


Introduction
Increased reliance on intensive animal production systems that utilise high-energy, vegetable-based feed ingredients has caused a general shift in the fatty acid composition of food animals and food-animal products. Modern intensive production systems result in foods that typically contain lesser amounts of omega-3 polyunsaturated fatty acids (PUFAs) and greater amounts of saturated fatty acids compared with foods derived from free-range and wild-caught animals (Raper et al., Reference Raper, Cronin and Exler1992). For several reasons this shift has raised significant public health concerns. A general decline in omega-3 fatty acid consumption is of concern because of the crucial roles these PUFAs play in human growth, development, physiologic function (Connor and Neuringer, Reference Connor and Neuringer1988, Das, Reference Das2006a) and because of their role in suppressing or preventing inflammation linked with cancer, cardiovascular diseases, and type-2 diabetes (Das, Reference Das2008, Azrad et al., Reference Azrad, Turgeon and Demark-Wahnefried2013). A general rise in the consumption of medium-chain, saturated fatty acids and trans fatty acids is similarly of concern because of their strong association with the development of obesity (Neal et al., Reference Neal, Sacks, Swinburn, Vandevijvere, Dunford, Snowden, Webster, Barquera, Friel, Hawkes, Kelly, Kumanyika, L'abbe, Lee, Lobstein, Ma, Macmullan, Mohan, Monteiro, Rayner, Sanders and Walker2013) and diseases such as cancer, cardiovascular diseases (Mente et al., Reference Mente, de Koning, Shannon and Anand2009, Siri-Tarino et al., Reference Siri-Tarino, Sun, Hu and Krauss2010, Santos et al., Reference Santos, Oliveira and Lopes2013), and type-2 diabetes (Ney, Reference Ney1991, Hu et al., Reference Hu, van Dam and Liu2001, Bauman and Griinari, Reference Bauman and Griinari2003, Das, Reference Das2006b, Riserus et al., Reference Riserus, Willett and Hu2009).
The essential PUFAs linoleic acid (LA) and α-linolenic acid (ALA) are used in the body to synthesise their longer chain derivatives required by humans; the omega-6 fatty acid arachidonic acid (ARA) is derived from LA, whereas the omega-3 fatty acids eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) are synthesised from ALA. Each of these PUFA can also be obtained directly through diet. The EFSA Panel on Dietetic Products, Nutrition, and Allergies (NDA) have proposed that an adequate daily intake of DHA and EPA of 250 mg for adults and 100 mg for infants and young children (6 months – 24 months old; EFSA, 2010). In pregnant and nursing women, a DHA intake of 300 mg/d is recommended (Simopoulos et al., Reference Simopoulos, Leaf and Salem1999) and in children (2–18 years old) a DHA intake of 300 to 400 mg/d is regarded as nutritionally desirable (Schuchardt et al., Reference Schuchardt, Huss, Stauss-Grabo and Hahn2010). However, based on a National Health and Nutrition Examination Survey between 2003–2008, mean dietary intake of DHA from foods in the United States was estimated to average only 51 and 75 mg/d for women and men, respectively (Papanikolaou et al., Reference Papanikolaou, Brooks, Reider and Fulgoni2014). To address these nutritional shortfalls, research has been conducted to develop DHA-enriched foods to help bolster intake levels without necessitating major changes in eating habits.
Supplementation using DHA-rich microalgae sources has been used successfully in monogastric livestock such as pigs and poultry to produce DHA-enriched meat and eggs (Bourre, Reference Bourre2005, Rymer and Givens, Reference Rymer and Givens2005, Cheng et al., Reference Cheng, Ou, Shen and Ding2006, Sardi et al., Reference Sardi, Martelli, Lambertini, Parisini and Mordenti2006, Meadus et al., Reference Meadus, Duff, Uttaro, Aalhus, Rolland, Gibson and Dugan2010, Fraeye et al., Reference Fraeye, Bruneel, Lemahieu, Buyse, Muylaert and Foubert2012, Moran et al., Reference Moran, Fusconi, Morlacchini and Jacques2017a,Reference Moran, Fusconi, Morlacchini and Jacquesb). Similarly, attention has been directed to producing DHA-enriched foods from ruminants, however, the extensive lipolysis and subsequent biohydrogenation of unsaturated fatty acids that occurs in the rumen presents unique challenges in achieving this goal (Jenkins et al., Reference Jenkins, Wallace, Moate and Mosley2008, Lourenco et al., Reference Lourenco, Ramos-Morales and Wallace2010, Shingfield et al., Reference Shingfield, Bonnet and Scollan2013).
Numerous studies have examined the effects of algae-derived supplements on ruminant feed intake, enteric methane production, and milk composition and yield (Franklin et al., Reference Franklin, Martin, Baer, Schingoethe and Hippen1999, Papadopoulos et al., Reference Papadopoulos, Goulas, Apolstolaki and Abril2002, Reynolds et al., Reference Reynolds, Cannon and Loerch2006, Boeckaert et al., Reference Boeckaert, Vlaeminch, Fievez, Maignien, Dijkstra and Boon2008a, Boeckaert et al., Reference Boeckaert, Vlaeminck, JDijkstra, Issa-Zacharia, Van Nespen, Can Straalen and Fievez2008b, Or-Rashid et al., Reference Or-Rashid, GKramer, Wood and McBride2008, AbuGhazaleh et al., Reference AbuGhazaleh, Potu and Ibrahim2009, Angulo et al., Reference Angulo, Mahecha, Nuernberg, Nuernberg, Dannenberger, Olivera, Boutinaud, Leroux, Albrecht and Bernard2012, Glover et al., Reference Glover, Budge, Rose, Rupasinghe, MacLaren, Green-Johnson II and Fredeen2012, Stamey et al., Reference Stamey, Shepherd, de Veth and Corl2012, Moate et al., Reference Moate, Williams, Hannah, Eckard, Auldist, Ribaux, Jacobs and Wales2013). Researchers have found that feeding algae may depress milk fat concentration (Franklin et al., Reference Franklin, Martin, Baer, Schingoethe and Hippen1999, Papadopoulos et al., Reference Papadopoulos, Goulas, Apolstolaki and Abril2002, Boeckaert et al., Reference Boeckaert, Vlaeminck, JDijkstra, Issa-Zacharia, Van Nespen, Can Straalen and Fievez2008b, Moate et al., Reference Moate, Williams, Hannah, Eckard, Auldist, Ribaux, Jacobs and Wales2013), increase milk omega-3 fatty acid concentration (Franklin et al., Reference Franklin, Martin, Baer, Schingoethe and Hippen1999, Papadopoulos et al., Reference Papadopoulos, Goulas, Apolstolaki and Abril2002, Boeckaert et al., Reference Boeckaert, Vlaeminck, JDijkstra, Issa-Zacharia, Van Nespen, Can Straalen and Fievez2008b, Stamey et al., Reference Stamey, Shepherd, de Veth and Corl2012), indirectly increase milk conjugated linoleic acid (CLA) concentration (Franklin et al., Reference Franklin, Martin, Baer, Schingoethe and Hippen1999, Boeckaert et al., Reference Boeckaert, Vlaeminck, JDijkstra, Issa-Zacharia, Van Nespen, Can Straalen and Fievez2008b, Or-Rashid et al., Reference Or-Rashid, GKramer, Wood and McBride2008, AbuGhazaleh et al., Reference AbuGhazaleh, Potu and Ibrahim2009), and inhibit voluntary dry matter intake (Franklin et al., Reference Franklin, Martin, Baer, Schingoethe and Hippen1999).
The objective of this study was to evaluate the efficacy of an unextracted algal (Aurantiochytrium limacinum) supplement ALL-G-RICH®, high in DHA and produced under heterotrophic and low sodium conditions, when fed to mid-lactation dairy cattle under commercial conditions on feed intake, milk yield and milk composition.
Material and methods
Animals and diets
The research protocol and animal care were in accordance with guidelines on the protection of animals used for scientific purposes (European Parliament and the Council of the European Union, 2010). After a 10-day pre-experimental adaptation period, 36 multiparous (n = 2.5 ± 0.92) Italian Friesian cows (622.3 ± 71.7 kg) were allocated based on parity, milk yield, and days in milk (DIM) into two similar groups (18 cows per group). In a randomised complete block design, 12-week experiment, groups received one of two dietary treatments: control (CON) total mixed ration (TMR) (108.2 ± 66.1 DIM) or TMR supplemented with microalgae (AURA) at 6.0 g/kg dry matter intake (DMI) (104.4 ± 54.6 DIM). The microalgae (AURA) product was provided by Alltech Inc. (ALL-G-RICH®, Nicholasville, KY, USA) and consisted of a heterotrophically grown, unextracted Aurantiochytrium limacinum (CCAP 4087/2), with a guaranteed minimum of 160 mg DHA/g and not more than 0.3% sodium.
The TMR (as-fed basis) contained corn silage (24.0 kg), concentrate + cotton seed mix (70:30; 5.5 kg), corn meal + barley flake mix (60:40; 1.5 kg), corn meal + sorghum meal mix (70:30; 4.5 kg), water (7.0 kg), ryegrass hay (1.8 kg), and dehydrated alfalfa hay (4.5 kg). The composition of the concentrate was as follows: soy protein, wheat bran, dehulled sunflower meal (34%), limestone, sodium bicarbonate, salt, corn meal, cane molasses, dicalcium phosphate, magnesium oxide, soluble molasses concentrate and corn germ meal. Moreover, each kg of concentrate contained vitamin A (47,640 IU), vitamin D3 (4,368 IU), vitamin E (85.68 mg), Ca (13.47 mg), P (6.02 mg), Mg (4.45 mg), Na (12.65 mg), Cu (47.35 mg), Fe (261.16 mg), Zn (135.90 mg), I (1.55 mg), Co (16.67 mg), Mn (138.65 mg), and Se (0.47 mg). AURA was pre-diluted with corn meal 50:50 and each cow received either 300 g corn meal (CON) or the AURA: corn meal mixture as top-dressing in the morning during the TMR administration. Cows were housed at the CERZOO Research Centre farm (Piacenza, Italy) in three pens with six cows per treatment. Housing management, feeding, and husbandry conditions were considered representative of modern, commercial, European dairy operations.
Sampling, measurement and analyses
The analytical composition of AURA was determined prior to the start of the study: crude protein (AOAC 990.03), crude fat (AOAC 954.02), fatty acid composition (AOAC 996.06), moisture (AOAC 930.15) and ash (AOAC 942.05). The nutrient composition of fresh TMR samples were analysed four times (every 28 days): crude protein (ISO 5983-1), ADF (ISO 13906), NDF (ISO 16472), starch (ISO 10520:1997E), crude fat (ISO 6492) and predicted metabolisable energy (Gallo et al., Reference Gallo, Moschini, Cerioli and Masoero2013). Dry matter was calculated weekly by force drying TMR samples at 103°C to a constant weight (ISO 6496). DHA in the corn meal: AURA mixture was quantified following fat extraction by the Folch method (Folch et al., Reference Folch, Lees and Sloane Stanley1957), esterification of extracted fat, separation of individual fatty acid methyl esters by gas chromatography and quantification against known standards (Bannon et al. Reference Bannon, Craske and Hiliker1985).
Performance data were collected as follows: live weight (per cow daily), TMR intake (per pen daily), and body condition score (BCS) (per cow weekly). Daily milk production (per cow) was reported as the sum of morning and evening milk yields. Milk was sampled from each cow in the morning and afternoon of d 0, 7, 14, 21, 28, 56, and 84. The morning and afternoon samples were combined for each cow, according to their milk production, and divided into two aliquots of not less than 50 ml each. The first aliquot was analysed for milk components (i.e., fat, protein, lactose, somatic cell, and urea). Fat corrected milk (FCM, 4%) was calculated as per Gaine's formula (1923): FCM (kg) = 0.4M + 15 F where: M = milk yield (kg). F = M x fat content (%). The second milk aliquot was analysed for individual fatty acid composition following an in situ preparation of fatty acid methyl esters, separation by gas chromatography and quantification against known standards (Park and Goins, Reference Park and Goins1994, Loor and Herbein, Reference Loor and Herbein2001). Milk fatty acid (FA) content was assumed to be 93.3% of total milk fat (Glasser et al., Reference Glasser, Doreau, Ferlay and Chilliard2007; Moates et al., 2013). DHA in milk yield (g/d) was calculated as:
 $$\eqalign{& {\rm 0}{\rm. 933} \times \hbox{mean daily milk fat yield} \left( {{\rm 100g/day}} \right) \cr & \quad {\rm} \times \hbox{DHA concentration in milk FA} \left( {{\rm g / 100g FA}} \right).} $$
$$\eqalign{& {\rm 0}{\rm. 933} \times \hbox{mean daily milk fat yield} \left( {{\rm 100g/day}} \right) \cr & \quad {\rm} \times \hbox{DHA concentration in milk FA} \left( {{\rm g / 100g FA}} \right).} $$
DHA transfer efficiency (%) from diet to milk was calculated (Moates et al., 2013) as:
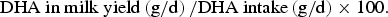 $$\hbox{DHA in milk yield} \left( {{\rm g/d}} \right) / \hbox{DHA intake} \left( {{\rm g/d}} \right) \times 100.$$
$$\hbox{DHA in milk yield} \left( {{\rm g/d}} \right) / \hbox{DHA intake} \left( {{\rm g/d}} \right) \times 100.$$
Statistical analyses
All statistical analyses were performed using SAS 9.3 software (SAS Institute Inc., 2011). Milk yield, fat content and DHA content were analysed using linear mixed models for repeated measures using proc MIXED procedure. For each the model included the fixed effects for treatment (CON vs AURA), day, and their interactions; for fat content and DHA, the model also included the random effect of cows. For milk yield, each model was subjected to two covariance structures (i.e., compound symmetry and autoregressive); the model with the smallest Akaike information criterion was then used. Because of the irregular sampling intervals used to determine milk fat and DHA content each of these models was subjected to compound symmetric and spatial power covariance structure. Two-sample t-tests were used to determine between-treatment differences. Mean body condition score was compared between treatments using the Mann Whitney U test (Sawilowsky, Reference Sawilowsky, Salkind and Rasmussen2007).
Results and discussion
Ingredient and diet analyses
The microalgae, AURA, used in the study primarily consisted of 66.9 g crude fat /100 g DM biomass composed of a significant level of palmitic acid and docosahexaenoic acid (DHA), 36.66 g and 16.12 g / 100 g DM biomass respectively. Additionally, AURA contained 12% crude protein, 3.2% ash and 2.2% moisture. No presence of conjugated linoleic acid (C18:2 c9, t11) was found in the fatty acid analysis of the test article.
The raw materials were analysed for each new lot of production of TMR during the study. The analytical characteristics of the raw materials (% of dry matter) used in the preparation of the TMR and the analysis of the complete TMR are provided in Table 1.
Table 1. Analytical characteristics (% of dry matter) of the raw materials used in TMR preparation and the complete TMR mix

Note: 1 Produced in the facility, 2 Provided by Consorzio Agrario of Cremona (CR, Italy), 3 Provided by Consorzio Agrario of Piacenza (PC, Italy); 4 Provided by ALIVERDE (RA, Italy)
Cow health and performance
Cows maintained good health status throughout the study and no veterinary treatments were required. Body condition score, an indicator of cow energy status, was similar between treatments over the 12-week experiment (Table 2). These results are consistent with other studies that showed dairy cow BCS was unaffected by microalgae supplementation (Franklin et al., Reference Franklin, Martin, Baer, Schingoethe and Hippen1999, Glover et al., Reference Glover, Budge, Rose, Rupasinghe, MacLaren, Green-Johnson II and Fredeen2012). All SCC counts were less than 400,000 CFU/ml of milk, the regulatory limit in Italy.
Table 2. Health and performance indicators for mid-lactation cows fed a control (CON) diet or CON supplemented with a docosahexaenoic acid (DHA) rich microalgae

SCC = somatic cell count
1 Cows were fed a control (CON) total mixed ration or TMR supplemented with unextracted Aurantiochytrium limacinum algae (AURA, Alltech Inc.) at 6 g/kg DM. 2 Standard error of the mean for n = 18. 3 Body condition score: 1 = Emaciated; 2 = Thin; 3 = Moderate; 4 = Stout; 5 = Obese. 4 Mean of day 7–84, data analysed using an ANOVA with repeated measures. 5 Mean of day 1–84 data analysed using an ANOVA with repeated measures. 6 Daily TMR intake on a dry matter (DM) basis
Average body weight gain of cows fed AURA was numerically higher (+10.7 kg) compared with CON (P = 0.592) (Table 2). Dry matter intake was unaffected by treatment in agreement with a study involving dairy cow supplementation with algal oil (Stamey et al., Reference Stamey, Shepherd, de Veth and Corl2012), but contrary to reports of dry matter intake depression in dairy cows in response to algae meal (Franklin et al., Reference Franklin, Martin, Baer, Schingoethe and Hippen1999, Moate et al., Reference Moate, Williams, Hannah, Eckard, Auldist, Ribaux, Jacobs and Wales2013) or fat-protected algae (Glover et al., Reference Glover, Budge, Rose, Rupasinghe, MacLaren, Green-Johnson II and Fredeen2012) and in ewes in response to algal oil supplementation (Reynolds et al., Reference Reynolds, Cannon and Loerch2006).
Milk yield
At the start of the experiment, milk yield was similar between treatment groups. As the study progressed, cows in the AURA group had slightly greater yield. This difference became most pronounced in the final three weeks of the experiment (Table 3). For the total 12-week feeding period there was a trend in productivity, with milk yield 1.9 kg/cow/d (5.4%) greater in cows that consumed the AURA (37.3 versus 35.4 kg/cow/d; P = 0.0948). Previous investigators have reported that dairy cow milk yield was unaffected by algae biomass or algal oil supplementation (Stamey et al., Reference Stamey, Shepherd, de Veth and Corl2012) or whole algal cell supplementation (Moate et al., Reference Moate, Williams, Hannah, Eckard, Auldist, Ribaux, Jacobs and Wales2013), however, these studies were restricted to supplementation periods of much shorter duration (i.e. 7 and 16 d, respectively). These data may suggest that feeding AURA over an extended period may lead to better persistency in the lactation curve when fed around mid–lactation; this needs to be verified in further studies.
Table 3. Milk production from mid-lactation cows fed a control (CON) diet or CON supplemented with a docosahexaenoic acid (DHA) rich microalgae

1 Cows were fed a control (CON) total mixed ration or TMR supplemented with unextracted Aurantiochytrium limacinum algae (AURA, Alltech Inc.) at 6 g/kg DM. 2 Data analysed using an ANOVA with repeated measures for each period and the entire study (days 1–84). 3 Standard error of the mean for n = 18. a,b Values within a row with different superscripts differ significantly at P < 0.10.
Milk fat
The AURA treated cows consistently produced milk with lower fat content (3.19%) compared with milk from CON cows (3.66%) (Table 4), a reduction of 12%. Fat production was also lower (P = 0.0538) in milk from the AURA treated cows, but the 4% fat corrected milk production did not appreciably decline. These findings are consistent with those of previous investigators (Franklin et al., Reference Franklin, Martin, Baer, Schingoethe and Hippen1999, Boeckaert et al., Reference Boeckaert, Vlaeminck, JDijkstra, Issa-Zacharia, Van Nespen, Can Straalen and Fievez2008b, Moate et al., Reference Moate, Williams, Hannah, Eckard, Auldist, Ribaux, Jacobs and Wales2013) who reported milk fat depression in response to algae supplementation of dairy cow rations. Conjugated linoleic acid biohydrogenation intermediates, especially the CLA c12t10 isomer, have been implicated in inhibiting mammary lipid metabolism (Shingfield et al., Reference Shingfield, Bonnet and Scollan2013) resulting in milk fat depression. Although the CLA isomer profile underwent changes in the current study, the CLA c12t10 isomer was not measured so we are unable to explain the milk fat depression observed based on our fatty acid profile data.
Table 4. Components and component output of milk from mid-lactation cows fed a control (CON) diet or CON supplemented with a docosahexaenoic acid (DHA) rich microalgae

1 Cows were fed a control (CON) total mixed ration or TMR supplemented with unextracted Aurantiochytrium limacinum algae (AURA, Alltech Inc.) at 6 g/kg DM. 2 Mean for days 7–84, data analysed as an ANOVA with repeated measures. 3 Standard error of the mean for n = 18. a,b Values within a row with different superscripts differ significantly at P < 0.10.
Milk protein, lactose and urea
Milk protein content (%) and protein production (kg/d) were unaffected by AURA supplementation (Table 4). Other investigators have similarly reported no effects on milk protein in response to supplementation with algae biomass, algal oil (Stamey et al., Reference Stamey, Shepherd, de Veth and Corl2012) or algae meal (Moate et al., Reference Moate, Williams, Hannah, Eckard, Auldist, Ribaux, Jacobs and Wales2013). Franklin and colleagues (1999) reported a tendency (P = 0.08) for the percentage of protein in milk to be lower in cows supplemented with algae, but that the total protein yield remained unaffected (P < 0.53) (Franklin et al., Reference Franklin, Martin, Baer, Schingoethe and Hippen1999). In the current study, lactose content and total production and urea content were unaffected by AURA treatment (Table 4).
Milk fatty acid profile
Supplementation with AURA significantly altered the fatty acid profile of milk (Table 5) to increase the percentages of unsaturated fatty acids (P = 0.0011), PUFAs (P = 0.0001), omega-3 fatty acids (P = 0.0001), DHA (P = 0.0001), and conjugated linoleic acid CLA (C18:2 c9, t11) (P = 0.0001). The omega-3 : omega-6 fatty acid ratio also increased from 0.20 to 0.35 (% of sum (Σ) total fatty acid) (P = 0.0001). Concomitant reductions occurred in the percentages of saturated fatty acids (P= 0.0011), γ-linolenic acid (C18:3n6) (P = 0.0001), oleic acid (P = 0.0287), stearic acid (P = 0.0653), palmitic acid (P = 0.0024), and undecanoic acid (P = 0.0095). These alterations are generally consistent with those of other investigators (Franklin et al., Reference Franklin, Martin, Baer, Schingoethe and Hippen1999, Boeckaert et al., Reference Boeckaert, Vlaeminck, JDijkstra, Issa-Zacharia, Van Nespen, Can Straalen and Fievez2008b, Stamey et al., Reference Stamey, Shepherd, de Veth and Corl2012, Moate et al., Reference Moate, Williams, Hannah, Eckard, Auldist, Ribaux, Jacobs and Wales2013).
Table 5. Repeated measures for fatty acid composition of milk (% of the Σ) from mid-lactation cows fed a control (CON) diet or CON supplemented with a docosahexaenoic acid (DHA) rich microalgae

1 Cows were fed a control (CON) total mixed ration or TMR supplemented with unextracted Aurantiochytrium limacinum algae (AURA, Alltech Inc.) 6 g/kg DM. 2 Mean for days 7–84, data analysed using an ANOVA with repeated measures. 3 Standard error of the mean for n = 18. 4 Ratio of omega 3 / omega 6 concentrations of total fatty acid content of milk
DHA was not found in the unsupplemented CON milk samples at any time point in the study. Incorporation of DHA in milk increased (P < 0.0001) over the course of the experiment, increasing from the start of trial until about d 28, where the concentration plateaued until the end of the study period, d 84 (Figure 1). AURA supplementation enriched (P = 0.0001) milk fat in DHA (% of sum (Σ) total fatty acid) to 0.37 ± 0.06% and 0.46 ± 0.020% following d 7–84 and d 28–84 days of continuous feeding respectively. Therefore, supplementation of dairy cows with approximately 146 g AURA/h/d under the conditions of this trial, for a minimum of 28 days, resulted in 13 mg DHA per 100 ml milk. A pattern of DHA concentration rise and plateau, in response to dietary enrichment using a rumen protected algae, was similarly observed by investigators in a six week trial (Franklin et al., Reference Franklin, Martin, Baer, Schingoethe and Hippen1999).

Figure 1. Temporal pattern of docosahexaenoic acid (DHA) incorporation (% of total) and transfer efficiency (%) into milk fatty acids in response to AURA supplementation of dairy cow TMR
In cows fed the AURA treatment there was a transfer of DHA from algae to milk with an efficiency of 18.1% (d 7–84; Table 6). A broad range of transfer efficiencies have been reported in the literature, from 1.0% - 16.7% (Franklin et al., Reference Franklin, Martin, Baer, Schingoethe and Hippen1999; Chilliard et al., Reference Chilliard, Feray and Doreau2001; Boeckaert et al., 2008; Stamey et al., Reference Stamey, Shepherd, de Veth and Corl2012; Moate et al., Reference Moate, Williams, Hannah, Eckard, Auldist, Ribaux, Jacobs and Wales2013).
Table 6. Transfer efficiency (%) of docosahexaenoic acid (DHA) to milk from feeding AURA to dairy cows

1 DHA in milk yield (g/d) = 0.933 x mean daily milk fat yield (100 g/day) x DHA concentration in milk fatty acid (g/100 g fatty acid). 2 TMR intake based on previous weekly average intake. 3 DHA intake based on 161.2 mg DHA/g AURA = 0.966 g DHA/ kg TMR DMI. 4 DHA transfer efficiency (%) from diet to milk = DHA in milk yield (g/d) / DHA intake (g/d) x 100. 5 Mean of d 7–84, data analysed using an ANOVA with repeated measures
Stamey and colleagues reported DHA transfer efficiencies from algae to milk fat ranging from 1.0 to 3.4% over a seven day feeding period and hypothesised that by feeding earlier in lactation the transfer efficiency may have improved (Stamey et al., Reference Stamey, Shepherd, de Veth and Corl2012). However, the cows on this current study were all in mid- to late- lactation and yet had a very high transfer efficiency. Albeit, the longer feeding period in the current study played a significant role in the increase in DHA yield in milk (Figure 1).
Two independent studies reported transfer efficiencies of 8.4% and 8.9% when feeding unprotected sources of algal biomass to dairy cows (Franklin et al., Reference Franklin, Martin, Baer, Schingoethe and Hippen1999; Moate et al., Reference Moate, Williams, Hannah, Eckard, Auldist, Ribaux, Jacobs and Wales2013). However, the DHA transfer efficiency found in the current study compared similarly with the transfer efficiency when feeding a protected algal biomass as reported by Franklin and colleagues (1999); 18.1 versus 16.7% respectively. This transfer efficiency exceeds those reported for the transfer of DHA from fish oil to milk fat, which has been reported as generally having a transfer efficiency of < 4% (Chilliard et al., Reference Chilliard, Feray and Doreau2001, Lock and Bauman, Reference Lock and Bauman2004, Palmquist, Reference Palmquist2009). The current study used an unextracted algal biomass which was produced under heterotrophic and low sodium conditions (non-marine) which may allow a degree of protection to the algal cell membrane during drying. Apajalahti et al. (unpublished) found in rumen simulation tests that the algal cell protects the DHA with no significant loss to leaching and oxidation over eight hours. This may explain the low degree of biohydrogenation products in the rumen and high transfer efficiency of DHA in this study.
A significant increase in the CLA content was observed in milk of cows receiving the AURA supplement compared to cows receiving the control diet, 0.86 versus 0.34% of sum of total fatty acid, respectively (P = 0.0001). The AURA supplement contained no CLA. However, the DHA content of the algae is thought to promote the accumulation of vaccenic acid in the rumen by inhibiting C18 biohydrogenation (Chow et al., Reference Chow, Fievez, Moloney, Raes, Demeyer and De Smet2004, Boeckaert et al., Reference Boeckaert, Vlaeminck, JDijkstra, Issa-Zacharia, Van Nespen, Can Straalen and Fievez2008b). Vaccenic acid is the primary of precursor for CLA synthesis in the mammary gland via Δ9-desaturase (Griinari et al., Reference Griinari, Corl, Lacy, Chouinard, Nurmela and Bauman2000, AbuGhazaleh and Jenkins, Reference AbuGhazaleh and Jenkins2004, Mosley et al., Reference Mosley, Shafii, Moate and McGuire2006, AbuGhazaleh et al., Reference AbuGhazaleh, Potu and Ibrahim2009). In the current study, vaccenic acid increased (P = 0.0001) from 1.99 to 7.03% in the fatty acid profile in response to the AURA supplementation, possibly explaining the observed increase (P = 0.0001) in CLA in milk from AURA-treated cows. This increase is consistent with increases in CLA (and C18:1 isomers) reported by other investigators (Franklin et al., Reference Franklin, Martin, Baer, Schingoethe and Hippen1999, Boeckaert et al., Reference Boeckaert, Vlaeminck, JDijkstra, Issa-Zacharia, Van Nespen, Can Straalen and Fievez2008b, Stamey et al., Reference Stamey, Shepherd, de Veth and Corl2012, Moate et al., Reference Moate, Williams, Hannah, Eckard, Auldist, Ribaux, Jacobs and Wales2013). The secondary enhancement of CLA potentially confers additional health benefits to DHA-enriched milk, since CLA has been shown to inhibit carcinogenesis (Kelley et al., Reference Kelley, Hubbard and Erickson2007, Amaru and Field, Reference Amaru and Field2009, Donnelly et al., Reference Donnelly, Olsen, Lewis, Eisenberg, Easstman and Kinlaw2009) and to increase lean body mass in humans (Steck et al., Reference Steck, Chalecki, Miller, Conway, Austin, Hardin, Albright and Thuillier2007).
Conclusions
Supplementing dairy cow TMR with the algae treatment ALL-G-RICH® (AURA) at levels of 6 g/kg DMI for 12 weeks altered the fatty acid profile of milk compared with milk from unsupplemented cows such that the proportion of unsaturated fatty acids increased and the proportion of saturated fatty acid content declined. Omega-3 fatty acid content increased and was accompanied by a favourable increase in the omega-3: omega-6 fatty acid ratio. A high transfer efficiency of DHA from feed to milk was observed and hypothesised to be related to the manufacturing method of the heterotrophic algal biomass.
Milk yield tended to be greater (+5.4%) in cows fed the algae supplement, whereas milk fat content and fat production significantly declined without a significant change in (4%) fat corrected milk. Together these results indicate that supplementing dairy cow diet with DHA-rich microalgae is a feasible means for creating DHA-enriched milk for human consumption.
Acknowledgements
The authors would like to express their gratitude to Mrs. Sylvie Andrieu for monitoring the study, Ms. Rebecca Timmons for her technical input on microalgae and to Dr. Helen Warren for her useful discussions during the manuscript writing phase.
Declaration of Interest
Funding for this work was provided by Alltech SARL (France). The author C.A. Moran is an employee of Alltech SARL, a subsidiary of Alltech Inc., which produces and markets ALL-G-RICH®, the commercial product used in this study.








 Open access
Open access