



Introduction
A stable climate is vitally important for life on Earth. However, the Earth's climate is now facing rapid changes with profound effects on the environment and its inhabitants. Specifically, melting of the polar ice caps and glaciers is responsible for rising sea levels (Raper and Braithwaite, Reference Raper and Braithwaite2006; Paul, Reference Paul2011), while heatwaves and droughts are becoming more severe and frequent in more and more regions of the globe (Olesen et al., Reference Olesen, Trnka, Kersebaum, Skjelvåg, Seguin, Peltonen-Sainio, Rossi, Kozyra and Micale2011; Trenberth, Reference Trenberth2018). All these changes have major consequences for human activities and the environment (Chen et al., Reference Chen, McCarl and Chang2012; Trenberth, Reference Trenberth2018). Furthermore, the increased risk to agricultural production seems to be one of the most important consequences of climate change (Cline, Reference Cline2007; Evangelista et al., Reference Evangelista, Young and Burnett2013), while the distribution of several wild plant species is expected to change (McDonald et al., Reference McDonald, Riha, DiTommaso and DeGaetano2009; Castellanos-Frías et al., Reference Castellanos-Frías, Garcia De León, Bastida and Gonzalez-Andujar2016; Reif et al., Reference Reif, Xystrakis, Gärtner and Sayer2017). On the contrary, it is expected to have a positive effect on agricultural production in high-latitude regions such as Russia and Canada, while its effects on other areas of the world are debatable (Lewis and Witham, Reference Lewis and Witham2012). The overall global effects of climate change on agriculture will be negative, in terms of food security and stability (Nelson et al., Reference Nelson, Rosegrant, Koo, Robertson, Sulser, Zhu, Ringler, Msangi, Palazzo, Batka, Magalhaes, Valmonte-Santos, Ewing and Lee2009; Wheeler and Von Braun, Reference Wheeler and Von Braun2013), while the effects are expected to be worse in countries already facing food shortages (Wheeler and Von Braun, Reference Wheeler and Von Braun2013). Recent data have revealed that the number of undernourished people increased to 815 million in 2016, representing about 11% of the global population (FAO et al., 2017); according to Dawson et al. (Reference Dawson, Perryman and Osborne2016), 31% of the global population is expected to face the risk of undernourishment by 2050.
In Southern Europe, climate change is predicted to result in temperature elevation and an increase in the frequency of drought (Lehner et al., Reference Lehner, Döll, Alcamo, Henrichs and Kaspar2006; Lovelli et al., Reference Lovelli, Perniola, Scalcione, Troccoli and Ziska2012). In the same region, climate change will also cause declines in crop yields (Debaeke et al., Reference Debaeke, Casadebaig, Flenet and Langlade2017; Gammans et al., Reference Gammans, Mérel and Ortiz-Bobea2017); thus, restructuring of crops, as well as changes in cultivation practices, have been suggested as alternative strategies to solve this problem (Olesen et al., Reference Olesen, Trnka, Kersebaum, Skjelvåg, Seguin, Peltonen-Sainio, Rossi, Kozyra and Micale2011; Nastis et al., Reference Nastis, Michailidis and Chatzitheodoridis2012). Here, it is important to point out that vegetables, pulses and cereals are more vulnerable to climate change (e.g. increased temperatures, drought incidence) than other arable crops (Acikgoz, Reference Acikgoz2011; Bahl, Reference Bahl2015; Gammans et al., Reference Gammans, Mérel and Ortiz-Bobea2017). A recent study by Gammans et al. (Reference Gammans, Mérel and Ortiz-Bobea2017) predicted yield losses of up to 21.0% for winter wheat, 17.3% for winter barley and 33.6% for spring barley due to yearly deviations from climate averages for temperature and precipitation.
Climate change is already occurring in Europe and it is anticipated that significant changes in weed flora will take place throughout Europe in the near future (Peters et al., Reference Peters, Breitsameter and Gerowitt2014; Castellanos-Frías et al., Reference Castellanos-Frías, Garcia De León, Bastida and Gonzalez-Andujar2016). Enhanced growth of C4 weeds (the species that use the C4 carbon fixation pathway [Hyvönen, Reference Hyvönen2011; Rodenburg et al., Reference Rodenburg, Meinke and Johnson2011]), spreading of perennial weeds such as Solanum elaeagnifolium Cav. (silverleaf nightshade) and Sorghum halepense (L.) Pers. (Johnsongrass) (Mekki, Reference Mekki2007; Leguizamón and Acciaresi, Reference Leguizamón and Acciaresi2014) and invasion of new species (Clements and DiTommaso, Reference Clements and DiTommaso2012) are some of the effects of climate change (e.g. increased temperatures, drought) on weed flora in Europe and other parts of the world. Understanding crop–weed interference in response to climate change is also a major challenge, as weeds remain the most important limiting factor in economically viable crop production (Ziska and Dukes, Reference Ziska and Dukes2011; Karkanis et al., Reference Karkanis, Bilalis, Efthimiadou and Katsenios2012). According to Lovelli et al. (Reference Lovelli, Perniola, Scalcione, Troccoli and Ziska2012), weeds’ competitiveness is already increasing as a result of elevated temperatures in the Mediterranean region. From this perspective, two critical questions need to be addressed (Neve et al., Reference Neve, Vila-Aiub and Roux2009):
• Are weeds capable of adapting to a changing environment?
• What effect does climate change have on vegetable–weed competition?
Taking the above questions into consideration, the current review aims to provide an up-to-date overview of the adaptation mechanisms in weed species commonly found in Southern Europe in order to expand the available knowledge regarding the adaptability of these weeds to drought and elevated temperatures. Emphasis is also placed on revealing the effects of drought and increased temperatures on vegetable–weed competition, and most importantly, its effect on crop yield, as well as on weed distribution and population composition in this region.
Weeds, drought and high temperatures
Plant response to drought is a complex biological process involving different mechanisms of defence (Karkanis and Petropoulos, 2017; Plesa et al., Reference Plesa, González-Orenga, Al Hassan, Sestras, Vicente, Prohens, Sestras and Boscaiu2018). Therefore, several weed species possess a wide variety of developmental and physiological mechanisms that enable them to adapt to increased temperatures and the limited availability of water (Fig. 1). In this section, a description is presented of the most important adaptation mechanisms, while emphasis has been placed on weed species commonly found in regions where vegetable crops are cultivated in Southern Europe.
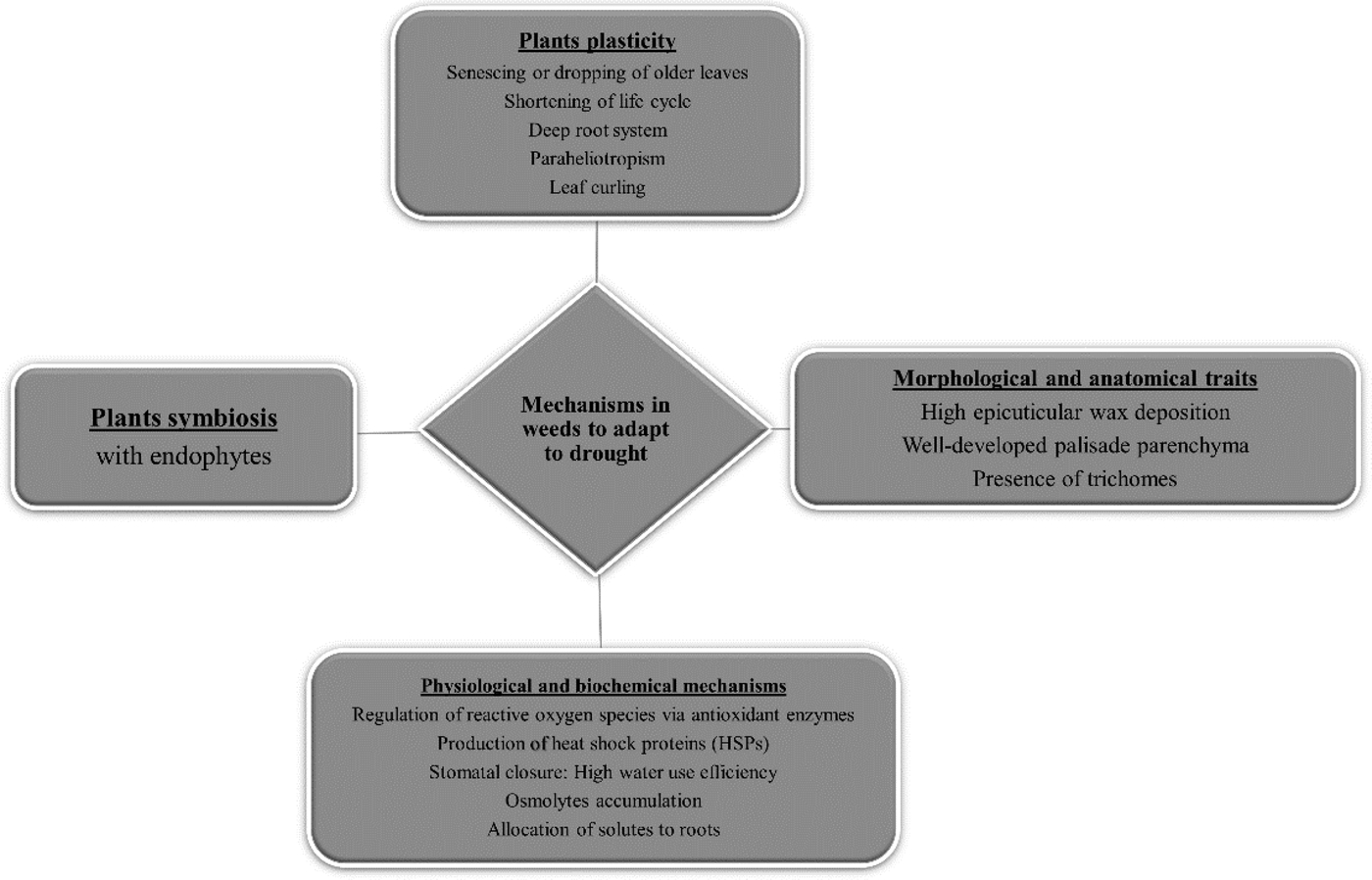
Fig. 1. Different mechanisms in weeds to adapt to drought and high temperatures.
Weed plasticity and drought
Several weed species decrease their water requirements by reducing the total plant leaf area. According to Schmidt et al. (Reference Schmidt, Blankenship and Lindquist2011), Abutilon theophrasti Medicus (velvetleaf) responds to water stress by senescing its oldest leaves, allowing the young leaves to support plant development and seed production. Similarly, Ward et al. (Reference Ward, Tissue, Thomas and Strain1999) observed that A. theophrasti reacts to drought by senescing and reducing the leaf area, allowing the remaining leaves to maintain high leaf water potential.
Plants growing at high temperatures adapt to heat stress by reducing the absorption of solar radiation either by growing hairs, which form a thick layer on leaves’ surfaces, or by rolling their leaves (Hasanuzzaman et al., Reference Hasanuzzaman, Nahar, Alam, Roychowdhury and Fujita2013). In addition, Karkanis et al. (Reference Karkanis, Bilalis and Efthimiadou2011) reported that A. theophrasti plants have the ability to adjust their temperature by orienting their leaf blades parallel to incident sunlight (Fig. 2).

Fig. 2. (Colour online) Mechanism in Abutilon theophrasti plants to adapt to water stress.
Furthermore, several weeds respond to drought by shortening their life-cycles. Indeed, A. theophrasti plants suffering from drought were observed to flower earlier compared with well-watered plants (Karkanis et al., Reference Karkanis, Bilalis and Efthimiadou2011). Such an early onset of reproduction in plants in response to water stress has been reported by Volis et al. (Reference Volis, Verhoeven, Mendlinger and Ward2004) and Volis (Reference Volis2009) for Hordeum spontaneum Koch (wild barley) and Avena sterilis (wild oat) as well.
High root/shoot ratios also contribute to drought tolerance in plants (Heschel et al., Reference Heschel, Sultan, Glover and Sloan2004; Xu et al., Reference Xu, Li, Shan, Ma, Ichizen and Huang2006). Heschel et al. (Reference Heschel, Sultan, Glover and Sloan2004) evaluated three populations of Polygonum persicaria L. (spotted ladysthumb) for their adaptation mechanisms to drought: the results of that study revealed that all populations increased their water use efficiency (WUE) and root biomass allocation under drought conditions. Similarly, drought resistance in S. elaeagnifolium is associated with its deep root system, which consists of creeping horizontal and deep vertical roots (Mekki, Reference Mekki2007). Travlos (Reference Travlos2013) similarly observed that S. elaeagnifolium plants exhibited a high root/shoot ratio under water stress. Furthermore, Zhu et al. (Reference Zhu, Wu, Stanton, Burrows, Lemerle and Raman2013) reported considerable morphological variations between plants collected from several regions with different rainfall levels. According to Zhu et al. (Reference Zhu, Wu, Stanton, Burrows, Lemerle and Raman2013), S. elaeagnifolium plants originating from areas with high rainfall levels had greater heights and larger leaves compared with those from regions with low rainfall levels, indicating changes in plant morphology as an adaptation mechanism to drought stress. Similar to S. elaeagnifolium, Convolvulus arvensis L. (field bindweed), which is considered to be a drought-tolerant species, develops a deep root system in order to overcome limited water availability (Sosnoskie and Hanson, Reference Sosnoskie and Hanson2016). The increase of root/shoot ratio in rice plants under drought-stress conditions has been associated with higher accumulation of dry matter and soluble sugars in roots as a response to higher activities of sucrose-phosphate synthase in shoots and invertase in roots, thus a higher transportation rate of sucrose from shoots to roots (Xu et al., Reference Xu, Cui, Xu, Nie, Huang and Peng2015). This function is a plant defence mechanism to sustain root growth by decreasing allocation of metabolites to shoots (Gargallo-Garriga et al., Reference Gargallo-Garriga, Sardans, Pérez-Trujillo, Rivas-Ubach, Oravec, Vecerova, Urban, Jentsch, Kreyling, Beierkuhnlein, Parella and Peñuelas2014). Moreover, these changes in root architecture and growth allow plants to increase water uptake from deeper soil layers and overcome the negative effects of drought stress, although this response is highly dependent on genotype, as well as severity and duration of exposure to stress conditions (Lemoine et al., Reference Lemoine, La Camera, Attanassova, Dédaldéchamp, Allario, Pourtau, Bonnemain, Laloi, Coutos-Thévenot, Maurousset, Faucher, Girousse, Lemonnier, Parrilla and Durand2013; Xu et al., Reference Xu, Cui, Xu, Nie, Huang and Peng2015; Sosnoskie and Hanson, Reference Sosnoskie and Hanson2016).
Drought resistance of weeds is also associated with a variety of anatomical traits that enable them to minimize water loss. For instance, Hatterman-Valenti et al. (Reference Hatterman-Valenti, Pitty and Owen2011) observed that A. theophrasti plants grown under drought conditions had greater leaf epicuticular wax deposition as compared with leaves of well-watered plants. Ecballium elaterium (L.) A. Rich (squirting cucumber) is also considered to be drought-resistant because of its leaf structure. According to Christodoulakis et al. (Reference Christodoulakis, Kollia and Fasseas2011), E. elaterium plants present a well-developed palisade parenchyma in comparison with spongy parenchyma, which is relatively compact, while numerous multicellular trichomes exist on both leaf surfaces. High palisade to spongy parenchyma ratio is considered to be a xeromorphic trait (Christodoulakis et al., Reference Christodoulakis, Kollia and Fasseas2011). Christodoulakis et al. (Reference Christodoulakis, Lampri and Fasseas2009) reported that the leaves of S. elaeagnifolium have four layers of palisade cells, while spongy parenchyma is absent from the mesophyll of its leaves. Similar to E. elaterium plants, numerous multicellular trichomes are also present on both leaf surfaces (Christodoulakis et al., Reference Christodoulakis, Lampri and Fasseas2009). The hairs on leaf surfaces are advantageous under drought conditions as they reduce the absorption of solar radiation, in addition to helping prevent water loss through the stomata (Hasanuzzaman et al., Reference Hasanuzzaman, Nahar, Alam, Roychowdhury and Fujita2013).
Physiological and biochemical adaptations to drought
Weeds possess a variety of physiological mechanisms that enable them to adapt to water stress. For instance, under drought conditions, A. theophrasti plants retain water through stomatal closure (Schmidt et al., Reference Schmidt, Blankenship and Lindquist2011). For the same species, Karkanis et al. (Reference Karkanis, Bilalis and Efthimiadou2011) reported that water stress resulted in lower stomatal conductance. Amaranthus retroflexus L. (redroot bigweed) is similarly considered to be a drought-tolerant weed, commonly found in Southern Europe. According to Lovelli et al. (Reference Lovelli, Perniola, Ferrara, Amato and Di Tommaso2010a), the invasiveness of A. retroflexus increases under drought conditions because of its capacity to maintain a high WUE compared with other less-resistant crop and weed species.
Under drought conditions, an increased allocation of solutes in the roots of Rumex obtusifolius L. Rumob. (broadleaf dock) compared with its aerial parts has been reported by Gilgen and Feller (Reference Gilgen and Feller2013). The allocation of solutes to the roots helps R. obtusifolius plants to recover quickly from drought stress; this rapid recovery is a key factor for overall plant performance and competitiveness against other species, especially under stress (Gilgen and Feller, Reference Gilgen and Feller2013, Reference Gilgen and Feller2014).
Drought stress also affects plant photosynthesis, either through stomatal closure that impairs carbon dioxide (CO2) diffusion or by inducing oxidative stress (Chaves et al., Reference Chaves, Flexas and Pinheiro2009). Under these conditions, over-production of reactive oxygen species (ROS) causes oxidative damage in the cell (Shigeoka et al., Reference Shigeoka, Ishikawa, Tamoi, Miyagawa, Takeda, Yabuta and Yoshimura2002), while several enzymes that play important roles in the metabolism of ROS, such as superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase, monodehydroascorbate reductase, dehydroascorbate reductase, glutathione reductase (GR), glutathione S-transferase, glutathione peroxidase (GPX) and peroxidases (POD), show lower activity (Shigeoka et al., Reference Shigeoka, Ishikawa, Tamoi, Miyagawa, Takeda, Yabuta and Yoshimura2002; Hasanuzzaman et al., Reference Hasanuzzaman, Hossain, da Silva, Fujita, Venkateswarlu, Shanker, Shanker and Maheswari2012). In this regard, Pandey et al. (Reference Pandey, Baig, Chandra and Bhatt2010) reported that A. sterilis plants exhibit drought-stress tolerance by displaying a high POD activity.
Cynodon dactylon (L.) Pers. (Bermuda grass) is a perennial weed listed as one of the ten most notorious weeds in the world, although it also used as a turf grass for landscaping purposes (Holm et al., Reference Holm, Plucknett, Pancho and Herberger1997; Shi et al., Reference Shi, Wang, Cheng, Ye and Chan2012). According to Shi et al. (Reference Shi, Wang, Cheng, Ye and Chan2012), C. dactylon plants display a range of mechanisms to withstand drought stress, including the control of water loss from leaves, accumulation of osmolytes (proline and soluble sugars) and the regulation of ROS via antioxidant enzymes (SOD, CAT, POD, GR and GPX). Production of molecular chaperones (i.e. heat shock protein 70; HSP70) by C. dactylon plants in response to drought has been reported by Shi et al. (Reference Shi, Ye and Chan2014) and Ye et al. (Reference Ye, Shi, Wang and Chan2015). Moreover, it is important to highlight a particular mechanism of drought resistance here, which is observed in Portulaca oleracea L. plants (common purslane). This species uses the C4 carbon fixation pathway, but under conditions of water stress shifts its photosynthetic process to crassulacean acid metabolism, which allows the plants to close their stomata during the day and open them at night in order to retain water (Lara et al., Reference Lara, Disante, Podestá, Andreo and Drincovich2003, Reference Lara, Drincovich and Andreo2004).
Drought and endophytes
Several winter grass weeds, such as Briza, Bromus and Poa, are infected by fungal symbionts, Epichloë/Neotyphodium endophytes, which grow asymptomatically in the intercellular spaces of the tissues of the aerial parts of these weeds (Iannone et al., Reference Iannone, White, Giussani, Cabral and Novas2011). Lolium rigidum and L. multiflorum Lam. (Italian ryegrass) are two important and commonly found weeds in areas cultivated with cereal crops that infected by the endophytic fungus, Neotyphodium occultans (Yamashita et al., Reference Yamashita, Iwamoto, Maruyama, Ichihara and Sawada2010; Kirkby et al., Reference Kirkby, Pratley, Hume, Faville, An and Wu2011), while L. perenne L. (perennial ryegrass) is also infected by the fungal endophyte species, N. lolii (Kane, Reference Kane2011). Recently, Leuchtmann et al. (Reference Leuchtmann, Bacon, Schardl, White and Tadych2014) recommended the reassignment of Neotyphodium species to the Epichloë genus with the exception of two species, Acremonium chilense and N. starrii.
The symbiosis of grasses with fungal endophytes helps the former alleviate the effects of drought stress (Kane, Reference Kane2011), while also improving their establishment, competitiveness and invasiveness (Yamashita et al., Reference Yamashita, Iwamoto, Maruyama, Ichihara and Sawada2010; Casas et al., Reference Casas, Gundel, Semmartin, Schnyder and Omacini2016). The infection of cool grasses with endophytes induces a range of adaptation mechanisms to drought stress, such as control of transpiration, improvement of water uptake due to a greater and deeper root system, and osmotic adjustment via the synthesis of various solutes (Malinowski and Belesky, Reference Malinowski and Belesky2000). Loline alkaloids (i.e. N-formylloline N-acetyllolin) have been identified in various endophyte-infected grasses (Schardl et al., Reference Schardl, Grossman, Nagabhyru, Faulkner and Mallik2007; Adhikari et al., Reference Adhikari, Boelt and Fomsgaard2016) and are known to play a significant role in drought tolerance of plants (Malinowski and Belesky, Reference Malinowski and Belesky2000; Schardl et al., Reference Schardl, Leuchtmann and Spiering2004).
Seed dormancy and soil moisture
The persistence of weed seeds in soil is a key factor that affects the density and competitiveness of weeds within a field (Efthimiadou et al., Reference Efthimiadou, Karkanis, Bilalis and Efthimiadis2009). The environmental conditions during seed development and maturation of maternal plants have been reported as factors that affect seed dormancy (Swain et al., Reference Swain, Hughues, Cook and Moss2006; Hoyle et al., Reference Hoyle, Steadman, Daws and Adkins2008). According to Swain et al. (Reference Swain, Hughues, Cook and Moss2006), the seeds of Alopecurus myosuroides Huds. (blackgrass) plants grown under warm and dry conditions were less dormant than those of plants grown under cool and wet conditions. Steadman et al. (Reference Steadman, Ellery, Chapman, Moore and Turner2004) also reported that seeds of L. rigidum Gaud. (annual ryegrass) developed at warm temperatures were also less dormant than those produced at low temperatures. Similar results were obtained by Figueroa et al. (Reference Figueroa, Herms, Cardina and Doohan2010), who found that the seeds of Senecio vulgaris L. (common groundsel) plants grown under cold conditions were more dormant as compared with those grown under warm conditions and which showed no dormancy.
Soil moisture in the maternal environment during seed development also affects the dormancy level of seeds. Wright et al. (Reference Wright, Seavers, Peters and Marshall1999) observed that seeds from Sinapis arvensis L. (wild mustard) plants subjected to water at 70% field capacity (FC) were more dormant than those from the plants subjected to water at 10% FC. Similar to these results, Luzuriaga et al. (Reference Luzuriaga, Escudero and Pérez-García2006) found that the provision of water in the maternal environment significantly reduced seed germination rate in S. arvensis, probably due to a higher dormancy level of the seeds.
In summary, the environmental conditions, particularly soil moisture and air temperature, during seed development in maternal plants of several weed species significantly affect the seed dormancy level; warm dry conditions have a positive effect on the germination rate of weed seeds, mostly due to lack of dormancy.
Allelopathy and drought
The allelopathic ability of weeds enhances their competitiveness against crops, while allelochemical production is influenced by both genetic and environmental factors (Qasem and Foy, Reference Qasem and Foy2001). Usually, water stress enhances the production of allelochemicals in several weed species, such as A. theophrasti Medic. (velvetleaf), Datura stramonium L. (jimsonweed) and Xanthium italicum Mor. (cocklebur; Borbély and Dávid, Reference Borbély and Dávid2008; Dávid and Borbély, Reference Dávid and Borbély2009).
Cyperus rotundus L. (purple nutsedge) is one of the most noxious weed species worldwide, which exhibits considerable allelopathic activity against vegetable crops due to its ability to release allelochemicals (i.e. alkaloids and phenolic acids) from its root system, while tolerating adverse environmental conditions (Dhima et al., Reference Dhima, Vasilakoglou, Stefanou, Gatsis, Paschalidis, Aggelopoulos and Eleftherohorinos2016; Peerzada, Reference Peerzada2017). Other important weed species that present significant allelopathic activity against various crops are A. retroflexus, Chenopodium album L. (fat-hen), Cirsium arvense (L.) Scop. (creeping thistle), C. dactylon, Papaver rhoeas L. (corn poppy), Solanum nigrum L. (black nightshade) and Xanthium strumarium L. (cocklebur) (Qasem and Hill, Reference Qasem and Hill1989; Tanveer et al., Reference Tanveer, Tahir, Nadeem, Younis, Aziz and Yaseen2008; Rezaie and Yarnia, Reference Rezaie and Yarnia2009; Ravlić, Reference Ravlić2016).
Vegetables’ production in an environment with changing climate
Any changes in environmental conditions are expected to significantly affect crop physiology and growth, yield, product quality and pesticide behaviour. However, the overall effects of climate change on agriculture depend on the extent of temperature increase, total rainfall and distribution in a region, the severity and frequency of storms and droughts, and the changes in plant × pathogens, plant × pests and plant × weeds interactions (IPCC, 2007; Siikamäki Reference Siikamäki2008). A brief overview of the effects of climate change on open-field vegetable production is presented in this section.
Overall, changes in temperatures and precipitation, as well as the limited availability of irrigation water and increased hailstorms and thunderstorms, are likely to affect vegetable yield significantly (Prasad and Chakravorty, Reference Prasad and Chakravorty2015). In the current century, an increase of 1.6 ± 0.27 °C in mean temperature is predicted in the Mediterranean region (Saadi et al., Reference Saadi, Todorovic, Tanasijevic, Pereira, Pizzigalli and Lionello2015). Moreover, the distribution of precipitation will vary significantly throughout the year, as well as between several countries of Southern Europe; however, in most of these countries, a decline in precipitation is also expected (Olesen et al., Reference Olesen, Trnka, Kersebaum, Skjelvåg, Seguin, Peltonen-Sainio, Rossi, Kozyra and Micale2011; Saadi et al., Reference Saadi, Todorovic, Tanasijevic, Pereira, Pizzigalli and Lionello2015). These changes in environmental conditions may affect vegetable production significantly, particularly production of spring crops, because of high temperatures and drought stress. For example, Ventrella et al. (Reference Ventrella, Charfeddine, Moriondo, Rinaldi and Bindi2012) reported that in the future climatic scenarios, an increase of air temperatures in the range of 2–5 °C may occur, which will decrease tomato yields in Italy. In addition, in areas that are suitable for open-field cultivation of tomato, yield is predicted to decline because of increased temperatures and drought stress (Silva et al., Reference Silva, Kumar, Shabani and Picanço2017). Moreover, high temperatures have been found to increase the length of the life-cycle of broccoli due to decreased developmental and growth rates, which result in delayed flowering (Lindemann-Zutz et al., Reference Lindemann-Zutz, Fricke and Stützel2016). Root development may also be affected by elevated soil temperatures due to alterations in several root functions (nutrient uptake, respiration, etc.) and architectural parameters (length and lateral root number, root branching) (Gray and Brady, Reference Gray and Brady2016).
Rising temperatures also affect vegetable quality negatively by affecting photosynthetic processes, such as the time taken for photoassimilation and the biosynthesis of sugars, organic acids and phenolic compounds (Mattos et al., Reference Mattos, Moretti, Jan, Sargent, Lima, Fontenelle, Ahmad and Rasool2014; Bisbis et al., Reference Bisbis, Gruda and Blanke2018). Furthermore, according to Potts et al. (Reference Potts, Biesmeijer, Kremen, Neumann, Schweiger and Kunin2010) and Nielsen et al. (Reference Nielsen, Reitan, Rinvoll and Brysting2017), a declining trend has already been observed in domesticated and wild pollinator species in Europe and other parts of the world; climatic changes have been reported as potential drivers of the reduction in pollinator species. Such changes can, therefore, indirectly affect vegetable production by affecting their respective pollinators (Prasad and Chakravorty, Reference Prasad and Chakravorty2015).
It should also be emphasized that decreasing soil moisture levels due to climatic change may lead to an increased persistence of herbicides, and thus, may lead to herbicide carryover to following crops, causing severe problems. It is well known that residues of soil-applied herbicides (i.e. sulphonylureas, triketones and imidazolinones) can cause severe damage to various vegetables, whereas according to Rahman et al. (Reference Rahman, James, Trolove and Dowsett2011), herbicide persistence is influenced by soil type and other environmental conditions (moisture, temperature). Ball et al. (Reference Ball, Yenish and Alby2003) observed that reduced soil moisture leads to a decline in imazamox degradation, while Wang et al. (Reference Wang, Liu, Wu, Huang, Xu and Tang2007) reported that an increase in soil moisture enhances microbial activity, as well as metsulphuron-methyl degradation. In other research, increased soil temperatures are reported to enhance herbicide dissipation via hydrolysis (Grey and McCullough, Reference Grey and McCullough2012). In this regard, Bailey (Reference Bailey2004) asserted that the duration of isoproturon efficacy against weeds declined by 25% in the UK over the period from 1980 to 2001 due to an increase in soil temperature. Consequently, the degradation of herbicides in soil is a complex process; the prediction of herbicide persistence in soil under the expected climate change scenarios of the future will provide useful information to help avoid crop damage by herbicide residues.
Effects of climate change on weed distribution and vegetable–weed competition in Southern Europe
Climate change is already occurring in Europe. Based on climate data from 1900 to 2005, rainfall has been increasing significantly in Northern Europe, while a decline has been witnessed in the Mediterranean region. By the end of the current century, a global temperature increase between 1.1 and 6.4 °C is predicted to occur under different climate scenarios (IPCC, 2007). Weiß et al. (Reference Weiß, Flörke, Menzel and Alcamo2007) reported that in the imminent future, incidences of drought are expected to increase by ten times in Southern Europe. If these projections are realized, it is anticipated that significant changes in weed flora composition and distribution will take place throughout Europe (Fig. 3) and other parts of the globe (McDonald et al., Reference McDonald, Riha, DiTommaso and DeGaetano2009; Castellanos-Frías et al., Reference Castellanos-Frías, Garcia De León, Bastida and Gonzalez-Andujar2016). Predicting weed invasion in a specific region undergoing climate change is a major challenge (Clements and DiTommaso, Reference Clements and DiTommaso2012). The spread of some weed species over new areas has already been recorded. Recently, Clements and DiTommaso (Reference Clements and DiTommaso2012) reported that several weed species have already expanded from the United States into Canada, while in Europe, climatic change during recent decades have already altered the weed flora in arable ecosystems (Peters et al., Reference Peters, Breitsameter and Gerowitt2014).
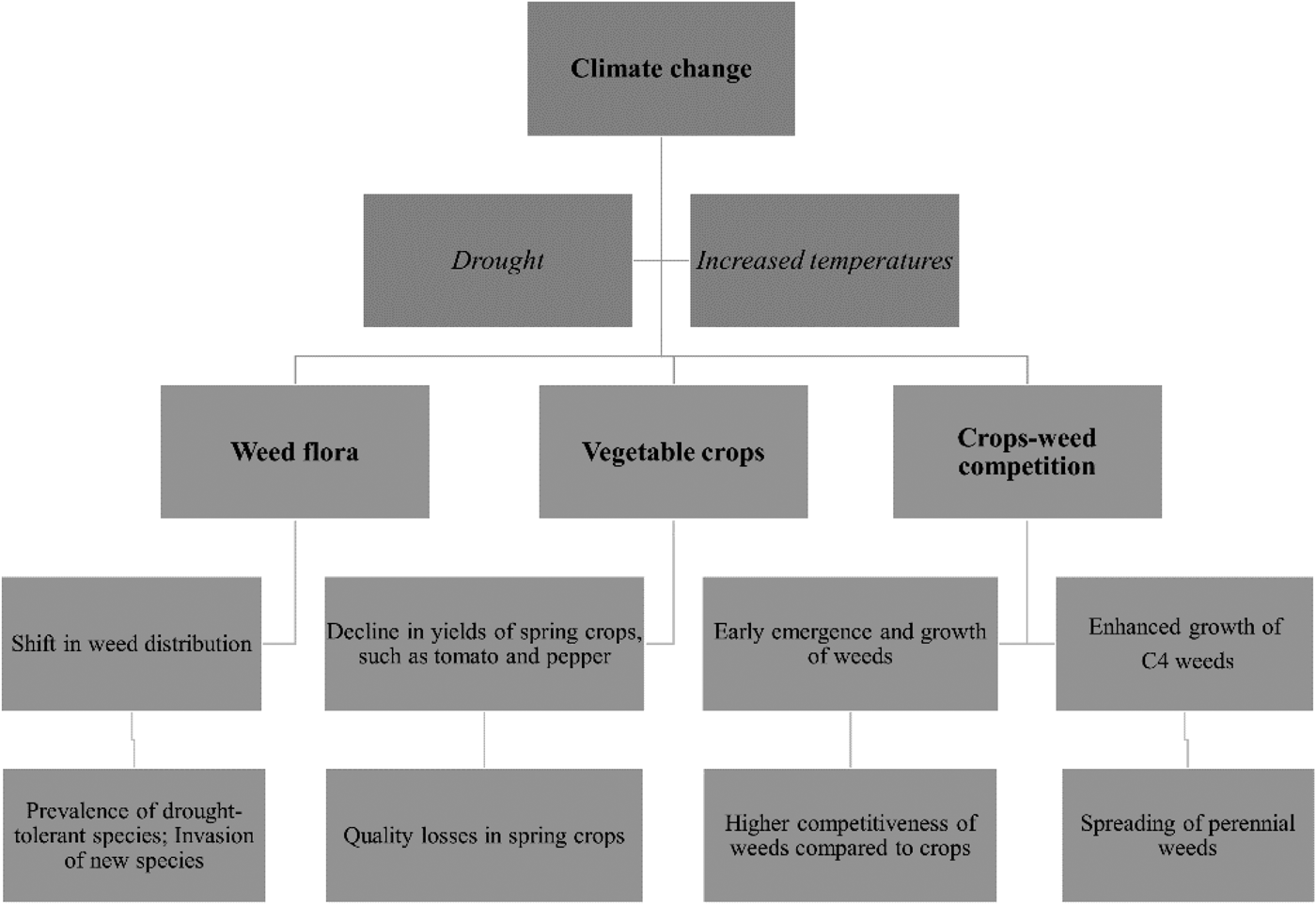
Fig. 3. Main effects of climate change on weed flora, vegetable production, and crop–weed competition in Southern Europe.
Irrespective of weed species, the projections about their geographical distribution under future climate conditions provide useful information for their early detection and management. For instance, Castellanos-Frías et al. (Reference Castellanos-Frías, Garcia De León, Bastida and Gonzalez-Andujar2016) reported that the geographic distribution of L. rigidum in Europe is expected to change, expanding to regions that are currently too cold for its survival but may not be in the near future. According to Castellanos-Frías et al. (Reference Castellanos-Frías, Garcia De León, Bastida and Gonzalez-Andujar2016), suitable regions in Europe for this weed species are predicted to increase by 108.7 or 167.3% depending on different climate change scenarios. Climate change can also influence the geographical distribution of crops and their losses caused by A. myosuroides. The effect of this weed on crop production has been projected to decline in regions where climate scenarios expected in the future predict more frequent periods of drought, whereas the diffusion of this weed is expected to increase from areas with higher temperatures to areas with lower temperatures (Stratonovitch et al., Reference Stratonovitch, Storkey and Semenov2012). Castellanos-Frías et al. (Reference Castellanos-Frías, García De Leõn, Pujadas-Salva, Dorado and Gonzalez-Andujar2014) reported that future environmental conditions may favour the spread of A. sterilis to Central and Northern Europe, while certain areas of the Mediterranean region may become unsuitable for its survival. Therefore, in Southern Europe, the winter weed species that are favoured by high soil moisture may spread to more suitable areas in the north, while its effect on winter pulses, cereals and vegetables that are commonly cultivated in the region is expected to decline.
Furthermore, climate change may favour the spread of drought-resistant perennial or spring weed species mentioned in previous sections of the current review; therefore, the negative effect of these weeds on crop production is projected to increase in southern Europe. Indeed, the present distribution of S. elaeagnifolium in several countries of southern Europe reveals the risk of further spread of the species to be noteworthy (Mekki, Reference Mekki2007). Moreover, rising temperatures are expected to enhance the early growth of D. stramonium as, according to Jursík et al. (Reference Jursík, Holec and Tyser2004), its minimum germination temperature is approximately 20 °C, while its optimum germination occurs at high temperatures of about 30 °C. These temperature requirements of D. stramonium seed germination prevent its earlier emergence at the ongoing temperatures in the regions (Cavero et al., Reference Cavero, Zaragoza, Suso and Pardo1999).
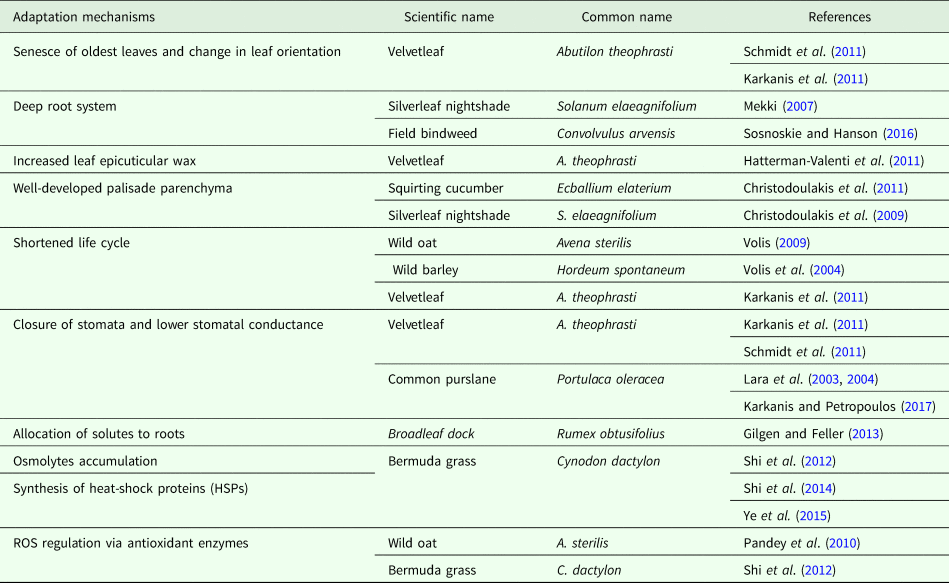
The projection of yield losses for vegetable crops due to weed competition under future climate conditions is also a challenge. It is important to point out once again that vegetable crops are more vulnerable to drought and increased temperatures due to their high water requirements and high water content. Moreover, crop–weed competition is a complex process that is influenced by several factors, including soil properties, environmental conditions, cultural practices and crop–weed competitiveness (Bilalis et al., Reference Bilalis, Karkanis and Efthimiadou2009; Efthimiadou et al., Reference Efthimiadou, Karkanis, Bilalis and Efthimiadis2009; Valerio et al., Reference Valerio, Lovelli, Perniola, Di Tommaso and Ziska2013). Drought-tolerant weed species may have a major effect on vegetable crops, making early management of these species necessary in the future (Table 1). Rodenburg et al. (Reference Rodenburg, Meinke and Johnson2011) reported that under conditions of drought and high temperatures, weed species with the C4 carbon fixation pathway will adapt better, thereby having a competitive advantage over C3 crop plants that use the C3 carbon fixation pathway in the photosynthesis process. Similarly, Hyvönen (Reference Hyvönen2011) speculated that rising temperatures will enhance the growth of C4 weeds, such as A. retroflexus, while Zand et al. (Reference Zand, Soufizadeh and Eskandari2006) observed that drought stress had a more negative effect on the competitiveness of C. album (C3 weed) compared with A. retroflexus (C4 weed). Regarding the effect of these conditions on crop–weed competition, Valerio et al. (Reference Valerio, Lovelli, Perniola, Di Tommaso and Ziska2013) concluded that yield losses during processing of tomatoes from weeds were greater under drought conditions. These results are related to the greater ability of weeds to adapt to drought stress than the vegetable crops (Korres et al., Reference Korres, Norsworthy, Tehranchian, Gitsopoulos, Loka, Oosterhius, Gealy, Moss, Burgos, Miller and Palhano2016). Similarly, Lovelli et al. (Reference Lovelli, Di Tommaso, Amato, Valerio and Perniola2010b) reported A. retroflexus to be more drought-resistant than pepper by exhibiting significant competition with this crop for water. Therefore, under drought conditions, weed control in pepper crop is more critical in order to achieve high yields (Lovelli et al., Reference Lovelli, Valerio, Di Tommaso and Perniola2013). Several other vegetables similarly exhibit lower competitiveness against weeds than field crops, such as maize and sunflower that have more vigorous early growth, higher leaf area and a deeper root system. It is important to highlight that the available herbicides for use on vegetable crops are limited in comparison with those for field crops, where pendimethalin is the major herbicide registered for control of grass and broad-leaved weeds in several vegetable crops, such as tomato, pepper, eggplant, cabbage, cauliflower, broccoli, leek, onion and parsley. Thus, timely weed control is pivotal for obtaining high yields, as well as for reducing the overall cost of weed management in vegetable crops. Moreover, the need for development of new herbicides that are suitable for vegetable crops is extremely important to achieve adequate weed control and to eradicate troublesome and noxious weeds, such as D. stramonium, P. oleracea, S. nigrum, X. strumarium, C. dactylon, C. rotundus, S. halepense and S. elaeagnifolium, which are projected to be more competitive under future climate conditions. Furthermore, according to Duke (Reference Duke2012), for almost two decades, no herbicide with a new mode of action has been introduced because of the increased cost of herbicide discovery and development. For the above-mentioned reasons, there is a growing need for new herbicides, especially for use in vegetable crops that are expected to be more vulnerable to weeds in the near future.
Table 1. Adaptation mechanisms to drought and high temperatures in different weed species commonly found in Southern Europe
In southern Europe, climate change is predicted to result in warmer winters. Rising temperatures may have a positive effect on weed growth, giving them an advantage in comparison with crops (Tungate et al., Reference Tungate, Israel, Watson and Rufty2007). In this regard, Wolfe et al. (Reference Wolfe, DeGaetano, Peck, Carey, Ziska, Lea-Cox, Kemanian, Hoffmann and Hollinger2018) reported that warmer winters can lead to increased weed pressure. Indeed, a long-term study conducted in Denmark during the period of 1987–1989 to 2001–2004 revealed that changes in environmental conditions (e.g. increased temperatures and rainfall levels) favour crop production, while the frequency of occurrence of several weed species increases (Andreasen and Stryhn, Reference Andreasen and Stryhn2012). Moreover, the base temperatures for shoot growth in A. myosuroides and G. aparine plants are 0.8 and −1.4 °C, respectively (Storkey and Cussans, Reference Storkey and Cussans2000). Considering that the critical temperatures for growth of several winter species are significantly low, it can be concluded that warmer winter temperatures will enhance their competitive ability against winter vegetable crops, particularly during the early growth stages. This is emphasized in the work of Mealor et al. (Reference Mealor, Cox and Booth2012), who found that the rise in spring temperatures enhanced early growth of Bromus tectorum L. (downy brome), while the frequency of its occurrence increased by 36% over a 5-year-long experiment. Ultimately, knowledge of the growth patterns of weeds during their early growth stages will prove to be a valuable decision support tool for weed management (Royo-Esnal et al., Reference Royo-Esnal, Torra, Conesa and Recasens2012).
Crop adaptation to climate change – future prospects
Under the projected future climate conditions, a multi-step approach must be implemented in order to minimize the negative effects of weeds on crop production. Firstly, it is important to develop maps predicting the geographical distribution of weeds under the expected climate change scenarios for specific areas (Chauhan et al., Reference Chauhan, Matloob, Mahajan, Aslam, Florentine and Jha2017). Secondly, extended trials should be conducted in order to evaluate the effect of extreme or increased temperatures and water stress on crop–weed competition, while combined effects of elevated CO2, increased temperatures and drought on weed and crop growth should also be examined. Moreover, studying the phenomenon of climate change can help understand the complex effects of climate change on weed-crop interference; in this context, it is also important to develop weed management strategies that are adapted to the expected future environmental conditions (Hayman and Sadras, Reference Hayman, Sadras, Preston, Watts and Crossman2006).
The adaptation of cultural practices (i.e. planting and sowing dates) to changing environmental conditions can help minimize the effect of climate change on crop production (Debaeke et al., Reference Debaeke, Casadebaig, Flenet and Langlade2017; Wiréhn, Reference Wiréhn2018). Under future climate scenarios, the discovery and development of new herbicides could make a significant contribution to the reduction of negative effects of weeds on crop production. It has furthermore been suggested that breeding of cultivars with improved stress tolerance (Chen et al., Reference Chen, McCarl and Chang2012) and enhanced competitiveness could help mitigate the negative effects of climate change on vegetable production (Korres et al., Reference Korres, Norsworthy, Tehranchian, Gitsopoulos, Loka, Oosterhius, Gealy, Moss, Burgos, Miller and Palhano2016). In this context, screening for drought tolerance and yield stability under stress conditions is pivotal for crop production, as well as a key step for the development of drought-tolerant cultivars (Cicevan et al., Reference Cicevan, Al Hassan, Sestras, Prohens, Vicente, Sestras and Boscaiu2016; Ganança et al., Reference Ganança, Freitas, Nóbrega, Rodrigues, Antunes, Gouveia, Rodrigues, Chaϊr, Pinheiro De Carvalho and Lebot2018). Finally, raising awareness among farmers regarding the effect of climate change on crop production is extremely important (Thi Lan Huong et al., Reference Thi Lan Huong, Shun Bo and Fahad2017), as some farmers consider climate change to be significant for their farming practices, while others do not believe in its occurrence (Takahashi et al., Reference Takahashi, Burnham, Terracina-Hartman, Sopchak and Selfa2016).
Conclusions
Several commonly found weeds in Southern Europe exhibit great potential to adapt to drought conditions and increased temperatures. Under future climate scenarios, the prediction of weed invasion in a specific region and the spread of weed species over new areas is a major challenge. In southern Europe, weed species that are favoured by high soil moisture may be spread over new areas to the north and their negative effects on vegetable production in these areas are thus expected to increase. Moreover, C4 weed species will be better adapted to future climate conditions; therefore, they will be more competitive in comparison with C3 weeds and crops. In addition, increasing temperatures can enhance the early growth of several weed species, giving them an advantage over vegetable crops. Accordingly, adaptation of cultural systems to these environmental conditions will be crucial in terms of minimizing the negative effects of climate change on crop production.
Author ORCIDs
A. Karkanis, 0000-0003-0477-9923
Financial support
This research received no specific grant from any funding agency, commercial or not-for-profit sectors.
Conflict of interest
None.
Ethical standards
Not applicable.




