



Introduction
Alveolar hydatid disease, also known as alveolar echinococcosis (AE), ranks as one of the most dangerous helminthic zoonoses in the world (Ito et al., Reference Ito, Nakao and Sako2007). Without appropriate medical measures and chemotherapeutic treatment strategies, this cancer-like disease has a death rate of more than 90% within ten years after diagnosis (Torgerson et al., Reference Torgerson, Schweiger, Deplazes, Pohar, Reichen, Ammann, Tarr, Halkik and Müllhaupt2008). Also, the veterinary aspect is of great concern. Animals that are not part of the (sylvatic) transmission cycle, such as pigs, apes or domestic animals such as dogs and cats can not only be (seriously) infected with the disease, but also increase the risk of infection for humans (Deplazes & Eckert, Reference Deplazes and Eckert2001). The causative agent of AE is the metacestode of Echinococcus multilocularis (Deplazes & Eckert, Reference Deplazes and Eckert2001). The distribution of E. multilocularis covers almost the entire Northern Hemisphere. Its endemic areas reach from western Europe to Russia, parts of Asia (including Hokkaido, Japan), Alaska (including St. Lawrence), Canada and the continental United States (Torgerson et al., Reference Torgerson, Keller, Magnotta and Ragland2010; Deplazes et al., Reference Deplazes, Rinaldi and Alvarez Rojas2017).
Echinococcus multilocularis measures only 1.2 mm to 4.5 mm and contains in its proglottis from 200 up to 300 eggs (Thompson, Reference Thompson and Thompson1986; Eckert et al., Reference Eckert, Deplazes, Kern, Palmer, Soulsby, Torgerson and Brown2011). Despite its small body size, E. multilocularis compensates with an enormous mass infestation. In individual cases, more than 100,000 worms can colonize a single host (Fischer et al., Reference Fischer, Reperant, Weber, Hegglin and Deplazes2005). This allows a release of an enormous number of eggs per host and defecation into the environment. For foxes, the definitive hosts, the maximum number of eggs excreted daily was found to be 98,638 (Kapel et al., Reference Kapel, Torgerson, Thompson and Deplazes2006). The consequential contamination of the environment can become a dangerous infection source.
Echinococcus multilocularis eggs belong to the Taeniidae. All of this family's eggs are similar in terms of their morphology (Lawson & Gemmell, Reference Lawson and Gemmell1983). They are round–elliptical in shape (Thompson, Reference Thompson and Thompson1986). The size of E. multilocularis eggs varies between 28 × 39 μm and 30 × 40 μm (Veit et al., Reference Veit, Bilger, Schad, Schäfer, Frank and Lucius1995).
They are ‘extremely resistant, enabling them to withstand a wide range of environmental temperatures’ (Thompson, Reference Thompson and Thompson1986) while sensitive to aridity and high temperatures (Lawson & Gemmell, Reference Lawson and Gemmell1983).
During an experiment in Germany exposing E. multilocularis eggs to natural climatic conditions, the eggs survived a maximum of 240 days during autumn and winter and 78 days in summer. Generally, it can be assumed that the eggs usually survive well over 100 days in the environment (Veit et al., Reference Veit, Bilger, Schad, Schäfer, Frank and Lucius1995).
The main infection path is hand–mouth contact after direct contiguity with affected primary hosts, namely foxes, but in some cases also dogs and other animals (see above), because the eggs get stuck in their fur (Conraths & Deplazes, Reference Conraths and Deplazes2015; Merck, 2018). Also, the handling of contaminated soil is mentioned (Romig et al., Reference Romig, Deplazes, Jenkins, Giraudoux, Massolo, Craig, Wassermann, Takahashi and De La Rue2017). Furthermore, E. multilocularis is third in the global ranking of food-borne parasites. Therefore, a transmission and infection by contaminated food, such as fruits, vegetables or even water should not be dismissed (Food and Agriculture Organization of the United Nations & World Health Organization, 2014). The routes of transmission and dispersion of E. multilocularis, respectively its eggs, are of highly complex pattern, yet, still insufficiently researched (Lawson & Gemmell, Reference Lawson and Gemmell1983; Ishikawa et al., Reference Ishikawa, Ohga and Doi2003; Tolnai et al., Reference Tolnai, Széll and Sréter2013; Hegglin et al., Reference Hegglin, Bontadina and Deplazes2015).
There have been prior studies using mathematical models to describe transmission dynamics of E. multilocularis for estimating the risk of infection. These models considered influences of important mechanisms and variables regarding the transmission of the parasite (e.g. treatment of foxes, age structures, seasonality, habitat types, deworming of dogs, environmental disinfection, etc.) (Atkinson et al., Reference Atkinson, Williams, Yakob, Clements, Barnes, McManus, Yang and Gray2013; Lewis et al., Reference Lewis, Otero-Abad, Hegglin, Deplazes and Torgerson2014; Hassan & Munganga, Reference Hassan and Munganga2019; Khan et al., Reference Khan, Ahmed, Sohail, Alam and Simsek2020; Woolsey & Miller, Reference Woolsey and Miller2021). While these models provide valuable insights into modulating infection risks, they ignore that there is a second category of transmission – the abiotic distribution. Meanwhile a well-known fact that dispersion of E. multilocularis eggs can be affected by various biotic factors such as herbivores, birds, numerous insects, especially flies, different beetles, butterflies, snails, etc. the abiotic distributors of eggs, rain and wind (Guo et al., Reference Guo, Xiao, Wang, Liu, Tang and H2017) are longstanding objects of controversial discussion (Lawson & Gemmell, Reference Lawson and Gemmell1983). To this day, the evidence that wind could contribute to the dispersal of E. multilocularis eggs has been absent. A study detecting a uniform radial dispersion of Taenia hydatigena eggs may argue against a dispersion by wind (Gemmell et al., Reference Gemmell, Johnstone and Boswell1978). In contrast, indications for the wind-borne dispersion of Echinococcus granulosus eggs have been shown by several studies (Bourée, Reference Bourée2001; Sánchez Thevenet et al., Reference Sánchez Thevenet, Alvarez, Torrecillas, Jensen and Basualdo2019). This supports the hypothesis of the involvement of wind also for E. multilocularis. Therefore, it is of huge importance to investigate the role of wind. If wind has an influence on transmission dynamics of E. multilocularis eggs, this could increase the risk of infection for AE.
Material and methods
Flight model
To analyse the possible epidemiological impact of wind, we developed a mathematical flight model.
In order to develop such a model, it is necessary to understand the relationships between the physical parameters that enable the flight of an E. multilocularis egg. Those dependencies are shown in fig. 1. The graphic (fig. 1a) explains the relationships. The starting point (S) (shown here as a stylized tree stump) determines the take-off height (h). The wind acts on the egg with the wind speed (u) and carries it on. As soon as the egg starts its flight, gravity forces it back to earth at its specific rate of descent (v) – proportional to the wind speed (u). The distance (e), that the egg covers during this process to the landing point (Z), gives the flight distance of the egg. The flight range is described by the same angle (∝) as the rate of descent in relation to the wind speed and the take-off height in relation to the distance. This flight path angle is determined as the equation in fig. 1b. Rearranging this equation to e results in the final formula for the calculation of the flight length (fig. 1c).
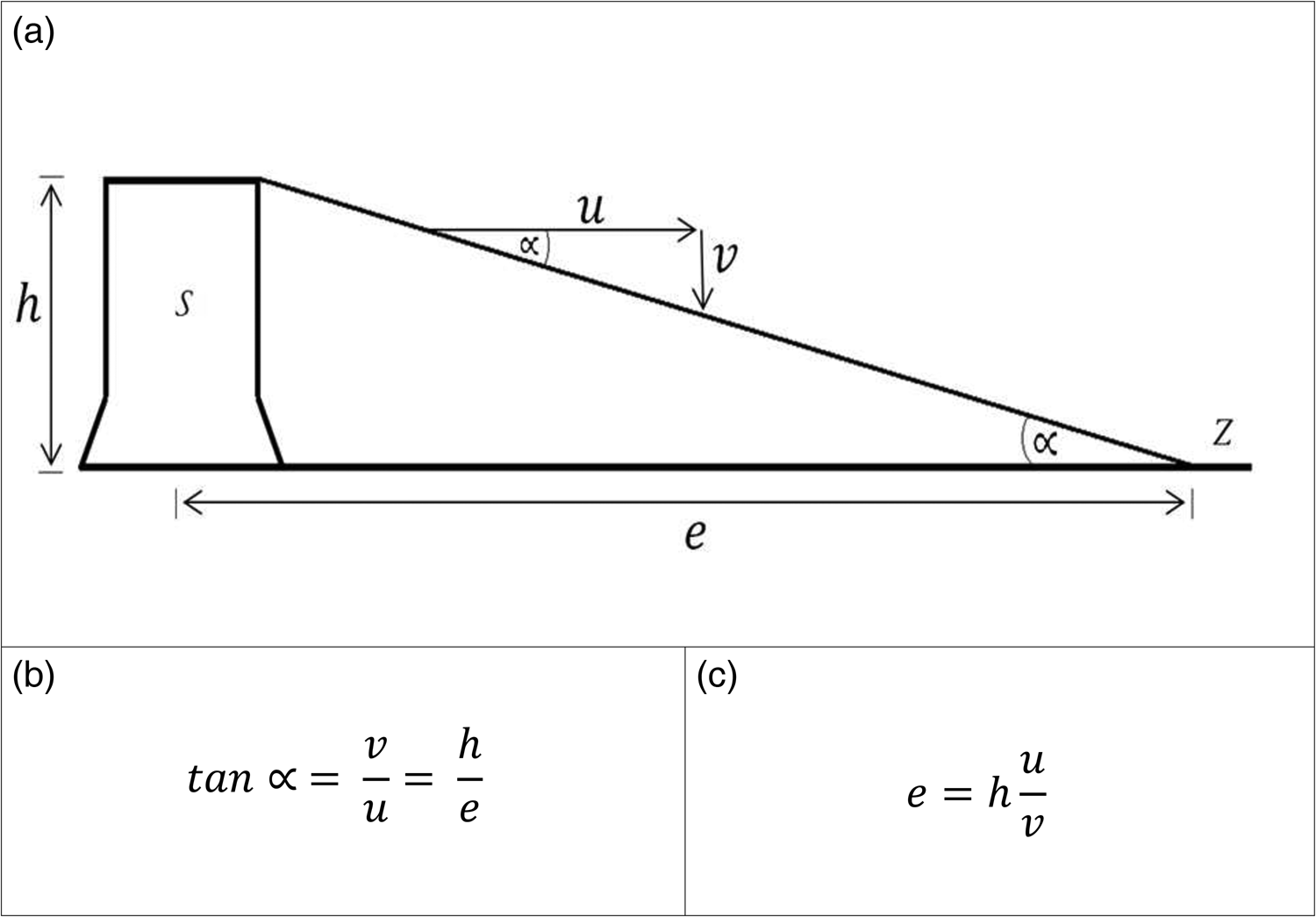
Fig. 1. Flight model describing the physical parameters and dependencies influencing the flight of an Echinococcus multilocularis egg (a) with mathematical description of the flight path angle (b) and the final formula for flight path calculation (c). h = take-off height, u = wind speed, v = rate of descent, e = distance, S = starting point (tree stump), Z = zero (finish).
This simple model calculates the minimal flight distance of an E. multilocularis egg depending on the constant ‘rate of descent’ (v) and the variable parameters ‘wind speed’ (u) and ‘starting altitude’ (h), provided that the flight is not disturbed.
Determination of the ‘rate of descent’ (v)
Two important forces are affecting the egg (fig. 2a): the frictional resistance (W) (fig. 2b) (Bird et al., Reference Bird, Stewart and Lightfoot2007) which has to be overcome in order to enable transport by air; and the gravitation (G) (fig. 2c) (Avison, Reference Avison1989) which forces the flying object back to the ground. The flight ability of an ovum is therefore largely dependent on its rate of descent.

Fig. 2. Graphical illustration describing the two important forces affecting the egg (a). Formula of frictional resistance (b) and gravitation (c) equated (d and e) and rearranged for calculation of the rate of descent (v) for an Echinococcus multilocularis egg. (Avison, Reference Avison1989; Bird et al., Reference Bird, Stewart and Lightfoot2007) W = frictional resistance (according to Stokes), η = dynamic viscosity of the fluid (liquid or gas) in which the body moves (here: 18.2 ⋅ 10−6 Nsec./mm2), r = radius (here: 0.017125 ⋅ 10−3 m), v = rate of descent, G = gravitation, m = mass (here: 0.024502 ⋅ 10−9 kg), g = gravitational acceleration (here: 9.81 m/sec2).
To determine the rate of descent, the size and the mass of an E. multilocularis egg are required. While the average size of these eggs is known (Veit et al., Reference Veit, Bilger, Schad, Schäfer, Frank and Lucius1995), we needed to determine the weight of an E. multilocularis egg. Figure 3 shows the mathematical methods used to approximate the volume and weight. The diameter measures between 28 × 39 μm and 30 × 40 μm (Veit et al., Reference Veit, Bilger, Schad, Schäfer, Frank and Lucius1995). In our calculations (fig. 3) we applied the mean value 0.029 × 0.0395 mm. According to the round but slightly elliptical shape of the egg (Thompson, Reference Thompson and Thompson1986; Veit et al., Reference Veit, Bilger, Schad, Schäfer, Frank and Lucius1995), we decided to calculate three different volumes and intersect them, to incorporate the special shape of the E. multilocularis egg as closely as possible in our approximation. Therefore, volume values were determined for both an ellipsoid (fig. 3a) (Weisstein, Reference Weisstein2002) and a sphere (fig. 3b) (Weisstein, Reference Weisstein2002). The spherical volume was calculated for a ‘small’ sphere (related to 0.029 mm), and for a ‘large’ sphere (related to 0.0395 mm). Both volumes were averaged. This ‘mean sphere’ was intersected with the volume of the ellipsoid, resulting in an E. multilocularis volume of 2 ⋅ 10−5 mm3.

Fig. 3. Formula for calculating the volume of an ellipsoid (a) and a sphere (b). (Weisstein, Reference Weisstein2002) V = volume, r = radius, a, b, c = half-axes.
Since the specific weight of eggs of the Taeniidae family is given as 1.2251 g/ml (David & Lindquist, Reference David and Lindquist1982), the weight of a single E. multilocularis egg equals 0.024502 μg.
In order to determine the rate of descent of the egg, W and G are equated (figs. 2d and e) and rearranged to v (fig. 2f). Consequently, the rate of descent is v = 4.09 ⋅ 10−2 m/s, respectively, v = 40.9 mm/s.
Determination of wind speeds (u)
To obtain reliable wind data for applying into the model, data sets originating from a dense stocked Norway spruce (Picea abies) stand (test area F1) in the Solling (Germany) were accessed (Meesenburg et al., Reference Meesenburg, Meiwes and Bartens1999; Paar et al., Reference Paar, Evers, Eichhorn, Paar, Evers and Eichhorn2013).
We averaged the hourly measured wind speeds of a ten-year period (1990 to 1999) as follows: maximum and minimum values of each month of each year were read out of the data pool, while error-values were ignored. In step two these minima and maxima were allocated to the four seasons of the year. In a third step, the determination of the means per season per year followed, as well as the mean over the entire year. The fourth step was the compilation of all mean values of the individual years and their seasons in a table in order to determine the mean values over all ten years. By thi step, the ten-year mean values at a height of 1 m for spring, summer, autumn, winter and as an annual total could be generated.
Determination of starting heights (h)
Based on the fox's behaviour about the placement of faeces in forest areas, there are many assumable heights for potential starting points (altitudes) of E. multilocularis eggs (e.g. litter, tree stumps, stones, trunks, etc.) (Goszczyński, Reference Goszczyński1990). Consequently, to the behaviour (Goszczyński, Reference Goszczyński1990) and body size (Cavallini, Reference Cavallini1995) of foxes, we took the heights 0.3, 0.5, 0.7, 0.8, 0.9 and 1.0 (in m) into consideration. Since wind speed data of only a height of 1 m were available, we needed to extrapolate the wind speeds for these heights using the formula in fig. 4 (Gualtieri & Secci, Reference Gualtieri and Secci2011). Here, the roughness length is required. The roughness length is a fictitious height above the ground at which the average wind speed approaches zero. This (vertical) length, usually named z0, is used in the logarithmic law of wind and describes the roughness of the ground surface, for example, the presence of grass or bushes (American Meteorological Society, 2019). The roughness length for dense areas of the forest (e.g. stocked stands) is given 0.8 m and 0.2 m for less dense areas (e.g. windthrow areas) (Pertmann, Reference Pertmann2011). By applying these values, we were able to extrapolate the wind speeds downwards using the formula in fig. 4 (Gualtieri & Secci, Reference Gualtieri and Secci2011) and thereby to simulate the flight of an E. multilocularis egg in two different environments: dense forest areas (with z0: 0.8 m, starting altitudes (in m): 1.0, 0.9, 0.8); and less dense forest areas (with z0: 0.2 m, starting altitudes (in m): 0.7, 0.5, 0.3). A further extrapolation downwards would be superfluous, since at heights below the roughness length calm prevails.

Fig. 4. Formula for the extrapolation of wind speeds (Gualtieri & Secci, Reference Gualtieri and Secci2011). v 2 = wind speed in m/s in the height h2, v 1 = reference wind speed in m/s measured on the reference height h1, z 0 = roughness length, h 2 = height above the ground, ln = natural logarithm.
Results
As described in the material and methods section, we developed the following mathematical model as a practical approach to estimate flight lengths of E. multilocularis eggs in dependence of wind speed and starting altitude summarized by the definitive formula (fig. 5).

Fig. 5. Formula to estimate flight lengths of Echinococcus multilocularis eggs. h = take-off height, u = wind speed, e = distance.
Table 1 presents the calculated flight distances of an E. multilocularis egg in a (coniferous) forest stand depending on the wind speed (Wind), the take-off height (Height) above the ground and – with extrapolated values – the roughness length for the two scenarios ‘Forest area (dense)’ and ‘Forest area (less dense)’. The model analysis has revealed, that in forest areas, an egg can be transported several metres by wind and that its flight distances are influenced by season. Further it showed the more the roughness length is lowered, the further the transport length rises.
Table 1. Flight length of an Echinococcus multilocularis egg in relation to the mean wind speed of a whole year according to the ten-year average and in relation to the mean wind speeds in seasons according to the ten-year average.

The values given in boldface represent the final calculated flight distances (output), while values in regular type represent parameters implemented in the flight model/equations (input).
Flight distances – overall
As can be seen in table 1, the results of this analysis reveal in the overall average of the analysed ten-year period the following: at a starting altitude (height) of 1 m above the ground, an E. multilocularis egg is carried approximately 17 m by the wind. From a starting point with a height of 90 cm, this distance is roughly halved. On less dense forest areas (with a lower roughness length), the egg can still fly more than 9 m from a starting point with a height of 70 cm. As soon as the start takes place only 50 cm above the ground, the E. multilocularis egg manages to cover a distance of almost 5 m. At a starting altitude of 30 cm, the wind still launches the egg, but just a little more than 1 m from its original starting point it will reach the ground.
Flight distances – seasons (spring, summer, autumn and winter)
It is noticeable when comparing the four seasons (table 1), in both cases, concerning the dense and the less dense forest areas, deviations regarding the flight distances in the overall annual mean can be observed. The flight range of the egg tends to decrease in spring. This declining tendency is found reinforced in summer, where also the lowest values occurred. In autumn, the flight length is slightly higher than the annual average. Here, the egg is transported about 60 cm further at a starting height of 1 m, compared to the annual average. The longest flight distances emerged in winter. At a height of 1 m, the trajectory was approximately 1.5 m longer than the annual standard value. Between summer and winter seasons, the achieved flight length differs by approximately 3 m.
Discussion
As outlined in the introduction, the possible distribution paths of the E. multilocularis eggs are generally uncertain, of highly complex pattern and showing knowledge gaps (Lawson & Gemmell, Reference Lawson and Gemmell1983; Eckert & Thompson, Reference Eckert and Thompson2017). At least since 1981 the impact of wind as one possible vector in the infection process of E. multilocularis is discussed (World Health Organization, 1982). Over time, this aspect and the research on it faded into the background.
In this study, the rate of descent of E. multilocularis eggs was calculated to verify whether an egg is physically able to be transported by wind. We found the eggs rate of descent is similar to hornbeam pollen (Carpinus betulus) (Rempe, Reference Rempe1937). In the context of this work, the theoretical flight ability of E. multilocularis eggs can be regarded as confirmed. In addition, a mathematical model was developed to calculate possible flight distances of the egg in forest areas depending on starting altitude and wind speed. In dependence of its (examined) starting altitudes, the flight lengths varied between approximately 8 m (height: 0.9 m) and approximately 17 m (height: 1 m) in dense forest areas and between 1.3 m (height: 0.3 m) and 9.38 m (height: 0.7 m) in less dense forest areas. While in the spring and summer seasons, the reached distances slightly declined, in winter and autumn they exceeded the overall average. Therewith, wind-related spreading of eggs is possible.
To the knowledge of the authors, these results are the first of their kind. These findings, being in line with those of uttering of Guo et al. (Reference Guo, Xiao, Wang, Liu, Tang and H2017), confirm the assumptions of the World Health Organization from 1981 that wind is a possible vector of E. multilocularis eggs (World Health Organization, 1982). They differ from the voices of those who argued against the involvement of wind as a possible distributor of E. multilocularis eggs (Lawson & Gemmell, Reference Lawson and Gemmell1983). Furthermore, this work not only confirms the transport by wind, but also, by introducing a simple mathematical model with high practical applicability on a wide range of landscapes, helps to estimate flight lengths and thereby the impact of wind as an epidemiological vector (in this study done for forest landscapes). The impact of different starting altitudes, landscape situations and climatic situations (seasons) could be shown. Compared to previously observed dispersals of Echinococcus and Taenia eggs, in which wind may have been involved, our calculated flight distances are settled below the distances from the deposition site found in real endemic scenarios (e.g. 115 m (Sánchez Thevenet et al., Reference Sánchez Thevenet, Alvarez, Torrecillas, Jensen and Basualdo2019) and 180 m (Bourée, Reference Bourée2001)). This could be explained by the different landscape types and the associated higher wind speeds, as well as the simultaneous influence of biotic vectors (e.g. beetles, flies and birds).
The significance of potential flight ranges, of course, are difficult to assess, regarding the risk to humans without determining the location of the starting point (e.g. distances to hiking trails, tourist attractions, etc.). Nevertheless, airborne spreading of E. multilocularis eggs, especially in fox-inhabited areas, can therefore be assumed as a considerable risk to hikers and berry pickers. The shorter flight distances, which were determined for spring and summer, can be seen as a positive factor reducing the potential risk of infection due to higher temperatures and aridity, which substantially reduce the viability of the eggs. Anyhow, the plentiful presence of fruits and berries, especially since these vector plants are often located on shady and cooler areas which could increase the survival time, can buffer the shorter flight lengths by serving as potential infection sources. While in the autumn and winter seasons flight distances raise due to higher windspeeds. Increased humidity and lower temperatures enhance the durability of the eggs. The resulting risk of infection appears to be notably increased. A mitigating factor in relation to the hand–mouth infection during these months is the extensive lack of fruits (berries) – except for mushrooms in autumn.
Anyhow, some limitations of this model are worth noting. Although the flight lengths are an exciting first glimpse of the impact of wind, a more complex model should be developed in which non-linear flight paths, multiple restarts, vertical and lateral wind movements, respectively, and updraughts are considered. The model presumes that the flight is not disturbed. Under real conditions, this might not always be the case. Obstacles, such a bushes, shrubs, ground vegetation or collisions with trees or animals may interrupt the flight. Nevertheless, eggs that got stuck in this way can be available for further (airborne) transport. Furthermore, the flight model neglects mechanical and thermal turbulence which could abbreviate the flight or catapult the eggs to higher altitudes with higher wind speeds and hence extend the flight of the E. multilocularis egg – similarly to observation with pollen (Miki et al., Reference Miki, Kawashima, Kobayashi, Takeuchi, Tseng and Nakamura2022). In the model the flight of a single egg is simulated. Under real conditions, however, it can be assumed that a group of eggs will start in the majority of flight events. Just as pollen generally do not fly solitary, but rather move in cloud structures (Hattemer et al., Reference Hattemer, Ziehe, Finkeldey, Fromm, Müller-Starck and Schubert2001), this could also be assumed for E. multilocularis eggs, concomitant with a raised potential of contamination and infectivity.
The demonstrated model can be applied to a wide range of simulations. Consequently, there calculations should be conducted for other forest stands such as managed needle stands, deciduous stands, mixed stands and further terrains such as fields, meadows and urban areas, where much higher wind speeds are expected.
For further verification it would be of utmost interest to put such calculations in context to other real data such as the prevalence of infected foxes, rodents or humans in a chosen region under continuous investigation. Additionally, the question whether E. multilocularis eggs lose or retain their contagiousness after transport by wind remains open and requires further research.
Despite the limitations shown, the model provides clear information. The resulted flight lengths, which vary greatly depending on the population structure and starting altitude, underline the potential epidemiological importance of wind with regard to the spread of E. multilocularis eggs. It is hoped that these findings will bring ‘new life’ in the research about wind as an impact factor on the infection paths of E. multilocularis. Future work should consolidate the initial gained knowledge by conducting a field experiment with accurate surrogate eggs.
Acknowledgements
We thank Andreas Dillmann from the German Aerospace Institute in Göttingen for his support in developing a flight length model and Heinrich Kreilein from the Bioclimatology Department of the Faculty of Forest Sciences and Forest Ecology in Göttingen, who made the wind data accessible. Furthermore, we thank the anonymous reviewers for their careful reading of our manuscript and their insightful comments and suggestions.
Author contributions
All of the authors made significant contributions to the concept, design, execution and interpretation of the research and the development of the outline. S.N. proposed and conducted the research and supervised the first draft. S.S. discussed and shaped the research and improved upon the manuscript.
Financial support
The authors declare that there was no financial support for this paper.
Conflict of interest
None.
Ethical approval
None.







 Open access
Open access