



Introduction
Lower Silurian (Llandovery) marine sediments are distributed along a NNE-SSW trend on the South China plate, which was largely occupied by the Yangtze region in the northwestern sector. During the Llandovery, the Yangtze region has been interpreted to represent a tropical or subtropical ‘epeiric platform’ in western Gondwana (e.g., Rong and Cocks, Reference Rong and Cocks2014). In the earliest Silurian (Rhuddanian stage), black shales with flysch are dominant sediments in the Yangtze region. Due to the rapid rise of sea level, late Aeronian limestone facies became widespread in the epeiric sea of the Yangtze region, however, the early Telychian is dominated by terrigenous sediments with minor calcareous interbeds due to the expansion of the Yangtze Uplift (Rong et al., Reference Rong, Chen, Su, Ni, Zhan, Chen, Fu, Li, Fan, Landing and Johnson2003, Reference Rong, Wang and Zhang2012; Deng et al., Reference Deng, Wang and Li2012). The early Telychian rocks are represented by the Hanchiatien (or Majiaochong) Formation, usually over a kilometer in thickness, spanning from northern Guizhou to southern Sichuan provinces and the southern Chongqing District. Li et al. (Reference Li, Ernst, Munnecke, Yu, Li and Deng2018) provided an in-depth study of the meter-sized, bryozoan-bearing bioherms, described preliminarily earlier in Chinese by Li et al. (Reference Li, Wang, Li, Ma, Zhang, Deng and Cai2012). The bryozoan fauna in those bioherms have not yet been given a systematic treatment. Previous studies of the early Telychian bryozoan fauna of the Yangtze region are scarce (Hu, Reference Hu1982; Xia and Qi, Reference Xia and Qi1989), thus, a detailed description of the Hanchiatien bryozoans can provide valuable insights into the faunal composition in the region.
Telychian bryozoans from South China were previously described in several papers. Hu (Reference Hu1982) identified six bryozoan species from the Cuijiagou Formation (lower Telychian) of Guangyuan, Sichuan: Fistulipora guangyuanensis Hu, Reference Hu1982, Fistuliramus eximius Hu, Reference Hu1982, Hennigopora multilamellosa Hu, Reference Hu1982, Hennigopora sp. indet., Trematopora sinensis Hu, Reference Hu1982, and Trematopora sp. indet. From the upper Telychian Ningqiang Formation of Ningqiang (Shanxi), he identified 14 species (Hu, Reference Hu1982, Reference Hu1990): Calloporella silurica Hu, Reference Hu1982, Cyclotrypa solidoscens Hu, Reference Hu1990, Cyphotrypa undulata Hu, Reference Hu1990, Fistulipora ningqiangensis Hu, Reference Hu1990, Fistuliramus transversus Hu, Reference Hu1982, Hennigopora petaliformis Hu, Reference Hu1982, Hallopora aggregata Hu, Reference Hu1982, Homotrypa ningqiangensis Hu, Reference Hu1990, Leioclema speciosum Hu, Reference Hu1982, Leioclema sp. indet., Monotrypa shaanxiensis Hu, Reference Hu1990, Orbignyella curvata Hu, Reference Hu1982, O. globata Yang, Reference Yang1951, and O. mui Yang, Reference Yang1951.
Xia and Qi (Reference Xia and Qi1989) described four species from the lower Telychian Chenxiacun Formation of Hanshan (Anhui): Hallopora elegantula (Hall, Reference Hall1852), Hallopora hanshanensis Xia and Qi, Reference Xia and Qi1989, Hallopora raritabulata Xia and Qi, Reference Xia and Qi1989, and Trematopora reflua Xia and Qi, Reference Xia and Qi1989.
Geological settings and depositional environment
The Hanchiatien Shale was originally defined by Ting (Reference Ting1930) and was subsequently treated as the Hanchiatien Group (e.g., Yin, Reference Yin1949; Mu, Reference Mu1962). Since 1978, it has been formally named the Hanchiatien Formation (e.g., Southwest Institute of Geologic Sciences, 1978; Rong et al., Reference Rong, Chen, Wang, Geng, Wu, Deng, Chen and Xu1990), characterized by brownish to greenish shales. The shales are usually disconformably underlain by the upper Aeronian Shihniulan Formation and disconformably capped by the lower Permian Liangshan or Tongkuangxi formations (Zhan and Jin, Reference Zhan and Jin2007). In most cases, there is a unit (normally < 10 m thick) of purplish red shales known as the Lower Red Beds at the base of the Hanchiatien Formation. Stratigraphically, the Lower Red Beds are a well-known marker unit in the Yangtze region (Rong et al., Reference Rong, Wang and Zhang2012).
Although the Hanchiatien Formation is poorly fossiliferous, diagnostic chitinozoans and graptolites were reported at the top of the formation in the Northeast Guizhou: Ancyrochitina brevicollis Geng, Reference Geng1986 and Streptograptus plumosus Baily, Reference Baily, Traill and Egan1871 suggest the formation should be correlated to early Telychian (Chen, Reference Chen1986; Geng et al., Reference Geng, Qian, Ding, Wang, Wang and Cai1997). Some carbonate interbeds (or marls) occur in the Hanchiatien Formation in a few localities (e.g., Li et al., Reference Li, Wang, Li, Ma, Zhang, Deng and Cai2012). Macrofossils (e.g., brachiopods, trilobites, echinoderms, bryozoans, and corals) are abundant in some of the carbonate interbeds. To date, no systematic study has been done on the bryozoans from this formation. Here, we provide the first taxonomic description of the bryozoans from the lower Hanchiatien Formation at the border of Guizhou Province.
The Hanchiatien Formation is thought to have been deposited in a subtidal setting, mainly based on the distributional pattern of the brachiopods (Rong et al., Reference Rong, Chen, Su, Ni, Zhan, Chen, Fu, Li, Fan, Landing and Johnson2003). However, different facies of the Hanchiatien Formation display a fluctuation of marine environments (Li et al., Reference Li, Ernst, Munnecke, Yu, Li and Deng2018). The paleogeographical setting of the Hanchiatien Formation indicates a seaward ramp oriented northward. Depositional environments could have varied from place to place in the Yangtze epeiric sea.
The Jiebei section is located at the boundary between Chongqing and Guizhou provinces (Fig. 1). The stratigraphic details were given by Li et al. (Reference Li, Ernst, Munnecke, Yu, Li and Deng2018) who described the lower Hanchiatien Formation as brownish to greenish shales with minor silstones, which cap the Shihniulan Formation disconformably. In this section, there are two bioherms 85–120 m above the base of the Hanchiatien Formation. At the bottom of the second bioherm, there is a layer (~1.7 m thick) of cross-bedded, oolitic grainstone with ripples (Fig. 2). Lacking any photic-related fossils, the bryozoan bioherms demonstrate an unusual heterozoan marine community, mainly produced by internal waves (Li et al., Reference Li, Ernst, Munnecke, Yu, Li and Deng2018). Our interpretation is that the lower Hanchiatien Formation in Jiebei represents mid-outer ramp settings.
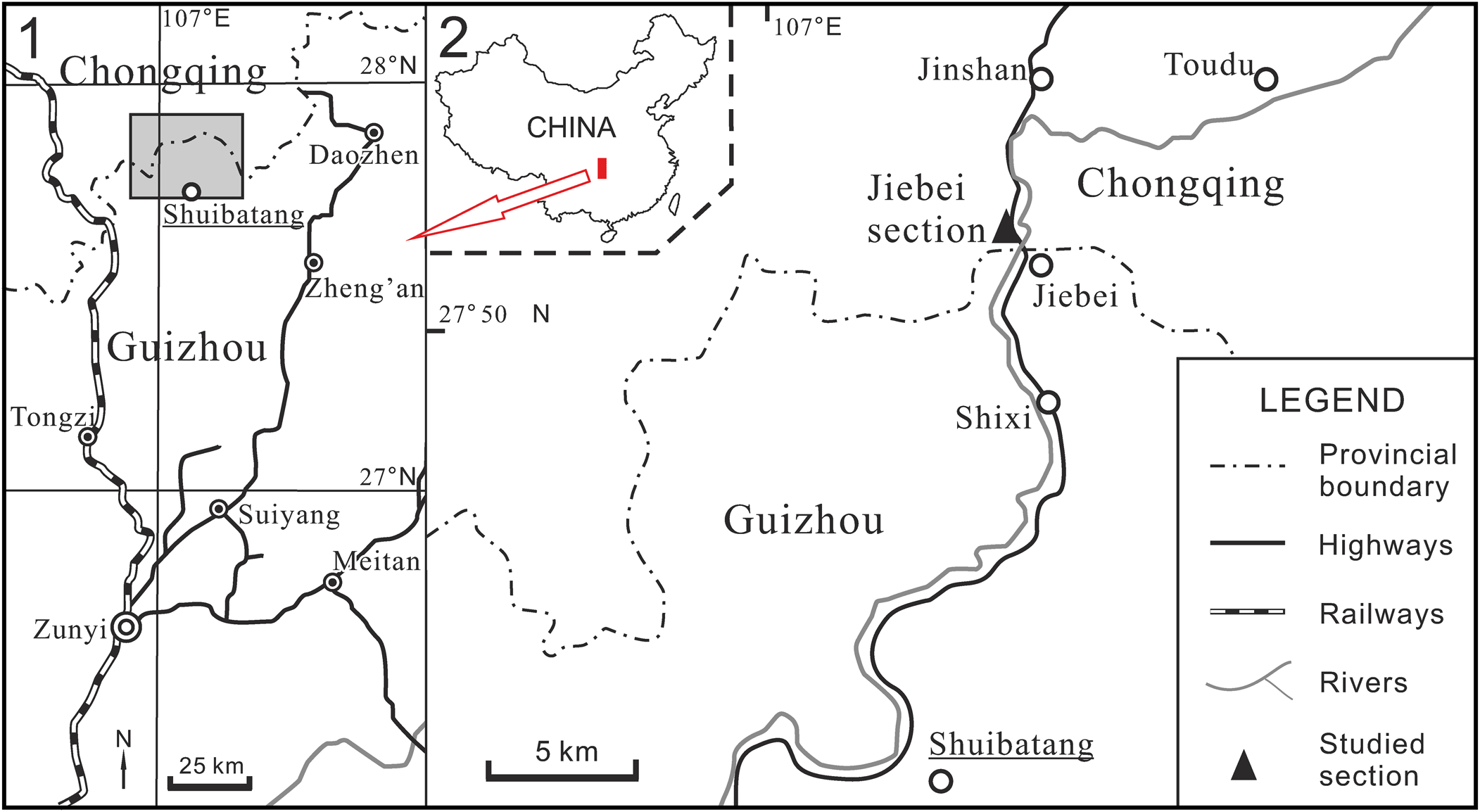
Figure 1. Location of the studied section between Chongqing and Guizhou provinces, South China: (1) region between Chongqing and Guizhou provinces; (2) detail of the shaded-box area in (1), showing the study section.
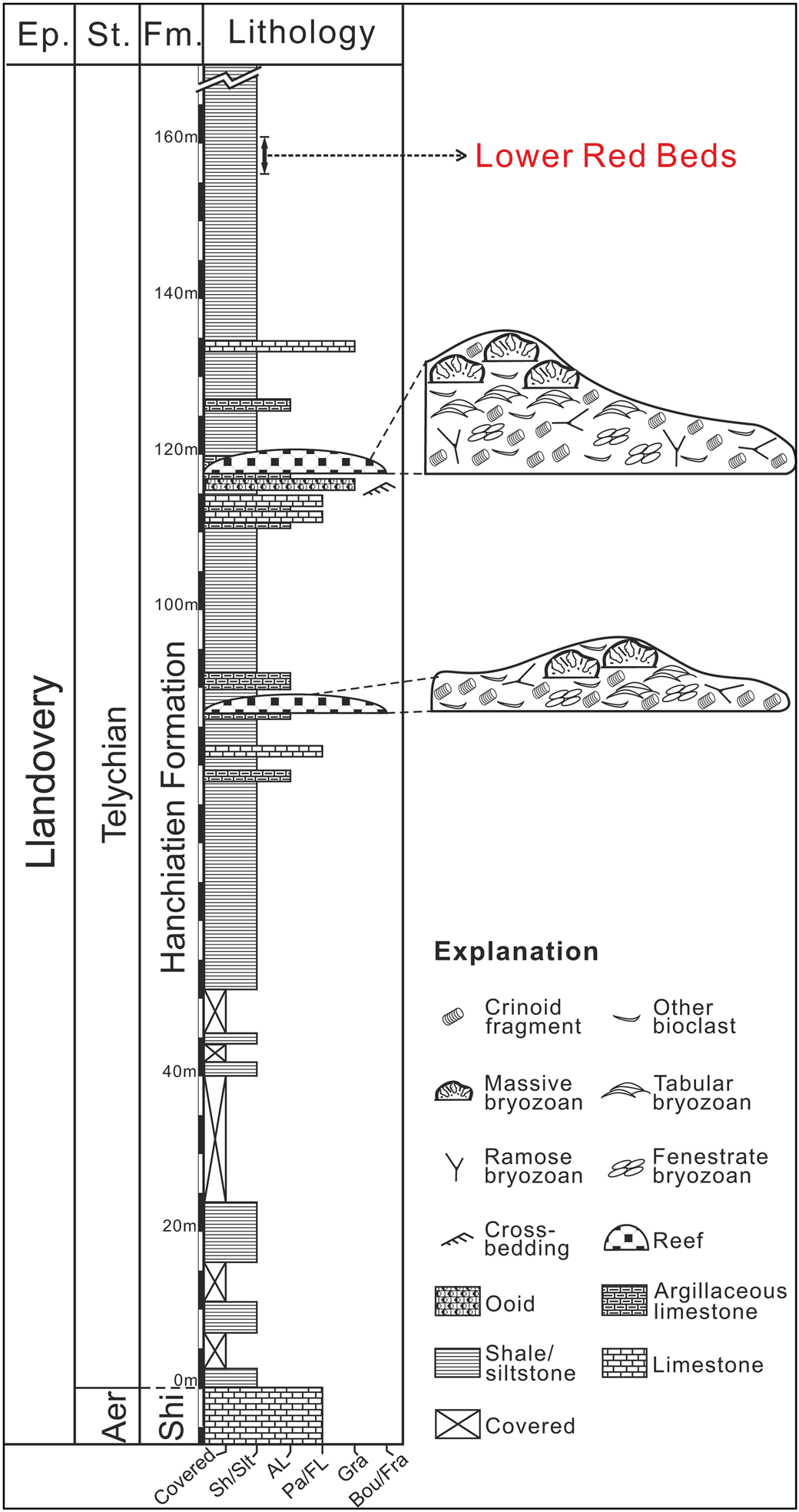
Figure 2. Lithologic column of the Jiebei section (from Li et al., Reference Li, Ernst, Munnecke, Yu, Li and Deng2018). Stratigraphic positions of reefs are indicated. Aer = Aeronian; AL = argillaceous limestone; Bou/Fra = boundstone/framestone; Ep. = epoch; Fm. = formation; Gra = grainstone; Pa/FL = packstone/floatstone; Sh/Slt = shale/siltstone; Shi = Shihniulan Formation; St. = stage.
Material and methods
All bryozoans were collected from the bioherms (Figs. 2, 3) in June 2016 by QJL. Thin sections were made in the laboratory of the GeoZentrum Nordbayern, Friedrich-Alexander University Erlangen-Nuremberg, Germany. Bryozoans were studied in thin sections using a binocular microscope under transmitted light. In total, 42 thin sections were prepared from rock samples. The spacing of structures is measured as the distance between their centers. Statistics were summarized using arithmetic means, sample standard deviations, coefficients of variation, and minimum and maximum values.

Figure 3. Field aspects of the studied section: (1) the flank facies of the second reef horizon, showing the branched bryozoan Trematopora; (2) the core facies of the second reef horizon; (3) plan view of the framestone, showing bulbous and knobby colonies of the first reef horizon; (4) the flank facies of the first reef horizon. Scale bars = 2 m (2); 8 cm (4); 2 cm (3); 1 cm (1).
Based on the genus-level occurrence databases of Silurian bryozoans (Hu, Reference Hu1982, Reference Hu1990; Xia and Qi, Reference Xia and Qi1989; Anstey et al., Reference Anstey, Pachut and Tuckey2003), pair-group cluster analysis (Euclidean similarity index) and detrended correspondence analysis were implemented with the PAST statistical package (version 2.16; Hammer et al., Reference Hammer, Harper and Ryan2001) to assess the paleobiogeographical relationship of the Telychian bryozoan fauna from the South China Block. Both cluster analysis and detrended correspondence analysis make no assumptions about the data structure, so they were suitable for our large-scale paleobiogeographical studies (Shi, Reference Shi1993). To ensure the robustness of our results, taxa that were potentially poorly known were removed from the published presence-absence dataset (see Appendix).
Repositories and institutional abbreviations
The newly studied material is deposited at the Naturmuseum Senckenberg (SMF), Frankfurt am Main, Germany. Other cited repositories are: NIGP = Nanjing Institute of Geology and Paleontology, China.
Systematic paleontology
Phylum Bryozoa Ehrenberg, Reference Ehrenberg1831
Class Stenolaemata Borg, Reference Borg1926
Superorder Palaeostomata Ma, Buttler, and Taylor, Reference Ma, Buttler, Taylor, Rosso, Wyse Jackson and Porter2014
Order Cystoporata Astrova, Reference Astrova1964
Suborder Fistuliporina Astrova, Reference Astrova1964
Family Fistuliporidae Ulrich, Reference Ulrich1882
Genus Fistulipora M‘Coy, Reference M'Coy1849
Type species
Fistulipora minor M‘Coy, Reference M'Coy1849; Carboniferous, England.
Fistulipora guangyuanensis Hu, Reference Hu1982
Figure 4.1–4.5; Table 1
- Reference Hu1982
Fistulipora guangyuanensis Hu, p. 296, pl. 3, figs. 11, 12.
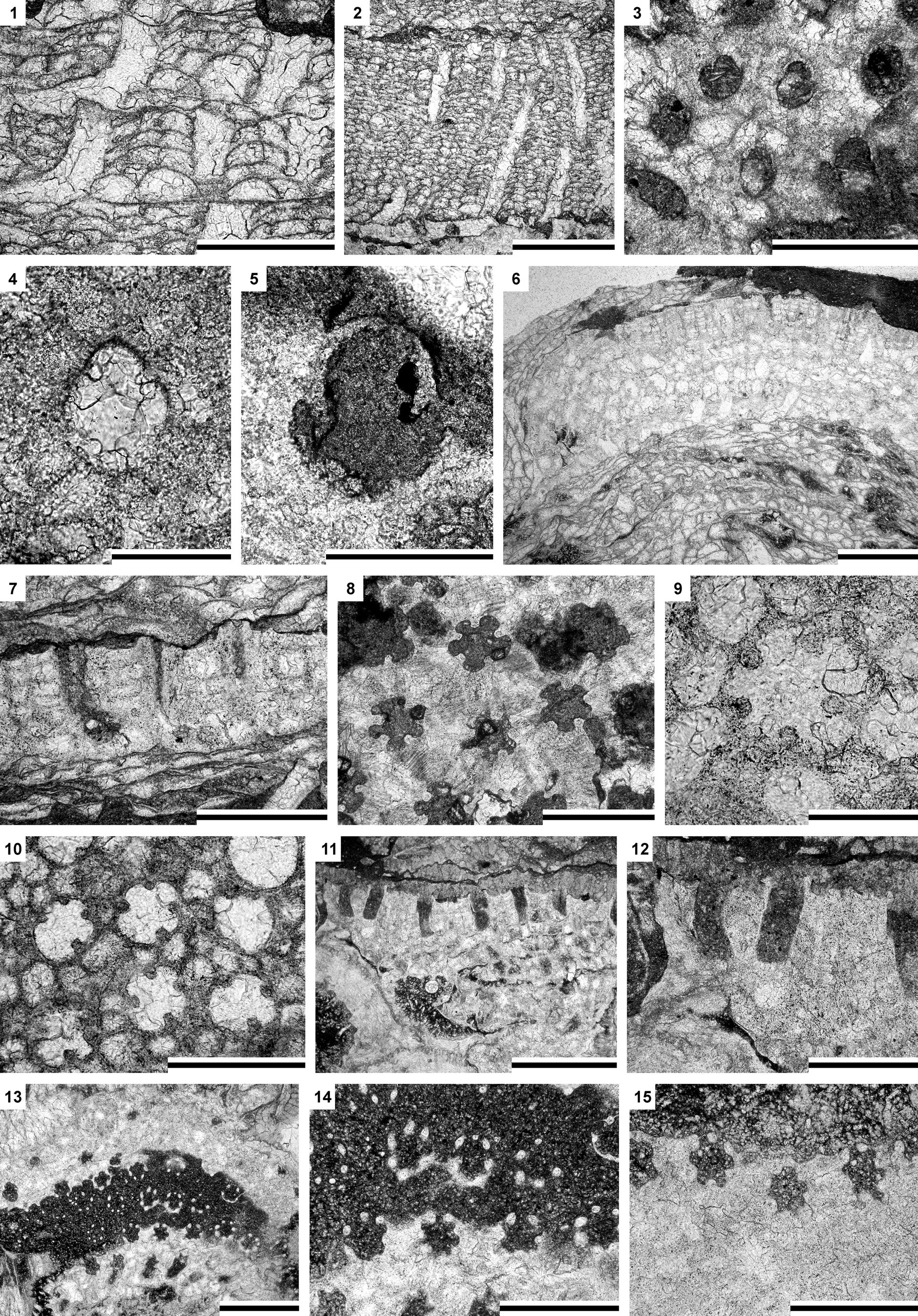
Figure 4. (1–5) Fistulipora guangyuanensis Hu, Reference Hu1982: (1) longitudinal section of a multilayered encrusting colony, SMF 60526; (2) longitudinal section of an encrusting colony, SMF 60526; (3, 4) tangential section showing autozooecial apertures and vesicles, SMF 60525; (5) tangential section showing autozooecial aperture with lunarium, SMF 60524; (6–10) Hennigopora multilamellosa Hu, Reference Hu1982: (6) longitudinal section, SMF 60570; (7) longitudinal section showing autozooecia and vesicles, SMF 60552; (8–10) tangential sections showing autozooecial apertures, acanthostyles, and vesicles: (8) SMF 60544; (9) SMF 60542; (10) SMF 60554; (11–15) Hennigopora sp. indet., SMF 60577: (11, 12) longitudinal section of an encrusting colony; (13–15) tangential sections showing autozooecial apertures, acanthostyles, and vesicles. Scale bars = 1 mm (2, 6, 11, 13); 0.5 mm (1, 3, 7, 8, 10, 12, 14, 15); 0.2 mm (4, 5, 9).
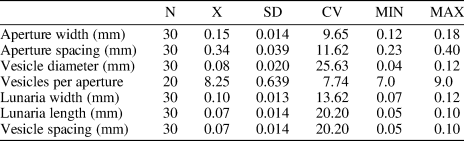
Table 1. Descriptive statistics of Fistulipora guangyuanensis Hu, Reference Hu1982. CV = coefficient of variation; Max = maximum value; Min = minimum value; N = number of measurements; SD = sample standard deviation; X = mean.
Holotype
NIGP 63771; Cuijiagou Formation, Telychian, Llandovery, lower Silurian; Sichuan Province, China.
Occurrence
Jiebei village, Chongqing, China; lower part of the Hanchiatien Formation, lower Telychian, Llandovery, lower Silurian.
Materials
SMF 60500−60538.
Remarks
Fistulipora guangyuanensis differs from Fistulipora ningqiangensis from the lower Silurian of Ningqiang, China in its smaller apertures (aperture width 0.12–0.18 mm vs. 0.17–0.25 mm [Hu, Reference Hu1990], respectively) and larger lunaria. Fistulipora guangyuanensis differs from Fistulipora ternavensis Astrova, Reference Astrova1965 from the lower−middle Silurian of Russia in its smaller autozooecial apertures (aperture width 0.12–0.18 mm vs. 0.18–0.23 mm [Astrova, Reference Astrova1965], respectively), larger lunaria, and less abundant vesicles (7–9 per autozooecial aperture vs. 8–10 vesicles [Astrova, Reference Astrova1965], respectively).
Family Xenotrypidae Utgaard, Reference Utgaard and Robison1983
Hennigopora Bassler, Reference Bassler1952
Type species
Callopora florida Hall, Reference Hall1852; Niagaran Group, Rochester Shale, Sheinwoodian, Wenlock, Silurian; Lockport, New York, USA.
Hennigopora multilamellosa Hu, Reference Hu1982
Figure 4.6–4.10; Table 2
- Reference Hu1982
Hennigopora multilamellosa Hu, p. 295, pl. 2, figs. 4, 5.
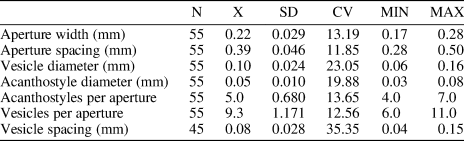
Table 2. Descriptive statistics of Hennigopora multilamellosa Hu, Reference Hu1982. Abbreviations as for Table 1.
Holotype
NIGP 63765; Cuijiagou Formation, Telychian, Llandovery, lower Silurian; Sichuan Province, China.
Occurrence
Jiebei village, Chongqing, China; lower part of the Hanchiatien Formation, lower Telychian, Llandovery, lower Silurian.
Materials
SMF 60539−60576.
Remarks
Hennigopora multilamellosa differs from Hennigopora florida (Hall, Reference Hall1852) from the middle Silurian of New York, USA in its smaller autozooecial apertures (aperture width 0.17−0.28 mm vs. 0.28−0.35 mm [Bassler, Reference Bassler1906], respectively). Hennigopora multilamellosa differs from Hennigopora hunanensis Yang and Xia, Reference Yang and Xia1974 from the lower Silurian of China in having smaller autozooecial apertures (aperture width 0.17−0.28 mm vs. 0.20−0.30 mm [Yang and Xia, Reference Yang and Xia1974], respectively) and in its less abundant vesicles (6−11 per aperture vs. 8−13, respectively). Hennigopora multilamellosa differs from Hennigopora apta Perry and Hattin, Reference Perry and Hattin1960 from the lower Silurian of Indiana, USA in the presence of 4−7 acanthostyles per autozooecial aperture instead of 3−5 (Ernst et al., Reference Ernst, Brett and Wilson2019) in the latter species. Hennigopora multilamellosa differs from Hennigopora petaliformis from the Ningqiang Formation (upper Telychian) of Shanxi in its encrusting instead of branched erect colony and in the presence of 4−7 acanthostyles per autozooecial aperture instead of 2−4 (Hu, Reference Hu1982) in the latter species.
Hennigopora sp. indet.
Figure 4.11–4.15; Table 3
Occurrence
Jiebei village, Chongqing, China; lower part of the Hanchiatien Formation, lower Telychian, Llandovery, lower Silurian.
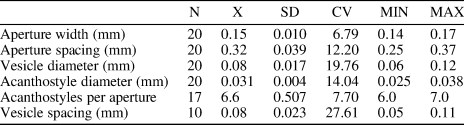
Table 3. Descriptive statistics of Hennigopora sp. indet. Abbreviations as for Table 1.
Description
Encrusting colony, 0.40−1.25 mm thick. Autozooecia originating from thin epitheca, bending in the early exozone to the colony surface, with rounded-polygonal, often petaloid, apertures due to indenting acanthostyles. Basal diaphragms rare, straight, thin. Vesicles generally large, separating autozooecia in one or two rows, 8−10 surrounding each autozooecial aperture, with rounded roofs, polygonal in tangential section. Acanthostyles abundant, relatively large, often deeply indenting autozooecial chambers, six or seven surrounding each autozooecial aperture. Autozooecial walls displaying obscure granular microstructure, 0.005−0.010 mm thick. Maculae not observed.
Materials
Single specimen, SMF 60577.
Remarks
Hennigopora sp. indet. differs from Hennigopora multilamellosa in its smaller autozooecial apertures (mean aperture width 0.15 mm vs. 0.22 mm [Table 2], respectively) and more abundant acanthostyles (mean 6.6 acanthostyles per autozooecial aperture vs. 5 [Table 2], respectively). Hennigopora sp. indet. differs from Hennigopora apta from the lower Silurian of Indiana, USA in the presence of six or seven acanthostyles per autozooecial aperture instead of 3−5 (Ernst et al., Reference Ernst, Brett and Wilson2019) in the latter species.
Order Trepostomata Ulrich, Reference Ulrich1882
Suborder Halloporina Astrova, Reference Astrova1965
Family Heterotrypidae Ulrich, Reference Ulrich1890
Genus Leioclema Ulrich, Reference Ulrich1882
[= Lioclema Ulrich, Reference Ulrich1882]
Type species
Callopora punctata Hall, Reference Hall, Hall and Whitney1858; lower Carboniferous, Iowa, USA.
Leioclema speciosum Hu, Reference Hu1982
Figure 5.1–5.6; Table 4
- Reference Hu1982
Leioclema speciosum Hu, p. 295, pl. 2, figs. 9−13.
- Reference Hu1990
Leioclema speciosum; Hu, p. 29, pl. 2, figs. 4−6.
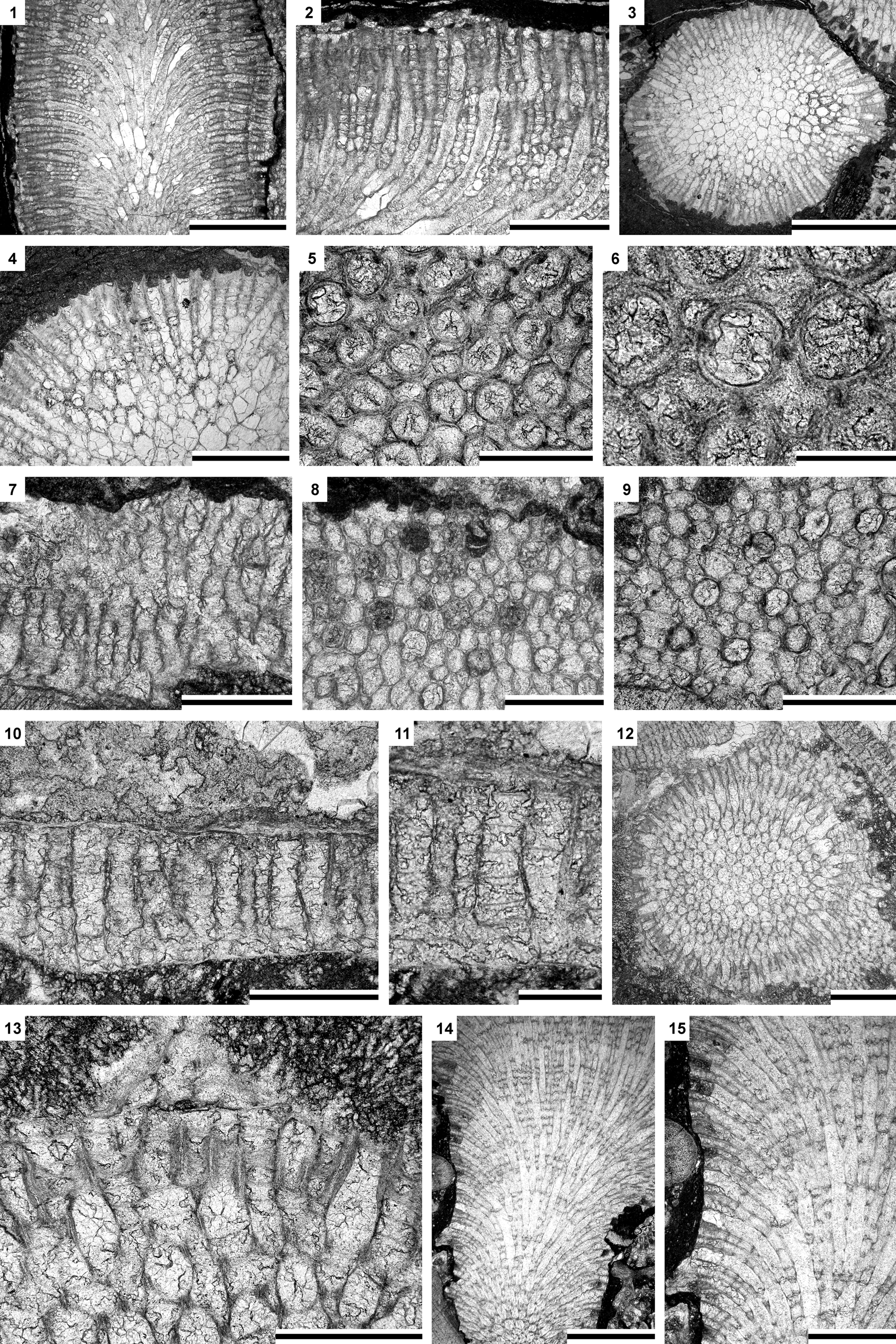
Figure 5. (1–6) Leioclema speciosum Hu, Reference Hu1982: (1) longitudinal section of branched colony, SMF 60587; (2) longitudinal section of exozonal part of branched colony, SMF 60587; (3, 4) transverse section of branched colony, SMF 60586; (5, 6) tangential sections showing autozooecial apertures, acanthostyles, and mesozooecia, SMF 60587; (7–11) Asperopora sinensis n. sp.: (7) longitudinal section, holotype, SMF 60599; (8, 9) tangential sections showing autozooecial apertures, acanthostyles, and mesozooecia, holotype, SMF 60599; (10, 11) longitudinal section showing autozooecia and mesozooecia, paratype, SMF 60606; (12–15) Trematopora jiebeiensis n. sp.: (12, 13) branch transverse section, paratype, SMF 60644; (14, 15) branch longitudinal section, paratype, SMF 60643. Scale bars = 2 mm (1, 3, 14); 1 mm (2, 4, 12, 15), 0.5 m (5, 7–10, 13); 0.2 mm (6, 11).
Table 4. Descriptive statistics of Leioclema speciosum Hu, Reference Hu1982. Abbreviations as for Table 1.
Holotype
NIGP 63767; Cuijiagou Formation, Telychian, Llandovery, lower Silurian; Sichuan Province, China.
Occurrence
Jiebei village, Chongqing, China; lower part of the Hanchiatien Formation, lower Telychian, Llandovery, lower Silurian.
Materials
SMF 60578−60598.
Remarks
Leioclema speciosum is similar to Leioclema tuvaensis Astrova, Reference Astrova1959 from the Wenlock of Tuva (Russia), but differs in having smaller colonies and less abundant and smaller acanthostyles (acanthostyle diameter 0.030−0.045 mm vs. 0.01−0.03 mm [Astrova, Reference Astrova1959], respectively). Leioclema speciosum differs from Leioclema densiporum Owen, Reference Owen1965 from the Llandovery of England in having larger autozooecial apertures (aperture width 0.10−0.20 mm vs. 0.10−0.12 mm [Owen, Reference Owen1965], respectively).
Genus Asperopora Owen, Reference Owen1969
Type species
Callopora aspera Hall, Reference Hall1852; Silurian (Wenlock), New York, USA.
Asperopora sinensis new species
Figure 5.7–5.11; Table 5
Type specimens
Holotype, SMF 60599; paratypes, SMF 60600−60635.
Table 5. Descriptive statistics of Asperopora sinensis n. sp. Abbreviations as for Table 1.
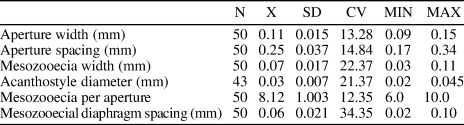
Diagnosis
Thin encrusting colonies; endozones short; autozooecial apertures rounded-polygonal; basal diaphragms few to common in exozone; one or two acanthostyles surrounding each autozooecial aperture; 6−10 mesozooecia surrounding each autozooecial aperture; maculae absent.
Occurrence
Jiebei village, Chongqing, China; lower part of the Hanchiatien Formation, lower Telychian, Llandovery, lower Silurian.
Description
Encrusting colonies, 0.4−1.0 mm thick. Autozooecia budding from a thin epitheca, growing a short distance parallel to the substratum, then bending sharply to the colony surface. Epitheca 0.003−0.005 mm thick. Autozooecial apertures rounded-polygonal. Basal diaphragms common in exozone, thin, slightly deflected orally. Acanthostyles common, one or two surrounding each autozooecial aperture, small, originating in the outer exozone, having distinct calcite cores and dark, laminated sheaths, indenting into autozooecial space. Mesozooecia abundant, 6−10 surrounding each autozooecial aperture, rounded-polygonal in transverse section, originating at the base of exozone, slightly beaded. Autozooecial walls granular, 0.003−0.005 mm thick in endozone, laminated, merged, 0.02−0.03 mm thick in exozone. Maculae not observed.
Etymology
The species is named after its occurrence in China.
Remarks
Asperopora sinensis n. sp. differs from Asperopora bellum (Pushkin, Reference Pushkin, Akimets and Govetskii1976) from the middle Silurian of Belarus and Norway in its slightly smaller autozooecial apertures (mean aperture width 0.11 mm vs. 0.14 mm [Pushkin, Reference Pushkin, Akimets and Govetskii1976], respectively) and in its more abundant mesozooecia (mean 8.12 per aperture vs. 7.4 [Pushkin, Reference Pushkin, Akimets and Govetskii1976], respectively). Asperopora sinensis n. sp. differs from Asperopora aspera (Hall, Reference Hall1852) from the middle Silurian of North America and Europe in its less abundant and larger mesozooecia (mean 8.12 per aperture vs. 10.5 [Ernst et al., Reference Ernst, Munnecke and Oswald2015], respectively), and in having one or two acanthostyles around each autozooecial aperture instead of 1−5 (Ernst et al., Reference Ernst, Munnecke and Oswald2015) in the latter species.
Family Trematoporidae Miller, Reference Miller1889
Genus Trematopora Hall, Reference Hall1852
Type species
Trematopora tuberculosa Hall, Reference Hall1852; lower Silurian (Niagaran), North America.
Trematopora jiebeiensis new species
Figures 5.12–5.15, 6.1–6.3; Table 6
Type specimens
Holotype, SMF 60636; paratypes, SMF 60637−60660.

Figure 6. (1–3) Trematopora jiebeiensis n. sp.: (1) branch longitudinal section showing secondary overgrowth, autozooecia, mesozooecia, and acanthostyles, holotype, SMF 60636; (2, 3) tangential sections showing autozooecial apertures, acanthostyles, and mesozooecia, holotype, SMF 60636; (4–9) Trematopora tenuis n. sp., holotype, SMF 60661: (4, 5) oblique section of a branch; (6) branch transverse section; (7, 8) longitudinal section of a branch showing autozooecia and acanthostyles; (9) tangential sections showing autozooecial apertures, acanthostyles, and mesozooecia. Scale bars = 1 mm (1, 4, 6–8); 0.5 mm (2, 3, 5); 0.2 mm (9).
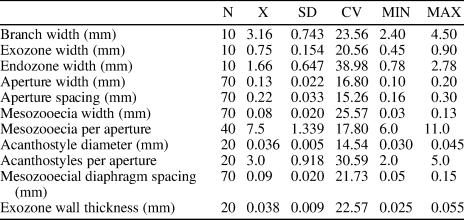
Table 6. Descriptive statistics of Trematopora jiebeiensis n. sp. Abbreviations as for Table 1.
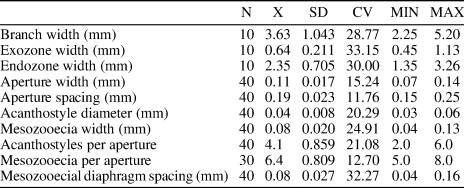
Diagnosis
Branched colonies with distinct exozones; autozooecial apertures rounded to slightly angular; basal diaphragms rare; mesozooecia beaded, 5−8 surrounding each autozooecial aperture; acanthostyles moderately large, 2−6 surrounding each autozooecial aperture, originating in endozone; maculae absent.
Occurrence
Jiebei village, Chongqing, China; lower part of the Hanchiatien Formation, lower Telychian, Llandovery, lower Silurian.
Description
Ramose branched colonies; branch width 2.25−5.20 mm. Exozone distinct, 0.45−1.13 mm wide; endozone 1.35−3.26 mm wide. Secondary overgrowths common. Autozooecia long, polygonal in cross section in endozone, bending sharply in exozone, with rounded to slightly angular apertures. Basal diaphragms rare, thin, concentrated mainly in the transition between exo- and endozones. Mesozooecia abundant, originating at base of exozone, beaded in places of development of diaphragms, 5−8 surrounding each autozooecial aperture. Diaphragms in mesozooecia straight, abundant. Acanthostyles moderately large, prominent, having distinct hyaline cores, 2−6 surrounding each autozooecial aperture, originating in endozone. Autozooecial walls 0.003−0.005 mm thick, granular-prismatic in endozone, showing reversed V-shaped lamination, integrated with locally visible dark border between zooecia, 0.02−0.06 mm thick in exozone. Maculae absent.
Etymology
The species is named after the type locality, the village of Jiebei in China.
Remarks
Trematopora jiebeiensis n. sp. differs from Trematopora sinensis from the lower Silurian (Llandovery) of China in having smaller autozooecial apertures (aperture width 0.07−0.14 mm vs. 0.30−0.36 mm [Hu, Reference Hu1982], respectively). Trematopora jiebeiensis n. sp. differs from Trematopora whitfieldi Ulrich, Reference Ulrich1883 from the Silurian (Wenlock) of North America in its smaller autozooecial apertures (mean aperture width 0.11 mm vs. 0.14 mm [Ulrich, Reference Ulrich1883], respectively).
Trematopora tenuis new species
Figure 6.4–6.9; Table 7
Type specimens
Holotype, SMF 60661; SMF 60662−60667.
Table 7. Descriptive statistics of Trematopora tenuis n. sp. Abbreviations as for Table 1.
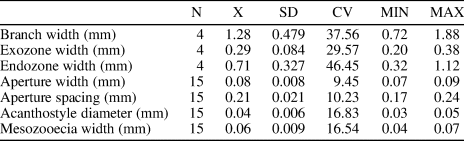
Diagnosis
Thin, branched colonies with distinct exozones; autozooecial apertures rounded to slightly angular; basal diaphragms rare; mesozooecia common, slightly beaded; acanthostyles moderately large, 6−8 surrounding each autozooecial aperture, originating in endozone; maculae absent.
Occurrence
Jiebei village, Chongqing, China; lower part of the Hanchiatien Formation, lower Telychian, Llandovery, lower Silurian.
Description
Ramose branched colonies; branch width 0.72−1.88 mm. Exozone distinct, 0.20−0.38 mm wide; endozone 0.32−1.12 mm wide. Secondary overgrowths not observed. Autozooecia long, polygonal in cross section in endozone, bending sharply in exozone, with rounded to slightly angular apertures. Basal diaphragms rare, thin, concentrated mainly in the transition between exo- and endozones. Mesozooecia common, originating at base of exozone, slightly beaded in places of development of diaphragms. Diaphragms in mesozooecia straight, abundant. Acanthostyles moderately large, prominent, having distinct hyaline cores, 6−8 surrounding each autozooecial aperture, originating at the base of exozone. Autozooecial walls 0.003−0.005 mm thick, granular-prismatic in endozone, showing reversed V-shaped lamination, integrated with locally visible, dark border between zooecia, 0.003−0.005 mm thick in exozone. Maculae absent.
Etymology
The species is named because of its thin branches (Latin tenuis = thin, narrow).
Remarks
Trematopora tenuis n. sp. is similar to Trematopora cristata Kopajevitch, Reference Kopajevitch1984 from the Wenlock of Mongolia, but differs from the latter in having more abundant acanthostyles (6−8 per autozooecial aperture vs. 1−4 [Kopajevitch, Reference Kopajevitch1984], respectively). Trematopora tenuis n. sp. differs from Trematopora minima Ernst in Suttner and Ernst, Reference Suttner and Ernst2007 from the Upper Ordovician of India in its less abundant mesozooecia and more abundant acanthostyles (6−8 per autozooecial aperture vs. four or five [Suttner and Ernst, Reference Suttner and Ernst2007], respectively).
Order Fenestrata Elias and Condra, Reference Elias and Condra1957
Suborder Fenestellina Astrova and Morozova, Reference Astrova and Morozova1956
Family Fenestellidae King, Reference King1849
Genus Moorephylloporina Bassler, Reference Bassler1952
Type species
Moorephylloporina typica Bassler, Reference Bassler1952; Middle Ordovician, Black Riverian, Virginia, USA.
Moorephylloporina parvula new species
Figure 7.1–7.8; Table 8
Type specimens
Holotype, SMF 60668; paratypes SMF 60669−60710.
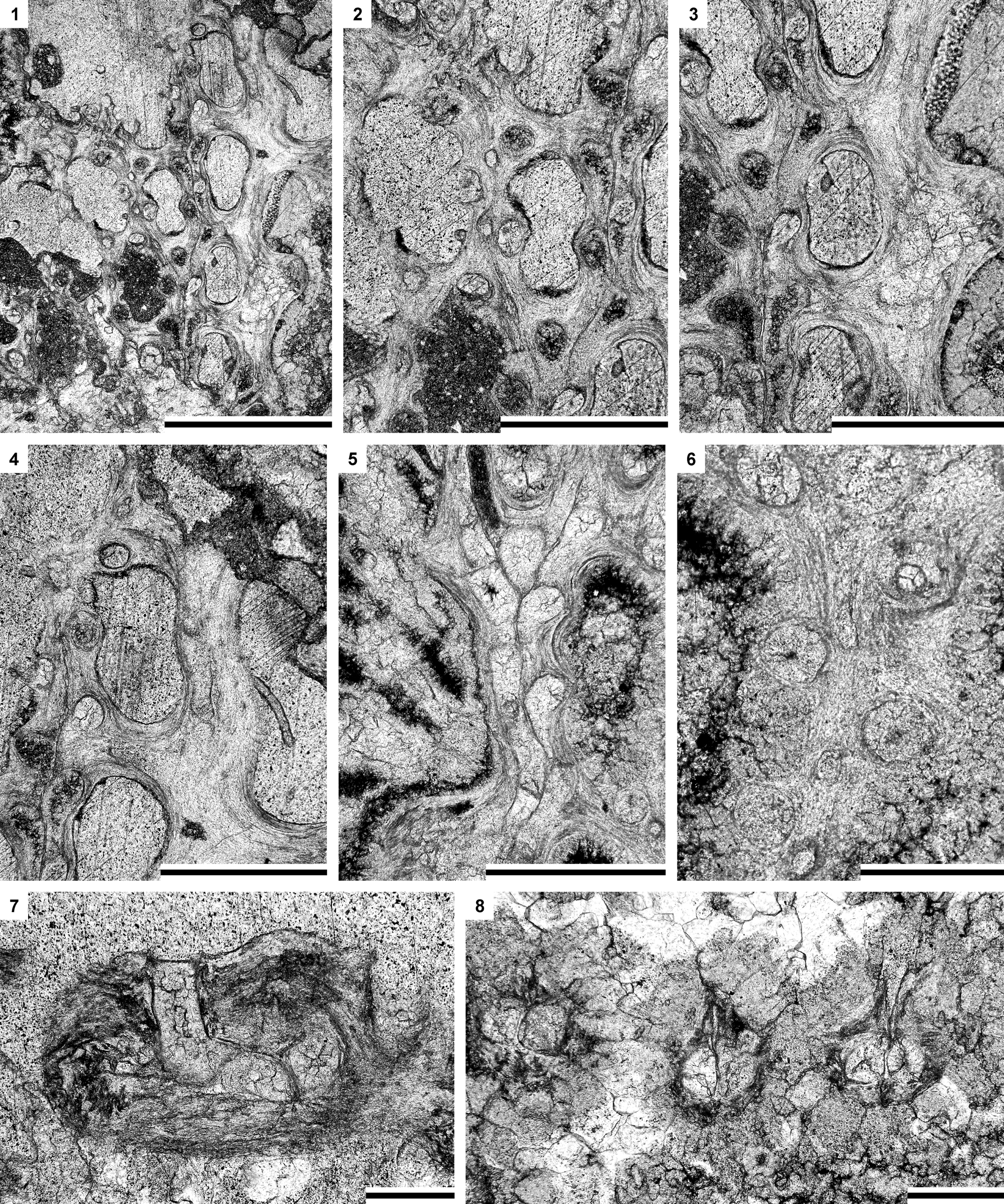
Figure 7. Moorephylloporina parvula n. sp.: (1–4) tangential section showing autozooecial apertures and keel nodes, holotype, SMF 60668; (5) midtangential section showing autozooecial chambers, paratype, SMF 60708; (6) tangential section showing autozooecial apertures and nodes, paratype, SMF 60708; (7) transverse section showing autozooecial chamber, paratype, SMF 60682; (8) transverse section showing autozooecial chambers, paratype, SMF 60702. Scale bars = 1 mm (1); 0.5 mm (2–5); 0.2 mm (6–8).
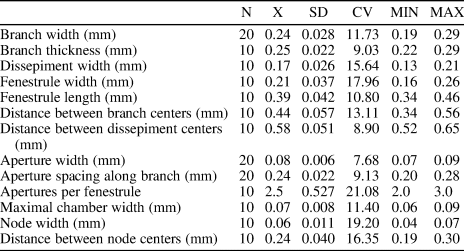
Table 8. Descriptive statistics of Moorephylloporina parvula n. sp. Abbreviations as for Table 1.
Diagnosis
Reticulate colonies with straight branches; dissepiments short, wide; fenestrules small, oval; autozooecial apertures rounded, two or three spaced per fenestrule length; keel wide, low, with high and moderately large nodes; hemisepta lacking; diaphragms present; vesicular skeleton present.
Occurrence
Jiebei village, Chongqing, China; lower part of the Hanchiatien Formation, lower Telychian, Llandovery, lower Silurian.
Description
Reticulate colonies with straight, frequently bifurcating branches, joined by short, wide dissepiments. Autozooecia arranged in two alternating rows on branches, having circular apertures with moderately high peristomes, two or three spaced per length of a fenestrule. Peristomes containing 10−12 nodes. Peristomal nodes 0.015−0.020 mm in diameter. Fenestrules oval. Keels wide, low. Keel nodes high, with moderate diameter and spacing, rounded to oval in their cross sections. Microacanthostyles on the reverse colony surface abundant, regularly spaced in longitudinal rows, 0.010−0.015 mm in diameter.
Interior description.—Autozooecia long, rectangular in the mid-tangential section, with well developed vestibule; axial wall straight; aperture positioned at distal end of chamber. Hemisepta absent. Diaphragms present. Internal granular skeleton thin, continuous, with obverse keel, nodes, peristome, and across dissepiments. Outer lamellar skeleton thin to moderately thick. Vesicular skeleton present.
Etymology
The species name refers to the small size of this species (Latin parvulus = very small).
Remarks
Moorephylloporina parvula n. sp. is similar to Moorephylloporina delicata (Nekhoroshev, Reference Nekhoroshev1961) from the Llandovery of Siberia. The new species differs in its wider branches (branch width 0.19−0.29 mm vs. 0.16−0.19 mm [Nekhoroshev, Reference Nekhoroshev1961], respectively) and smaller fenestrules (fenestrule width 0.16−0.26 mm vs. 0.30−0.40 mm [Nekhoroshev, Reference Nekhoroshev1961], respectively; fenestrule length 0.34−0.46 mm vs. 0.58−0.60 mm [Nekhoroshev, Reference Nekhoroshev1961], respectively).
Discussion
In the Jiebei section, bryozoans occur exclusively in the carbonate interbeds in the lower part of the Hanchiatien Formation (Figs. 2, 3). Characterized by reticular colonies, Moorephylloporina can be found in all types of facies in these meter-scale reefs, indicating its eurytopic nature. However, such distribution could be also explained by taphonomic processes and postmortem transportation of Moorephylloporina fragments from reef areas inhabited by this species within the whole reef. Although the abundance of Moorephylloporina is relatively low in the framestone (Fig. 3.3) of the reefs, these fenestellid bryozoans provide hard substrata for Fistulipora and Asperopora. In contrast, branched Trematopora and Leioclema tend to occur out of the reef core (framework) (Fig. 3.1). Like Champlainopora (Atactotoechus) chazyensis Ross, Reference Ross1963 in some Ordovician reefs (e.g., Cuffey et al., Reference Cuffey, Robison and Mehrtens2002), Trematopora and Leioclema might have formed reef-flank thickets developed under more agitated conditions. Moorephylloporina, Trematopora, and Leioclema represent pioneering bryozoans that were able to grow on weakly lithified substrata. In contrast, encrusting Fistulipora, Hennigopora, and Asperopora relied more on hardgrounds and occupied a large proportion in the reefs, layered on top of one another and so forming densely compact framestones (Li et al., Reference Li, Ernst, Munnecke, Yu, Li and Deng2018) that represent a typical crust-mound stage from an evolutionary ecological perspective (Cuffey, Reference Cuffey2006).
The Telychian of the Upper Yangtze Platform is characterized by terrigenous sediments (e.g., Rong et al., Reference Rong, Wang and Zhang2012), and bryozoans are mostly reported from interbedded argillaceous limestones (or marls) (Hu, Reference Hu1982, Reference Hu1990; Xia and Qi, Reference Xia and Qi1989), probably related to internal-wave deposits (Li et al., Reference Li, Ernst, Munnecke, Yu, Li and Deng2018) in some cases. Systematic studies of Telychian bryozoans are limited compared to other macrofossil groups, e.g., brachiopods, trilobites, and corals. Here, we provide the first detailed comparison of the composition of the bryozoan fauna (Table 9). In the South China block, Asperopora and Moorephylloporina are only reported from the Hanchiatien Formation, whereas Fistulipora and Trematopora are widely distributed across the platform during the early Telychian, recorded in all three studied sections in the literature. Apart from the Jiebei section, Hennigopora and Leioclema have been documented from one other Telychian section, indicating a moderately wide paleogeographic distribution. It is worth noting that bryozoans in some sections are not diverse, or else have been inadequately sampled. Further studies should be carried out to confirm their distributional patterns on the platform.
Table 9. Distribution of bryozoan species in the Telychian localities of South China.
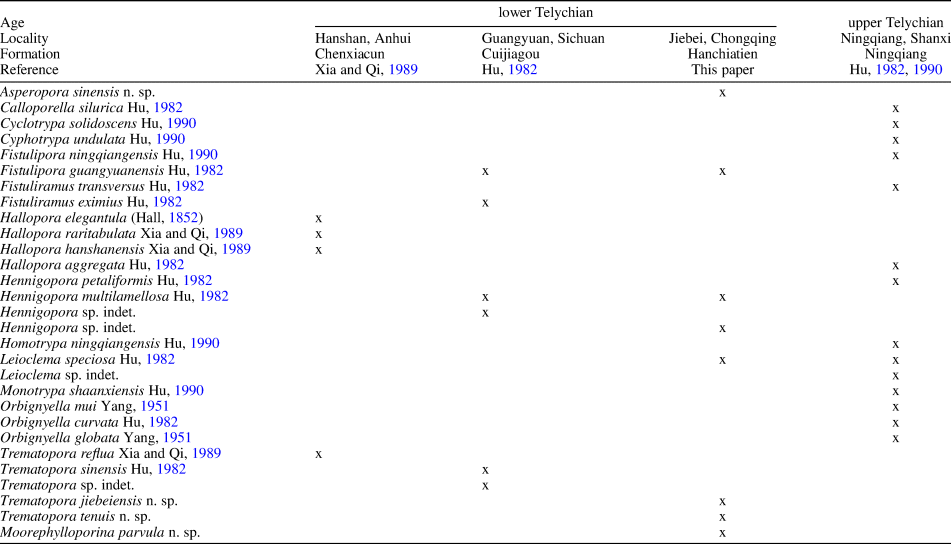
Bryozoans from the Jiebei section belong to genera with predominantly cosmopolitan distributions during the Telychian. Outside of South China, representatives of the genus Fistulipora are known from North America and Siberia. Species of Hennigopora are known from the USA (New York, Indiana) and Siberia. The genus Leioclema is known from North America, Europe (England, Ukraine), and Tuva (Russia). Records of Asperopora are known from North America (New York, Canada), Sweden (Gotland), and Siberia, whereas Trematopora is known from North America and China. The fenestrate genus Moorephylloporina is largely restricted to the Ordovician, except for two species from the lower Silurian (Llandovery) of Siberia, and the new species described here. It disappeared during the lower Silurian.
The early Silurian was a period of exceptional cosmopolitanism for benthic species with dispersive larvae (e.g., Cocks, Reference Cocks2001). Bryozoans, as in other groups of benthic organisms, showed low provinciality (e.g., Tuckey, Reference Tuckey, McKerrow and Scotese1990; Anstey et al., Reference Anstey, Pachut and Tuckey2003; McCoy and Anstey, Reference McCoy and Anstey2010; Buttler et al., Reference Buttler, Wyse Jackson, Ernst, McKinney, Harper and Servais2013). The cluster analysis and detrended correspondence analysis (Fig. 8) reveal distinct clustering of South China with Siberia and Indiana (USA). Bryozoan faunas of Estonia seem to be close to those of New York (USA), whereas Gotland (Sweden) clusters with Anticosti (Canada) and Michigan (USA). These results agree with existing paleogeographical reconstructions for the early Silurian (e.g., Cocks and Torsvik, Reference Cocks and Torsvik2002). However, most existing references for bryozoans in the lower Silurian need critical reassessment, because the descriptions are often far from present-day standards.

Figure 8. Paleobiogeographical affinities of Jiebei bryozoan association and selected Telychian bryozoan faunas (data from Anstey et al., Reference Anstey, Pachut and Tuckey2003, updated 2020): (1) the hierarchical relationship based on pair-group cluster analysis (Euclidean similarity index); (2) detrended correspondence analysis. All analyses performed using PAST (Hammer et al., Reference Hammer, Harper and Ryan2001).
Acknowledgments
This study was supported by the Youth Innovation Promotion Association of the Chinese Academy of Sciences (CAS) no. 2019310, the Natural Science Foundation of China (NSFC) no. 41702003, and CAS no. XDB26000000 and XDA19050101. We are grateful to B. Leipner-Mata (GeoZentrum Nordbayern, Erlangen, Germany) for the preparation of thin sections and to S. Yu (Nanjing Institute of Geology and Palaeontology) for assistance in the field. This study is a contribution to the International Geoscience Projects (IGCP) 653 and 668. QJL is thankful for his invitation as a guest researcher at GeoZentrum Nordbayern. P.D. Taylor (London) and H.A. Nakrem (Oslo) are thanked for their helpful and constructive reviews of the manuscript.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/jpa.2020.86




















 Open access
Open access