010). Faunas of the Shady and Tomstown dolomites indicate a mid-Dyeran age (McMenamin et al., Reference McMenamin, Debrenne and Zhuravlev2000; MW unpublished observations).
Previous paleontological work on the Murray Shale
Biostratigraphic data from the subjacent and superjacent units (summarized above) constrain the Murray Shale to be of early Cambrian, and no younger than mid-Dyeran, age. According to the current working hypothesis of the Cambrian time scale (Peng et al., Reference Peng, Babcock and Cooper2012), this indicates a numerical age somewhere between 541 Ma and ~514 Ma. This is congruent with the coarse age constraint imposed by radiometric dating of glauconite grains from the unit (539±30 Ma; Walker and Driese, Reference Walker and Driese1991). However, the uncertainty in age associated with these constraints is large, at least in comparison to the high-resolution biostratigraphic framework that exists for the early Cambrian of the Cordilleran margin of Laurentia (e.g., Hollingsworth, Reference Hollingsworth2011; Webster, Reference Webster2011; Webster and Bohach, Reference Webster and Bohach2014). This section reviews previous paleontological discoveries within the Murray Shale and discusses the extent to which those finds refine the age estimate for the unit.
Initial fossil discoveries
Fossils from the Chilhowee Group were first found by Cooper Curtice during the geological resurvey of eastern Tennessee (noted by Walcott, Reference Walcott1891). The nature of Curtice’s fossils is nowhere mentioned, but they were apparently found “in the shales interbedded in the quartzite of Chilhowee Mountain” (Walcott, Reference Walcott1891, p. 302). Curtice’s discovery presumably occurred in 1885 (see Yochelson and Osborne, Reference Yochelson and Osborne1999), although the significance of the find might not have been immediately realized: Walcott (Reference Walcott1889, table on p. 386) reported that fossils were unknown from the lower Cambrian of Tennessee. Nevertheless, in 1889, Walcott visited Chilhowee Mountain and discovered body fossils in the “banded shales at and near the summit of the [Chilhowee Group]” that allowed him to determine an early Cambrian age for the unit (Walcott, Reference Walcott1890, p. 536–537). Walcott (Reference Walcott1890, p. 570, 583; 1891, p. 154) listed these fossils as a hyolith, the arthropod Isoxys chilhoweanus Walcott, Reference Walcott1890 (as Isoxys chilhoweana), an ostracod crustacean, and “an undetermined species of Olenellus.” In the associated table of fossil occurrences, Walcott (Reference Walcott1890, table on p. 575) tentatively identified the trilobite as Olenellus thompsoni (Hall, Reference Hall1859), and later referred to it as “a species of Olenellus closely allied to Olenellus thompsoni and O. asaphoides in that portion of the head preserved” (Walcott, Reference Walcott1891, p. 154; “O. asaphoides” is now Elliptocephala asaphoides Emmons, Reference Emmons1844). That faunal list—in part or in full—has been repeated many times in the literature by subsequent workers (e.g., Resser, 1933, Reference Resser1938; Grabau, Reference Grabau1936; King et al., Reference King, Hadley and Neuman1952; King and Ferguson, Reference King and Ferguson1960; Neuman and Nelson, Reference Neuman and Nelson1965), but very few of the fossils from Walcott’s collection have been illustrated. Two hyolith specimens were figured by Resser (Reference Resser1938, pl. 4, figs. 30, 31; USNM 18447, from lot USNM 26979). Two specimens of Isoxys chilhoweanus were figured by Walcott (Reference Walcott1890, pl. 80, figs. 10, 10a; also Williams et al., Reference Williams, Siveter and Peel1996, fig. 7.2) and nine specimens currently reside in the USNM (lots USNM 18444 and USNM 18445, including the holotype). The “ostracod crustacean” mentioned by Walcott (Reference Walcott1891, p. 154) is the bradoriid Indota tennesseensis (Resser, Reference Resser1938), first named and illustrated as Indianites tennesseensis by Resser (Reference Resser1938, p. 107, pl. 3, fig. 47; holotype USNM 94759; for subsequent taxonomic revisions see Siveter and Williams, Reference Siveter and Williams1997). The trilobite mentioned by Walcott (1890, Reference Walcott1891) was never illustrated or described. This is unfortunate, because the historical identification of this trilobite as “Olenellus sp.” has been used to support a Dyeran age for the Murray Shale (e.g., Simpson and Sundberg, Reference Simpson and Sundberg1987), but such age assignment cannot be substantiated without modern systematic treatment of the taxon. The sole known specimen from Walcott’s original trilobite collection is illustrated for the first time herein (Fig. 4.8). This specimen does not represent a species of Olenellus, but instead a new species of a much older olenelline genus (described herein).
Walcott’s fossils were collected from two areas on Chilhowee Mountain—Little River Gap and near Montvale Springs (Walcott, Reference Walcott1890, p. 626; Fig. 3.1, 3.2)—but the exact locations of the fossil-bearing sites are unclear (see Appendix). The stratigraphic provenance of the fossils is also ambiguous. Walcott reported only that they were found “in the shale about 20 feet above the quartzite in the upper shale bed” (Walcott, Reference Walcott1891, p. 302), and that Skolithos burrows were present in the sandstone underlying the fossil-bearing shale (Walcott, Reference Walcott1890, p. 626; 1891, p. 154). Those descriptions could apply to either the Murray Shale (above the Nebo Quartzite) or a shaly interval within the Helenmode Formation (above the Hesse Quartzite).
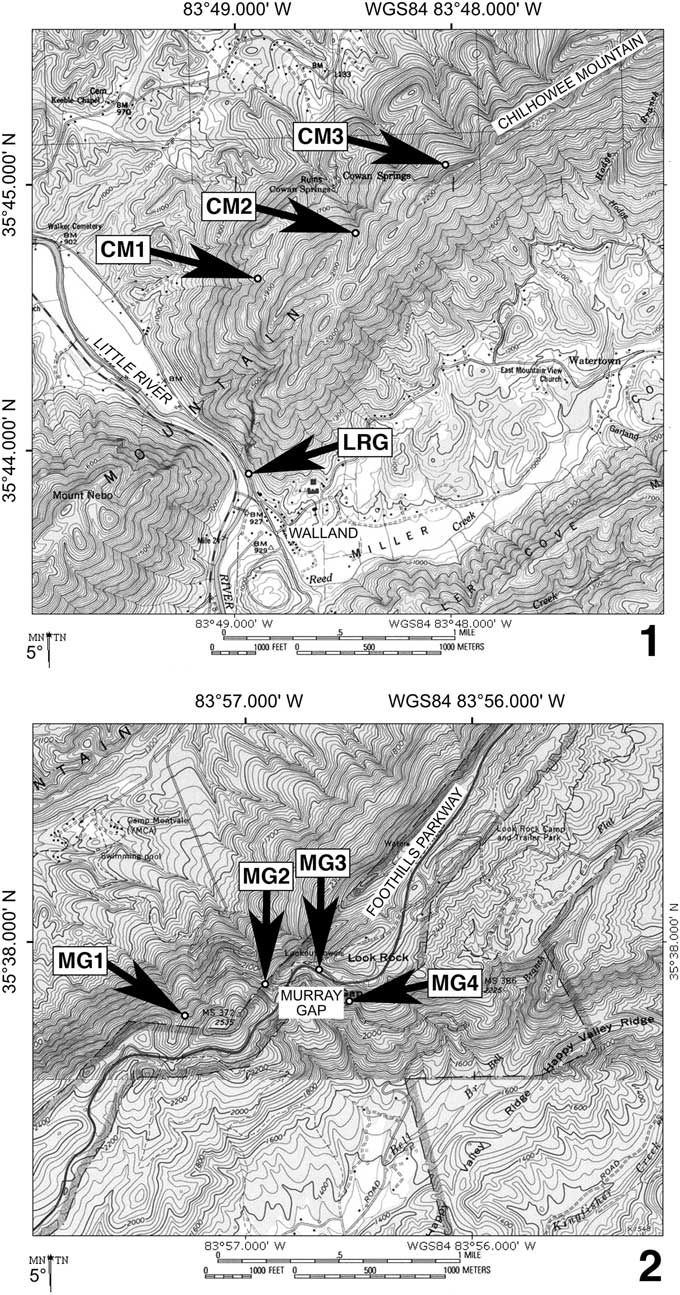
Figure 3 Maps for localities on Chilhowee Mountain, Blount County, Tennessee, U.S.A., discussed in text. (1) Little River Gap area, near Walland. (2) Murray Gap area, near Montvale Springs. Walland (in 1) is located ~15.3 km (9.5 miles) northeast of Murray Gap (in 2); general location of these two maps within Tennessee shown by star symbol in Figure 1. Locality abbreviations: CM, newly discovered fossiliferous exposures on Chilhowee Mountain, including within Nichols Shale (CM1), lowest few meters of Murray Shale (CM2), and Buenellus-bearing site within Murray Shale (CM3); LRG, classic Little River Gap roadside exposure; MG1, base of Murray Shale exposed alongside disused bridleway; MG2 and MG3, roadcuts through Murray Shale collected by Laurence and Palmer (Reference Laurence and Palmer1963) and Wood and Clendening (Reference Wood and Clendening1982); MG4, roadside exposure at intersection of Happy Valley Road and Flats Road. Maps created with TOPO! software (©National Geographic Society, 2002).
In 1893 Walcott revisited Chilhowee Mountain with mapper Arthur Keith (Laurence and Palmer, Reference Laurence and Palmer1963; Yochelson, Reference Yochelson1998), and the two collected additional fossils. Keith’s (Reference Keith1895) report was the first to explicitly state that fossils had been found in the Murray Shale. He (Keith, Reference Keith1895, p. 3) noted that brachiopods and trilobites had been discovered in the Murray Shale “on the east side of Little River Gap and on the crest of the mountain above Montvale Springs,” the latter site being the present-day Murray Gap area (see Appendix). He did not specify which of the two localities yielded the trilobites and which yielded the brachiopods, or whether both fossil types occurred at each locality. Museum documentation shows that Walcott’s original trilobite (Fig. 4.8) was sourced from the Little River Gap area. Brachiopods had not been mentioned in Walcott’s (1890, Reference Walcott1891) original faunal lists, so that occurrence appears to have been a novel find of the 1893 trip. More (and perhaps better) bradoriid specimens were also found on the 1893 trip: Laurence and Palmer (Reference Laurence and Palmer1963, p. C54) found museum documentation stating that the bradoriid specimens described by Resser (Reference Resser1938) “were collected by Walcott and Keith in 1893.”

Figure 4 Buenellus chilhoweensis n. sp. from the Murray Shale, Chilhowee Mountain, Blount County, Tennessee, U.S.A. (1) Internal mold of cephalon from ICS-10567, USNM 633932; (2) internal mold of cephalon from ICS-10568, USNM 645832.; (3, 4) internal and external mold, respectively, of holotype cephalon from ICS-10567, USNM 645831; (5, 6) internal and external mold, respectively, of cephalon from ICS-10568, USNM 645833; (7) external mold of cephalon from ICS-10568, USNM 645834; (8) latex peel of external mold of incomplete cephalon found by Walcott in 1889 and mentioned by Walcott (1890, Reference Walcott1891), USNM 18446, dorsal view. (1–7) from upper part of Murray Shale, locality CM3; (8) from Little River Gap area (USNM Locality 17). Scale bars 5 mm.
The purported age and source of Walcott’s and Keith’s collections were subsequently cast into doubt by several authors. Referring to the collection of olenelline trilobites and Isoxys chilhoweanus from the Little River Gap locality, Resser (Reference Resser1933, p. 746) stated that “(t)he circumstances surrounding the collection of these fossils cause some doubt as to their stratigraphic position.” Skepticism over the stratigraphic provenance of the fossils was repeated by Grabau (Reference Grabau1936). Resser (Reference Resser1938, p. 25) later stated that the material from the Little River Gap and above Montvale Springs was of “uncertain age” because the genera—therein listed as Hyolithes Eichwald, Reference Eichwald1840; Isoxys Walcott, Reference Walcott1890; and Indianites Ulrich and Bassler, Reference Ulrich and Bassler1931—“appear rather to be Middle Cambrian” (the occurrence of the diagnostically early Cambrian olenelline trilobite was not mentioned).
Stose and Stose (Reference Stose and Stose1944) expressed doubt as to whether the collections mentioned by Keith (Reference Keith1895) came from the Murray Shale. Their skepticism stemmed from ambiguities over the mapping in the Little River Gap and from the fact that all other Chilhowee Group fossils had otherwise been reported only from the uppermost beds marking the transition into the Shady Dolomite (see references above). Those observations led Stose and Stose (Reference Stose and Stose1944, p. 388) to hypothesize that the fossils from the Little River Gap locality might have been sourced from the “transitional beds at the top of the Hesse” (i.e., the Helenmode Formation) rather than the Murray Shale. That hypothesis was subsequently repeated by King (Reference King1949, p. 520). Later, King et al. (Reference King, Hadley and Neuman1952, p. 15; also King and Ferguson, Reference King and Ferguson1960) explicitly stated that Walcott’s collections from Little River Gap had actually been sourced from the Helenmode Formation rather than the Murray Shale, a conclusion also reached by Neuman and Nelson (Reference Neuman and Nelson1965, p. D28–D29) (see Appendix for further details). However, despite his reservations over the stratigraphic provenance of the fossils from Little River Gap, King (Reference King1949, p. 520) acknowledged that the fossil collection from the crest of Chilhowee Mountain above Montvale Springs mentioned by Keith (Reference Keith1895) was almost certainly from the Murray Shale and represented the stratigraphically oldest occurrence of trilobites and brachiopods in the southern Appalachians. The occurrence of Isoxys chilhoweanus at both sites (Walcott, Reference Walcott1890, p. 626) could be construed as biostratigraphic support for the Little River Gap fossils having also been sourced from the Murray Shale (contra the concerns reviewed above), but it is also possible that Isoxys chilhoweanus has a long stratigraphic range that spans both the Murray Shale and the Helenmode Formation. Our new discoveries (below; Appendix) demonstrate that Walcott’s original fossil collection at Little River Gap was indeed made in the Murray Shale.
Subsequent fossil discoveries
Since those initial discoveries in the 1880s and 1893, several workers have searched for additional body fossils at Little River Gap and Murray Gap. For nearly seventy years such efforts were almost invariably unsuccessful (e.g., King et al., Reference King, Hadley and Neuman1952; Neuman and Nelson, Reference Neuman and Nelson1965), although Neuman and Nelson (Reference Neuman and Nelson1965) reported finding fragments of an inarticulate brachiopod in the Helenmode Formation at Little River Gap.
However, new roadcuts at Murray Gap were made in 1962 (Fig. 3.2, Localities MG2 and MG3; Appendix). The fresh roadcuts exposed much of the Murray Shale, from which additional specimens of the bradoriid Indota tennesseensis were recovered (Laurence and Palmer, Reference Laurence and Palmer1963). Those new specimens came from ~6.1–18.3 m above the base of the Murray Shale (Laurence and Palmer, Reference Laurence and Palmer1963, p. C53). Wood and Clendening (Reference Wood and Clendening1982) subsequently collected the bradoriid from 6.3–10.6 m above the base of the Murray Shale at the same locality. Tracks and trails, but no body fossils, were recovered from the overlying portion of the Murray Shale (Laurence and Palmer, Reference Laurence and Palmer1963). Acritarchs were also described from the lower part (6.3–46.7 m above the base) of the Murray Shale at those roadcuts (Wood and Clendening, Reference Wood and Clendening1982, their “locality 1”) and from a similar stratigraphic interval in a roadcut in northeasternmost Tennessee (Wood and Clendening, Reference Wood and Clendening1982, their “locality 2”).
Those discoveries offer limited biostratigraphic utility. The occurrence of the bradoriid in the new roadcuts was sufficient for Laurence and Palmer (Reference Laurence and Palmer1963) to confirm assignment of the Murray Shale to the lower Cambrian, based on the occurrence of the genus (then identified as Indiana Matthew, Reference Matthew1902) in lower Cambrian rocks elsewhere in North America and Europe. However, the Murray Shale bradoriid has subsequently been reassigned to Indota Öpik, Reference Öpik1968 (Siveter and Williams, Reference Siveter and Williams1997), a widespread genus that apparently ranges into the early middle Cambrian (Williams et al., Reference Williams, Siveter, Popov and Vannier2007). Indeed, based on tentatively proposed junior synonymies, Indota tennesseensis itself might occur in the Ordian Yelvertoft Beds of Australia (Öpik, Reference Öpik1968; Siveter and Williams, Reference Siveter and Williams1997; Jones and Laurie, Reference Jones and Laurie2006). The Ordian Stage of Australia has been hypothesized to correlate to the uppermost Dyeran and a portion of the overlying Delamaran stages of Laurentia (Kruse et al., Reference Kruse, Jago and Laurie2009; Peng et al., Reference Peng, Babcock and Cooper2012). Given that the Murray Shale can be no younger than mid-Dyeran (see above), this suggests that—if the Australian occurrence is correct and the intercontinental correlation is accurate—Indota tennesseensis must have a relatively long stratigraphic range.
Wood and Clendening (Reference Wood and Clendening1982, p. 259) documented the acritarch Medousapalla choanoklosma Wood and Clendening, Reference Wood and Clendening1982, which subsequently was recognized as a junior synonym of Skiagia ornata (Volkova, Reference Volkova1968) (see Zang, Reference Zang2001 for taxonomic revisions), from 10.6 m above the base of the Murray Shale at their Locality 1. Skiagia ornata is widespread and has a long stratigraphic range, spanning from approximately the base of the trilobite-bearing portion of the traditional lower Cambrian through into the traditional middle Cambrian (Zang, Reference Zang2001; Moczydlowska and Zang, 2006). The occurrence of this acritarch in Tennessee therefore suggests that the lower part of the Murray Shale is probably no older than the base of the Montezuman Stage.
In summary, previous work unambiguously demonstrates that the Murray Shale contains Isoxys chilhoweanus, Indotes tennesseensis, hyoliths, acritarchs, and abundant trace fossils. We herein confirm that the olenelline trilobite reported by Walcott (1890, Reference Walcott1891) was also sourced from the Murray Shale. The presence of brachiopods within the Murray Shale, as reported by Keith (Reference Keith1895), cannot be unambiguously substantiated due to vague documentation of the site(s) of collection and apparent loss of the specimens: it remains possible that they were actually sourced from the Helenmode Formation. The hitherto described and formally named fossils that were undoubtedly sourced from the Murray Shale, in combination with constraints imposed by the underlying and overlying units, suggest that the age of the base of the Murray Shale is no older than Montezuman (~520 Ma) and the top of the unit is no younger than mid-Dyeran (~514 Ma).
New paleontological work on the Murray Shale
The new trilobite-bearing locality
Recent fieldwork on Chilhowee Mountain to the northeast of the classic Little River Gap roadcut resulted in the discovery of a 2 m thick exposure of the Murray Shale (Fig. 3.1, Locality CM3; Appendix) that yielded a well-preserved cephalon of an olenelline trilobite (Hageman and Miller, Reference Hageman and Miller2016, fig. 7d) plus abundant hyoliths. Subsequent visits to the site have yielded six additional specimens of that trilobite, described below as Buenellus chilhoweensis n. sp. (see Systematic Paleontology section).
The trilobite-bearing exposure is located in the bank of a jeep trail on an otherwise forested hillside, and attempts to measure a stratigraphic section are frustrated by vegetation and soil cover. Nevertheless, it can be ascertained that the trilobite occurs in the upper portion of the Murray Shale within a few meters of the base of the overlying Hesse Quartzite. Other fossiliferous horizons on the hillside—lower within the Murray Shale and in stratigraphically underlying units—are consistent with this determination (see Appendix).
Walcott’s original trilobite specimen (Fig. 4.8) is conspecific with Buenellus chilhoweensis n. sp. The lithology of the newly discovered trilobite-bearing site matches that of the slabs bearing Walcott’s original trilobite specimen and the type material of Isoxys chilhoweanus. It is therefore possible that Walcott’s (1890, Reference Walcott1891) “Little River Gap” locality included material sourced from the Murray Shale on the northwest-facing flank of Chilhowee Mountain northeast of Little River Gap, and maybe even from the trilobite-bearing site described herein (as was believed by Hageman and Miller, Reference Hageman and Miller2016, p. 146). Walcott’s (Reference Walcott1891, p. 302) statement that his fossils were recovered from “about 20 feet above the quartzite in the upper shale bed” offers a potentially testable means of determining whether the new site is a re-discovery (or at least a stratigraphic equivalent) of the original 1889 site. However, lack of sufficient exposure currently hinders such a test.
Refined age assignment for the Murray Shale
Buenellus chilhoweensis n. sp. is known only from the vicinity of the type locality and therefore offers no current utility for species-level correlation or biostratigraphy. However, the phylogenetic affinity of Buenellus chilhoweensis n. sp. can be used—with caveats—to indirectly constrain the age of the Murray Shale. The new species is hypothesized to be most closely related to Buenellus higginsi Blaker, Reference Blaker1988, the type and only other known species of Buenellus Blaker, Reference Blaker1988 (see Systematic Paleontology section). Buenellus higginsi is known only from the Sirius Passet Lagerstätte in the lower portion of the Buen Formation of North Greenland (Blaker and Peel, Reference Blaker and Peel1997; Babcock and Peel, Reference Babcock and Peel2007; Ineson and Peel, Reference Ineson and Peel2011). That Lagerstätte has been considered to belong to the middle to upper part of the Montezuman Stage, based on the fact that it bears a trilobite (and is therefore younger than the pre-trilobite Begadean Series) and is below strata that contain early Dyeran trilobites (Palmer and Repina, Reference Palmer and Repina1993; Blaker and Peel, Reference Blaker and Peel1997; Palmer and Repina in Whittington et al., Reference Whittington, Chatterton, Speyer, Fortey, Owens, Chang, Dean, Jell, Laurie, Palmer, Repina, Rushton, Shergold, Clarkson, Wilmot and Kelly1997; Babcock and Peel, Reference Babcock and Peel2007); acritarch biostratigraphic data are also consistent with that age assignment (summarized by Babcock and Peel, Reference Babcock and Peel2007). To the extent that such closely related species as Buenellus higginsi and Buenellus chilhoweensis n. sp. are likely to be of generally similar geologic age, the upper part of the Murray Shale is provisionally hypothesized to be of mid- to upper Montezuman age (i.e., between ~518 Ma and 515.5 Ma sensu Peng et al., Reference Peng, Babcock and Cooper2012; Fig. 2.2). A Montezuman age for Buenellus chilhoweensis n. sp. is also congruent with the biostratigraphic constraints on the age of the Murray Shale imposed by other sources of data (see above): we are unaware of any data that unambiguously contradict this age inference.
The hypothesis that the Murray Shale (or at least its upper part) is coarsely age-equivalent to the lower part of the Buen Formation comes, of course, with the non-trivial caveat that the correlation is based solely on the two lithostratigraphic units that were deposited in widely separated basins on the Iapetan and Innuitian margins of Laurentia, respectively, sharing a genus in common. A hypothesis of age-equivalence of strata is most robust when those strata are from geographically closely spaced sections and share species in common, because under such conditions the assumptions regarding isochrony of local stratigraphic ranges are less prone to dramatic violation (e.g., Landing et al., Reference Landing, Geyer, Brasier and Bowring2013). We therefore do not claim precise age-equivalence of the upper Murray Shale and lower Buen Formation within the Montezuman Stage (although such equivalence is possible), and we stress that our provisional age assignment for the Murray Shale (Fig. 2.2) is a working hypothesis that should be tested with additional data.
Materials and methods
Repositories and institutional abbreviations
All Murray Shale trilobite specimens in this study are housed in the paleobiology collection of the U.S. National Museum of Natural History (USNM). The first specimen was found in situ on the exposure; all other specimens were recovered from bulk samples extracted from the outcrop. The bulk samples were taken from a stratigraphic interval ~50 cm thick that included the horizon on which the first specimen was found. Comparative data for species belonging to other nevadioid genera were obtained through study of specimens housed within the collections of the Institute for Cambrian Studies (ICS), University of Chicago. Fossil-bearing localities within the Murray Shale are described in the Appendix.
Morphometric data
Traditional morphometric data (linear dimensions and angles) were taken from digital images of specimens (for the Tennessee material) or from scans of published images (for Buenellus higginsi). Data were collected using the ImageJ software (http://rsb.info.nih.gov/ij/index.html). Values for some variables were estimated on incompletely preserved or moderately effaced specimens, but only when those estimates were replicable within a small margin of error (typically <0.05 mm on large cephala). Values for variables relating to transverse measurements that span the sagittal line were obtained on some specimens by doubling a transverse measurement from the sagittal line to one end-point of the variable. Such estimates are designated as “approximate” values in the description. Measurement error introduced through these approximations is likely to be negligible.
Terminology
The morphological terminology applied herein largely follows that of Palmer and Repina (Reference Palmer and Repina1993) and Whittington and Kelly in Whittington et al. (Reference Whittington, Chatterton, Speyer, Fortey, Owens, Chang, Dean, Jell, Laurie, Palmer, Repina, Rushton, Shergold, Clarkson, Wilmot and Kelly1997), with modifications to olenelline terminology proposed by Webster (2007a, Reference Websterb, Reference Webster2009) and Webster and Bohach (Reference Webster and Bohach2014).
Systematic paleontology
Order Redlichiida Richter, Reference Richter1932
Suborder Olenellina Walcott, Reference Walcott1890
Superfamily “Nevadioidea” Hupé, Reference Hupé1953
Remarks
Palmer and Repina (Reference Palmer and Repina1993; also Palmer and Repina in Whittington et al., Reference Whittington, Chatterton, Speyer, Fortey, Owens, Chang, Dean, Jell, Laurie, Palmer, Repina, Rushton, Shergold, Clarkson, Wilmot and Kelly1997) included Buenellus alongside Nevadia Walcott, Reference Walcott1910, Nevadella Raw, Reference Raw1936, Cirquella Fritz, Reference Fritz1993, and Pseudojudomia Egorova in Gorjansky et al., Reference Gorjansky, Egorova and Savitskiy1964 within the Family Nevadiidae Hupé, Reference Hupé1953. That familial designation has been accepted by most other workers (e.g., Blaker and Peel, Reference Blaker and Peel1997; Jell and Adrain, Reference Jell and Adrain2003; Babcock and Peel, Reference Babcock and Peel2007). Lieberman (Reference Lieberman2001) found that taxa traditionally assigned to the Nevadiidae formed part of a paraphyletic grade between the Fallotaspidoidea Hupé, Reference Hupé1953 and [Olenelloidea Walcott, Reference Walcott1890+Judomioidea Repina, Reference Repina1979]; he termed that grade the “Nevadioidea” Hupé, Reference Hupé1953 and did not define families within it. Relationships among “nevadioids” are far from settled, however: Buenellus was not included in that cladistic analysis, for example; nor was Limniphacos Blaker and Peel, Reference Blaker and Peel1997, another possible nevadiid from North Greenland. A forthcoming, more comprehensive cladistic analysis of olenelline trilobites will resolve relationships among these taxa (Webster, in preparation). Pending publication of that new analysis, and given the uncertainty over monophyly of the traditional Nevadiidae, we herein conservatively avoid family-level assignment within the “Nevadioidea.”
Genus Buenellus Blaker, Reference Blaker1988
Type species
Buenellus higginsi Blaker, Reference Blaker1988 from the lower part of the Buen Formation, Peary Land, North Greenland, by original designation.
Other species
Buenellus chilhoweensis n. sp. (see below).
Diagnosis
(Emended from Babcock and Peel, Reference Babcock and Peel2007.) Proximal portion of posterior cephalic margin approximately transverse or weakly posterolaterally oriented when traced distally from axial furrow to adgenal angle; adgenal angle weak or absent so that base of genal spine lies slightly posterior to or opposite lateral margins of LO. Genal spine broad-based. Intergenal spine absent or reduced to small dorsal swelling on posterior cephalic border immediately distal to adgenal angle. Glabella slightly tapered anteriorly. SO deepest midway between sagittal line and axial furrow, extremely shallow or not incised adjacent to axial furrow. Cephalic border furrow, axial furrow, and glabellar furrows, especially those anterior to SO, very shallow. Short preglabellar field. Weak parafrontal band extends around lateral and anterior margins of LA. Ocular lobe narrow (tr.), anterior portion of more subdued relief than posterior portion, summit lower than LA and separated from it by break in slope; inner margin poorly defined from interocular area; posterior tip transversely opposite lateral margin of SO or L1. Interocular area slightly narrower to slightly wider (tr.) than width (tr.) of extraocular area opposite S1. Intergenal ridge and posterior ocular line converge at intergenal swelling. Fine granulations on external surface of exoskeleton. Thorax (only known from type species) of 17 or 18 segments, maintaining width or widening slightly backward to eighth segment, then tapering posteriorly. Pygidium simple; may bear one thoracic-like segment fused to anterior edge. Posterior margin of pleurae of first nine or ten segments sigmoidally curved. Pleural spines short (exsag.), tips opposite axial ring of next one or two segments.
Remarks
Buenellus was previously only known with certainty from the type species in North Greenland (see Babcock and Peel, Reference Babcock and Peel2007, p. 404, for discussion of a supposed occurrence in Novaya Zemlya). Discovery of Buenellus chilhoweensis n. sp. demonstrates that the genus occupied both the Innuitian and Iapetan margins of Laurentia. The generic diagnosis is herein emended to accommodate Buenellus chilhoweensis n. sp. and expanded to include several previously unspecified features that distinguish Buenellus from similar taxa.
Blaker and Peel (Reference Blaker and Peel1997) discussed differences between Buenellus and several other similar genera, including the nevadioids Nevadella and Nevadia, the possible nevadioid Limniphacos, the holmiid olenelloids Holmia Matthew, Reference Matthew1890 and Kjerulfia Kiaer, Reference Kiaer1917, and the problematic Callavia Matthew, Reference Matthew1897, which has been variously considered a holmiid (Palmer and Repina in Whittington et al., Reference Whittington, Chatterton, Speyer, Fortey, Owens, Chang, Dean, Jell, Laurie, Palmer, Repina, Rushton, Shergold, Clarkson, Wilmot and Kelly1997; Jell and Adrain, Reference Jell and Adrain2003) or a judomioid (Lieberman, Reference Lieberman2001). Buenellus also shares many features with Pseudojudomia egregia Egorova in Gorjansky et al., Reference Gorjansky, Egorova and Savitskiy1964, which is the type and only known species of Pseudojudomia, including details of the glabellar furrows, the nature of the contact between the ocular lobes and the anterior glabella, and a general cephalic effacement. Although it is possible that some of these shared features represent symplesiomorphies or homoplasies, it is likely that the two genera are closely related. (A formal hypothesis of their relationship will be presented in a forthcoming cladistic analysis [Webster, in preparation].) The two genera are most reliably distinguished by differences in the form of the posterior cephalic margin: in Pseudojudomia egregia the posterior cephalic margin arcs posterolaterally and uniformly curves into the inner margin of the genal spine so that the spine and the cephalic border are smoothly confluent; whereas in Buenellus the posterior cephalic margin is more transversely oriented (often with a slight anterior deflection at the adgenal angle) when traced abaxially and there is a more distinct (although certainly not sharply) curved geniculation marking where the genal spine contacts the posterior cephalic border. Other publications relevant to the diagnosis and validity of the genus Buenellus include: Blaker (Reference Blaker1988, p. 34–35), Palmer and Repina (Reference Palmer and Repina1993, p. 31), Blaker and Peel (Reference Blaker and Peel1997, p. 50–52), Palmer and Repina in Whittington et al. (Reference Whittington, Chatterton, Speyer, Fortey, Owens, Chang, Dean, Jell, Laurie, Palmer, Repina, Rushton, Shergold, Clarkson, Wilmot and Kelly1997, p. 428), Jell and Adrain (Reference Jell and Adrain2003, p. 353, 474), and Babcock and Peel (Reference Babcock and Peel2007, p. 411–412).
Buenellus chilhoweensis new species
1890 undetermined species of Olenellus; Reference WalcottWalcott, p. 570.
1890 Olenellus thompsoni?; Reference WalcottWalcott, table on p. 575 (eastern Tennessee occurrence).
1890 Olenellus, sp.?; Reference WalcottWalcott, p. 583.
1891 species of Olenellus closely allied to Olenellus thompsoni and Olenellus asaphoides in that portion of the head preserved; Reference WalcottWalcott, p. 154.
1895 trilobites; Keith, Reference Keith1895, p. 3.
1933 olenellid trilobites; Reference ResserResser, p. 746.
1936 olenellid trilobites; Reference GrabauGrabau, p. 12.
1949 trilobites; Reference KingKing, p. 520.
1952 Olenellus; Reference King, Hadley and NeumanKing et al., table 2 on p. 4.
1952 Olenellus sp.; Reference King, Hadley and NeumanKing et al., p. 15.
1960 Olenellus; Reference King and FergusonKing and Ferguson, p. 36.
1965 Olenellus; Reference Neuman and NelsonNeuman and Nelson, p. D29.
2016 nevadiid trilobite; Reference Hageman and MillerHageman and Miller, p. 146, fig. 7d.
Diagnosis
Length of genal spine at least one-fifth that of sagittal cephalic length. Posterior margin of glabella drawn out posteriorly into long occipital spine, estimated to be approximately half glabella length (sag.).
Occurrence
Collections ICS-10567 and ICS-10568, upper part of the Murray Shale, locality CM3 (type locality, Appendix), Chilhowee Mountain, Blount County, Tennessee, U.S.A. Also from the Murray Shale in the closely adjacent Little River Gap area (Walcott, 1890, Reference Walcott1891; Appendix). These occurrences are provisionally assigned to the mid- to upper Montezuman Stage, Waucoban Series, traditional “lower” Cambrian of Laurentia (see above), which is likely to fall within provisional Stage 3, Series 2 of the developing global chronostratigraphic zonation of the Cambrian System (Peng et al., Reference Peng, Babcock and Cooper2012).
Description
Cephalon semicircular in outline; proximal portion of posterior cephalic margin oriented very slightly posteriorly when traced distally, distal portion flexed anteriorly by ~20° relative to proximal portion at rounded adgenal angle located less than half of distance from axial furrow to base of genal spine. Greatest observed cephalic length estimated to be ~18.8 mm (sag.). Genal spine broad-based, inner margin of spine smoothly arcs into distal portion of posterior cephalic margin, base of spine transversely opposite posterior portion of lateral or posterior margin of LO; length unknown, but at least one-fifth cephalic length (sag.; Fig. 4.2–4.6, 4.8). Intergenal spine absent or reduced to small dorsal swelling on posterior cephalic border immediately distal to adgenal angle. Cephalic border of low dorsal convexity, poorly defined around entire cephalon by very shallow border furrow; width of anterior border opposite junction of ocular lobes with LA estimated to be slightly less than length (exsag.) of LO. Glabella bullet-shaped in outline, generally tapering anteriorly; ~74−83% of cephalic length (sag.), preglabellar field short (sagittal length approximately equal to or slightly more than that of anterior cephalic border). Maximum width of LA ~87% basal glabellar width (tr.). Posterior margin of glabella strongly convex posteriorly, drawn out posteriorly into long, broad-based occipital spine; length of occipital spine unknown, but estimated to be approximately half glabella length (sag.; Fig. 4.7). All glabellar furrows extremely shallow. SO barely incised over axis, deepest midway between sagittal line and axial furrow, extremely shallow or not incised adjacent to axial furrow, abaxial end slightly anterior to adaxial end. LO subtrapezoidal, slightly widens anteriorly, lateral margins bow outward slightly; more-or-less confluent with L1 anterodistally, ~15–20% glabellar length (sag., excluding occipital spine). S1, S2, and S3 barely visible, shallower than SO, clearest abaxially, absent over axis. S1 approximately parallel to SO; S2 approximately transversely oriented; S3 oriented slightly posterolaterally when followed distally. L1 subtrapezoidal, slightly narrows anteriorly; length (exsag.) ~20% glabellar length (sag., excluding occipital spine). L2 subtrapezoidal, slightly narrows anteriorly; length (exsag.) ~15% glabellar length (sag.). L3 subquadrate to subtrapezoidal, slightly widens anteriorly; length (exsag.) ~10% glabellar length (sag.). Axial furrow shallow at lateral margins of LO and L1, shallows anteriorly, absent around anterior margin of LA. LA slightly wider (tr.) than long (sag.), separated from extraocular area by a subtle break in slope, weakly convex dorsally; widest point at intersection with inner margin of ocular lobes. Weak parafrontal band extends around lateral and anterior margins of LA; anteriorly confluent with extremely weakly defined, broad plectrum; posteriorly merges with outer margin of ocular lobe. Each ocular lobe diverges from exsagittal line at ~42−51° (measured from most abaxial point of outer margin to anterior contact of outer margin with LA) or ~30° (measured from posterior tip to contact of inner margin with glabella), crescentic, flat-topped, posterior tip approximately transversely opposite distal tip of SO or posterior portion of lateral margin of L1; anterior portion more subdued in relief than posterior portion, summit lower than LA and separated from it by break in slope; inner margin poorly defined from interocular area; ocular furrow not apparent. Interocular area slopes outwards and down away from glabella (subhorizontal on USNM 633932, Fig. 4.1); almost twice as wide (tr.) as ocular lobe and ~75−110% width (tr.) of extraocular area opposite S1 (compare Fig. 4.5, 4.6 to Fig. 4.2). Intergenal ridge and posterior ocular line run posterolaterally behind ocular lobe, converge at intergenal swelling. Fine granulations over entire surface on well-preserved specimens. Terrace lines on cephalic doublure at base of genal spines (Fig. 4.5, 4.6). Hypostome, rostral plate, thorax, and pygidium unknown.
Etymology
Named for the location of its discovery, Chilhowee Mountain, Tennessee.
Materials
The species is known from the holotype plus seven additional specimens: USNM 18446 (external mold; Fig. 4.8), USNM 633932 (internal mold; Fig. 4.1), USNM 645832 (part and counterpart; Fig. 4.2), USNM 645833 (part and counterpart; Fig. 4.5, 4.6), USNM 645834 (part and counterpart; Fig. 4.7), USNM 645835 (external mold), USNM 645836 (counterpart).
Remarks
Specimens of Buenellus chilhoweensis n. sp. are preserved as internal and external molds in shale. On some specimens, key morphological features such as the occipital spine are better exhibited on the external mold. Latex peels of the external molds were not made due to the friable nature of the shale: damage to the already very limited number of specimens available was deemed too likely to occur. Instead, internal and external molds of those specimens are figured herein.
Buenellus chilhoweensis n. sp. is very similar to Buenellus higginsi. The most striking differences are in the length of the genal spine (longer in Buenellus chilhoweensis n. sp. than in Buenellus higginsi) and in the size of the axial structure on the occipital ring (a long, prominent spine in Buenellus chilhoweensis n. sp. versus a much smaller spine or node in Buenellus higginsi). No obvious, consistent interspecific differences in other aspects of cephalic shape were observed. Quantitative exploration for any subtle interspecific difference in shape is rendered futile for several reasons. First, the general effacement of the cephalon of both species makes many morphometric variables hard to identify and consistently measure (e.g., width of the cephalic border, or dimensions of particular glabellar lobes). Second, the available sample size is cripplingly low for Buenellus chilhoweensis n. sp., so that the ability to discern statistical significance for any subtle differences between the species is greatly compromised. Finally, both taxa are only known from moldic specimens preserved in shale, and all specimens exhibit some degree of breakage or deformation induced by taphonomic compaction. This not only further reduces sample size for quantitative analysis (because not all variables can be reliably measured on all specimens), but also complicates the interpretation of any such analyses. Compaction-related deformation is known to inflate the degree of variation seen in fossils (see Webster and Hughes, Reference Webster and Hughes1999 and Webster, Reference Webster2015 for quantitative analyses of the effects of compaction on cephalic shape in olenelloid trilobites, and see Babcock and Peel, Reference Babcock and Peel2007 for a discussion of compaction-related variation in Buenellus higginsi). The high degree of variation in proportional width of the extraocular area in Buenellus chilhoweensis n. sp. (compare Fig. 4.5, 4.6 to Fig. 4.2) probably relates to different collapse patterns in response to compaction of an originally strongly convex extraocular area (similar to that seen in non-compacted specimens of Pseudojudomia egregia).
The differences in genal spine size and form of the axial structure on the occipital ring are sufficiently marked that they are robust against such taphonomic issues and thus provide a defensible means for diagnosing the Appalachian form as a distinct species. These differences are interpreted as interspecific rather than ontogenetic in nature because they are expressed in comparably sized specimens (sagittal cephalic lengths range from ~10.7 mm to ~18.8 mm in the sample of Buenellus chilhoweensis n. sp., and from ~5.6 mm to ~24.1 mm in the studied sample of Buenellus higginsi).
Discussion
Buenellus chilhoweensis n. sp. is the oldest known trilobite from the Iapetan margin of Laurentia. Occurring within the Murray Shale, Buenellus chilhoweensis n. sp. is older than the trilobites found in the uppermost Chilhowee Group (Helenmode/Antietam Formation) of the southern and central Appalachians (see above; Fig. 2.2). The oldest reported trilobites in the northern Appalachians of the U.S.A. occur in the Cheshire Formation of Vermont, Massachusetts, and New York State (Dwight, Reference Dwight1887; Walcott, Reference Walcott1888; Gordon, Reference Gordon1911; Shaw, Reference Shaw1954; Knopf, Reference Knopf1962; Landing, 2007, Reference Landing2012). The Massachusetts occurrence documented by Walcott (Reference Walcott1888), however, might be sourced from the top of the stratigraphically older Pinnacle Formation, as noted by Landing (2007, Reference Landing2012, and references therein). Landing (2007, Reference Landing2012) also reported an unidentifiable trilobite fragment from the top of the Bomoseen Member of the Nassau Formation in the Taconics of New York State, which is believed to be age-equivalent to the upper Pinnacle Formation. All those specimens were historically identified (sometimes tentatively) as “Olenellus” (Dwight, Reference Dwight1887; Walcott, Reference Walcott1888; Gordon, Reference Gordon1911) and have been taken to infer a Dyeran age (Landing, 2007, Reference Landing2012). The northern Appalachian occurrences and identifications are currently being re-evaluated (Webster and Landing, in preparation), but to date no specimens have been observed that would indicate a Montezuman age. The oldest trilobites known from western Newfoundland and Labrador occur in the basal Forteau Formation (Schuchert and Dunbar, Reference Schuchert and Dunbar1934; North, Reference North1971; Stouge and Boyce, Reference Stouge and Boyce1983), for which a mid-Dyeran age is well established (Palmer and James, Reference Palmer and James1979; Debrenne and James, Reference Debrenne and James1981; James et al., Reference James, Stevens, Barnes and Knight1989). The oldest trilobites reported from eastern Greenland occur in the Bastion Formation and are also of mid-Dyeran age (Poulsen, Reference Poulsen1932; Cowie, Reference Cowie1971; Skovsted, Reference Skovsted2006; Stein, Reference Stein2008).
The apparent absence of Montezuman trilobites from the Iapetan margin of Laurentia was curious, given that trilobites of that age have been reported from the adjacent Innuitian margin (Buenellus higginsi, see above) and that Montezuman-age trilobites—including fallotaspidoids, which occur in lowermost Montezuman strata—are diverse and abundant along the Cordilleran margin (e.g., Fritz, 1972, Reference Fritz1973; Nelson, 1976, Reference Nelson1978; Hollingsworth, 2005, 2007, Reference Hollingsworth2011). Was there some paleobiogeographic reason why trilobites did not invade the Iapetan margin until much later? Discovery of Buenellus chilhoweensis n. sp. resolves that dilemma: trilobites did inhabit the Iapetan margin, at least locally, during the Montezuman. Trilobites in rocks of this age are clearly difficult to find in the Appalachians, but the Chilhowee Mountain discovery demonstrates their presence and encourages further effort in the field to more fully document their early history there.
Detailed correlation of lower Cambrian strata between the Iapetan and Cordilleran margins of Laurentia has proven difficult. The difficulty arises in part from the absence of a detailed and widely applicable biostratigraphic zonation of the Iapetan strata (although a local zonation scheme has been developed for the upper Dyeran of eastern New York State; Bird and Rasetti, Reference Bird and Rasetti1968). Recent and ongoing fieldwork in the Appalachians is yielding new fossils that can promote the development of an Iapetan margin biostratigraphy and assist in circum-continental correlation (e.g., Hageman and Miller, Reference Hageman and Miller2016; Webster and Landing, Reference Webster and Landing2016; this study). Sequence stratigraphic data might also prove useful in this endeavor. For example, attempts have been (and continue to be) made to correlate the late Dyeran “Hawke Bay Regression” around Laurentia and further afield (e.g., Palmer and James, Reference Palmer and James1979; Palmer, Reference Palmer1981; Landing et al., 2002, Reference Landing, Geyer and Heldmaier2006; Bordonaro, Reference Bordonaro2003; Landing, Reference Landing2012; Geyer and Vincent, Reference Geyer and Vincent2015; Webster and Landing, Reference Webster and Landing2016). Sequence stratigraphic interpretations and sometimes sea level curves are being developed for Montezuman and lower Dyeran strata of both the Iapetan (Whisonant, Reference Whisonant1974; Mack, Reference Mack1980; James et al., Reference James, Stevens, Barnes and Knight1989; Barnaby and Read, Reference Barnaby and Read1990; Lavoie et al., Reference Lavoie, Burden and Lebel2003; Landing, 2007, Reference Landing2012; Tull et al., Reference Tull, Allison, Whiting and John2010; see above) and Cordilleran margins (Hollingsworth 2005, 2007, Reference Hollingsworth2011; Dilliard et al., 2007, Reference Dilliard, Pope, Coniglio, Hasiotis and Lieberman2010; English and Babcock, Reference English and Babcock2010; Webster and Bohach, Reference Webster and Bohach2014). Discovery of Buenellus chilhoweensis n. sp. within the upper part of the Murray Shale provides a valuable biostratigraphic calibration for the depositional sequences recognized along the Iapetan margin. With this and future fossil discoveries in the Chilhowee Group, it might ultimately become possible to recognize time-equivalent sedimentary packages around Laurentia and thus add sequence stratigraphy to the arsenal of tools for high-resolution circum-continental correlation of lower Cambrian rocks.
Finally, the occurrence of Buenellus chilhoweensis n. sp. and Isoxys chilhoweanus in the Murray Shale, and of Buenellus higginsi and Isoxys volucris Williams, Siveter, and Peel, Reference Williams, Siveter and Peel1996 in the Sirius Passet Lagerstätte, is noteworthy. Buenellus is known only from those two units and therefore is either poorly sampled or appears to have been rather limited in its environmental tolerance. Isoxys was an arthropod genus with a wide geographic and stratigraphic range, but it possessed a very thin, probably non-mineralized, shield with low preservation potential, and thus is only preserved in Konservat-Lagerstätten (Williams et al., Reference Williams, Siveter and Peel1996; Vannier and Chen, Reference Vannier and Chen2000; Stein et al., Reference Stein, Peel, Siveter and Williams2010). Does the combined occurrence of Buenellus and Isoxys, in sediments that accumulated within a deep water, low energy, outer shelf paleoenvironment, indicate that the Murray Shale has the potential to yield a soft-bodied fauna analogous to the Sirius Passet Lagerstätte? The possibility is intriguing, and provides another reason for further fieldwork in the Appalachians.
Acknowledgments
We thank M. Smith, R. Benfield, and students from Appalachian State University for assistance in the field. The first trilobite specimen from Chilhowee Mountain found as part of the present study was discovered by M. Smith accompanied by SJH. Blackberry Farm generously provided access to the locality at which Buenellus chilhoweensis n. sp. was found and donated multiple specimens to the USNM. Accessioning of specimens to the USNM was facilitated by D. Levin. Helpful comments were provided by reviewers J.S. Peel and P. Ahlberg, and associate editor B. Pratt. This work was funded in part by NSF Research Grant EAR Integrated Earth Systems 1410503 to MW.
Appendix: Localities
Little River Gap area, Walland, TN
Original locality of Walcott and Keith.—Walcott (Reference Walcott1890) and Keith (Reference Keith1895) reported finding fossils on the east side of Little River Gap, which is a river gorge cut through Chilhowee Mountain near the town of Walland (Fig. 3.1). This site was subsequently referred to as “USNM Locality 17” by Resser (Reference Resser1938; the Little River Gap locality was also mentioned by Resser [1933, p. 746], but was erroneously stated to be in Virginia). The fauna definitely known from USNM Locality 17 consists of Buenellus chilhoweensis n. sp. (Fig. 4.8), the arthropod Isoxys chilhoweanus, the bradoriid Indota tennesseensis, and the hyolith figured by Resser (Reference Resser1938, pl. 4, fig. 31). It is also possible that the brachiopods mentioned by Keith (Reference Keith1895) were sourced from this locality, although some of or all those specimens might have been collected from the Murray Gap area.
The precise location of the original fossil-bearing site at Little River Gap is not known. Labels on the specimens denote only that the locality was at “the east end of the Little River Gap, Chilhowee Mountain, Tennessee” (Neuman and Nelson, Reference Neuman and Nelson1965, p. D29). This vagueness, combined with the failure of subsequent investigators to discover additional fossils in the area (e.g., King et al., Reference King, Hadley and Neuman1952, p. 15), also led to uncertainty over the stratigraphic provenance of the original fossils. Although the fossils were stated to have been collected from the Murray Shale (Keith, Reference Keith1895), King et al. (Reference King, Hadley and Neuman1952) described the roadside section at Little River Gap and noted that the Murray Shale has been cut out of the section by faulting. They (King et al., Reference King, Hadley and Neuman1952, p. 15, 17, table 5) concluded that, at Little River Gap, the fossil-bearing “Murray Shale” of Keith (Reference Keith1895) is actually the Helenmode Formation. The same conclusion was reached by King and Ferguson (Reference King and Ferguson1960) and by Neuman and Nelson (Reference Neuman and Nelson1965, p. D28, D29).
The classic Little River Gap roadcut (Fig. 3.1, Locality LRG; GPS coordinates 35°43.914’N, 083°48.936’W) was examined by the present authors in 2016. Our observations are congruent with the geologic map presented by King et al. (Reference King, Hadley and Neuman1952, fig. 5). The outcrop on the northeast side of the old road (on the east side of Little River) comprises an intermittently exposed stratigraphic section from the Cochran Formation to the Nebo Quartzite. The Hesse Quartzite is perhaps also exposed at the east end of the outcrop, but interpretation of the stratigraphy is complicated by a fault and by soil cover. The Nebo Quartzite contains several shale intervals, each about 20−30 cm thick, that resemble the lithology of the Murray Shale, but the Murray Shale itself appears to be absent from the roadcut. Trace fossils occur in thin-bedded shale-siltstone and fine sandstone beds of the Nebo Member, but no body fossils were observed. Given that all other occurrences of Buenellus chilhoweensis n. sp. and Isoxys chilhoweanus are in the Murray Shale (herein; Walcott, Reference Walcott1890), we conclude that the fossils assigned to USNM Locality 17 were sourced from the Murray Shale at a site on Chilhowee Mountain close to (but not at) the roadside exposure on the east side of Little River Gap.
New Localities on Chilhowee Mountain.—Several fossil-bearing exposures in the Chilhowee Group were discovered by SJH on Chilhowee Mountain to the northeast of the classic Little River Gap roadcut. This series of exposures is on privately owned land, maintained as the Three Sisters Conservation Area (managed by Blackberry Farm). We stress that access to the localities requires expressed permission from the landowners: the locality information presented here is published with their approval. The exposures are in the banks of an unmarked jeep trail that winds through several switchbacks to reach the summit ridgeline of Chilhowee Mountain (Fig. 3.1). The trail crosses over much of the Chilhowee Group stratigraphy, from the Cochran Formation (at the foot of the trail) to the Hesse Quartzite (forming the summit ridgeline).
The stratigraphically lowest exposure of interest is in the Nichols Shale (Fig. 3.1, Locality CM1; GPS coordinates 35°44.649’N, 083°48.892’W). Several person-hours of collecting and splitting of ~100 kg of bulk samples from the shale to siltstone and fine-grained sandstone at this site yielded several trace fossils. The absence of macroscopic body fossils despite the suitable lithology for their preservation is consistent with assignment of the Nichols Shale to a pre-trilobite age (Begadean Series), in accord with previous studies (Hageman and Miller, Reference Hageman and Miller2016). This provisional age assignment could be tested with further collecting effort within this stratigraphic unit, particularly if acritarchs could be extracted.
Further up the mountainside lie exposures of the ridge- and cliff-forming Nebo Quartzite. Cross-bedding and Skolithos burrowing were observed within that unit, but no serious effort to look for body fossils was made due to the discouraging lithofacies. Higher still, several meters of shale within the lowermost part of the Murray Shale are exposed in close juxtaposition to the top ledge of the Nebo Quartzite (Fig. 3.1, Locality CM2; GPS coordinates 35°44.817’N, 083°48.446’W). That shale yielded bradoriids, indeterminate carbonaceous filaments, and abundant trace fossils. The bradoriid specimens have yet to be identified, but their stratigraphic position within the lower few meters of the Murray Shale is consistent with the occurrence of Indotes tennesseensis at Murray Gap (Laurence and Palmer, Reference Laurence and Palmer1963; Wood and Clendening, Reference Wood and Clendening1982).
Seven specimens of Buenellus chilhoweensis n. sp. and abundant hyoliths were recovered from an ~0.5 m thick interval within the upper portion of the Murray Shale at an exposure located further east and up the hillside (Fig. 3.1, Locality CM3; GPS coordinates 35°45.076’N, 083°48.030’W). Cliffs of the cross-bedded, Skolithos-bearing Hesse Quartzite form the ridgeline summit above this locality. The exact stratigraphic distance of the trilobite-bearing interval below the base of the Hesse Quartzite cannot be measured due to soil cover, but hillside topography suggests that the distance is on the order of 10−20 m. The lithology of the trilobite-bearing interval is a friable shale and siltstone that weathers into chips.
Murray Gap Area
Original locality of Walcott and Keith.—In his description of Isoxys chilhoweanus, Walcott (Reference Walcott1890, p. 626) reported that some of the fossils were sourced from “near Montvale Springs” on Chilhowee Mountain. Keith (Reference Keith1895, p. 3) also noted “the crest of the mountain above Montvale Springs” as a source for fossils from the Murray Shale. This indicates that the fossils were collected from the Murray Gap area. Unfortunately, the exact location of the original fossil-bearing site in the Murray Gap area cannot be determined from the available documentation. Comparison of modern and historical maps reveals that the present-day position of roads (and thus roadcuts) is not precisely congruent with the distribution of trails that would have provided access to the ridge crest in the late 19th Century. During reconnaissance of the area, SJH discovered an exposure of the Murray Shale alongside an old, disused bridleway (Fig. 3.2, Locality MG1; GPS coordinates 35°37.732'N, 083°57.266'W) ~700 m west of the present-day road junction at Murray Gap. Much of the lower third of the Murray Shale, including the basal contact with the underlying Nebo Quartzite, is exposed. It is conceivable, but not certain, that the original collection mentioned by Walcott (Reference Walcott1890) and Keith (Reference Keith1895) was sourced from this outcrop because the present-day Happy Valley Road did not exist at that time (Keith, Reference Keith1895 topographic map). No fossils were observed at the site during a brief visit in 2016.
Murray Gap Roadcuts.—Laurence and Palmer (Reference Laurence and Palmer1963) and Wood and Clendening (Reference Wood and Clendening1982) described fossils collected from the Murray Shale at a series of roadcuts that were excavated in 1962. One of these roadcuts, on Happy Valley Road (Fig. 3.2, Locality MG2; GPS coordinates 35°37.845'N, 083°56.911'W), was found during reconnaissance by the authors in December 2016 to be mostly obscured by soil and leaves. A second, much larger, roadcut alongside the Foothills Parkway (Fig. 3.2, Locality MG3; GPS coordinates 35°37.897'N, 083°56.678'W) exposes a spectacular section through much of the Murray Shale, and offers the potential for detailed paleontological and sedimentological study. Both these roadcuts are located within the bounds of the Foothills Parkway National Park, and a permit is required to conduct work at either site.
A roadside cliff at the intersection of Happy Valley Road and Flats Road (Fig. 3.2, Locality MG4; GPS coordinates 35°37.781'N, 083°56.552'W) exposes ~30 m of the Murray Shale. To our knowledge, this site has not been mentioned by previous workers. Numerous hyolith and bradoriid specimens and abundant trace fossils were observed at this site during reconnaissance by the authors in December 2016. The position of this fossiliferous interval within the Murray Shale cannot be directly measured because formational contacts are not exposed at this locality. However, if the as-yet-unidentified bradoriid is Indota tennesseensis, then a position within the lower part of the Murray Shale could be hypothesized based on constrained stratigraphic occurrences of that taxon in the nearby roadcuts (Laurence and Palmer, Reference Laurence and Palmer1963; Wood and Clendening, Reference Wood and Clendening1982).





 Open access
Open access