



No CrossRef data available.
Published online by Cambridge University Press: 22 May 2023
We consider a branching random walk on a d-ary tree of height n (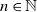 $n \in \mathbb{N}$), in the presence of a hard wall which restricts each value to be positive, where d is a natural number satisfying
$n \in \mathbb{N}$), in the presence of a hard wall which restricts each value to be positive, where d is a natural number satisfying 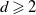 $d\geqslant2$. We consider the behaviour of Gaussian processes with long-range interactions, for example the discrete Gaussian free field, under the condition that it is positive on a large subset of vertices. We observe a relation with the expected maximum of the processes. We find the probability of the event that the branching random walk is positive at every vertex in the nth generation, and show that the conditional expectation of the Gaussian variable at a typical vertex, under positivity, is less than the expected maximum by order of
$d\geqslant2$. We consider the behaviour of Gaussian processes with long-range interactions, for example the discrete Gaussian free field, under the condition that it is positive on a large subset of vertices. We observe a relation with the expected maximum of the processes. We find the probability of the event that the branching random walk is positive at every vertex in the nth generation, and show that the conditional expectation of the Gaussian variable at a typical vertex, under positivity, is less than the expected maximum by order of 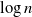 $\log n$.
$\log n$.
 $|\nabla \phi|^p$
surfaces: a large deviation bound for all
$|\nabla \phi|^p$
surfaces: a large deviation bound for all
 $p\geq 1$
. Boll. Unione Mat. Ital. 10, 451–466.CrossRefGoogle Scholar
$p\geq 1$
. Boll. Unione Mat. Ital. 10, 451–466.CrossRefGoogle Scholar $d \geq 3$
. Commun. Math. Phys. 206, 447–462.CrossRefGoogle Scholar
$d \geq 3$
. Commun. Math. Phys. 206, 447–462.CrossRefGoogle Scholar