



Introduction
Addressing the challenges climate change poses on feed and food production emerges as a global priority. Although crucial for feed and food production, livestock and crops are frequently cited as significant contributors to deforestation and soil degradation, impacting ecosystem services in tropical and subtropical environments (Carriazo et al., Reference Carriazo, Labarta and Escobedo2020). Besides, the variability in global climatic conditions impacts land use and Earth's ecosystems at various scales (Cardona et al., Reference Cardona, Ramírez, Morales, Murgueitio, Orozco, Vera, Sánchez, Flores, Sánchez and Rosales2014).
The increasing demand for natural resources, global concerns about climate change and evolving environmental laws have been forcing the development of technologies to increase land-use efficiency and sustainability of production systems in agriculture. Adopting integrated systems, such as silvopastoral systems (SPSs), is an alternative to contributing to the sustainability of agricultural systems (Jose and Dollinger, Reference Jose and Dollinger2019). SPSs constitute a modality of agroforestry systems where trees or shrubs, pastures and animals are managed in the same area (Alonso, Reference Alonso2011; Costa et al., Reference Costa, De Mello, Dubeux, Dos Santos, Lira, Oliveira and Apolinário2016; Dos Santos et al., Reference Dos Santos, Junior, Dos Santos, De Andrade Lira, De Oliveira Apolinário, Costa, De Lima Coêlho, Peixôto and Da Silva Santos2020; Herrera et al., Reference Herrera, De Mello, Apolinário, Dubeux Júnior, Da Silva, Dos Santos and Da Cunha2020; De Macêdo Carvalho et al., Reference De Macêdo Carvalho, De Mello, Da Cunha, De Oliveira Apolinário, Da Silva, De Miranda Costa, De Carvalho and Dos Santos2022a, Reference De Macêdo Carvalho, De Mello, Da Cunha, De Oliveira Apolinário, Júnior, Pessoa, De Oliveira Neto and Da Silva2022b).
The interaction among different components of SPSs provides different ecosystem services, including increased soil fertility and conservation (Lima et al., Reference Lima, Dubeux, Santos, Mello, Lira and Cunha2018; Herrera et al., Reference Herrera, De Mello, Apolinário, Dubeux Júnior, Da Silva, Dos Santos and Da Cunha2020; Paciullo et al., Reference Paciullo, Fernandes, Carvalho, Morenz, Lima, Maurício and Gomide2021; Smith et al., Reference Smith, Bentrup, Kellerman, MacFarland, Straight, Ameyaw and Stein2022). Additionally, the arboreous component in SPSs typically contributes to increased litter deposition. When legume trees or shrubs are incorporated into SPSs, they may offer numerous advantages, including biological N2 fixation (BNF) and enhanced nutrient cycling via deposition and decomposition of litter and animal excreta richer in nitrogen (N) compared to litter or cattle excreta from grass-based systems (Apolinário et al., Reference Apolinário, Dubeux, Lira, Ferreira, Mello, Santos, Sampaio and Muir2015; Dubeux et al., Reference Dubeux, Muir, Apolinário, Nair, Lira and Sollenberger2017; Da Silva et al., Reference Da Silva, Dubeux, De Melo, Da Cunha, Dos Santos, Apolinário and De Freitas2021). Furthermore, trees may contribute to improving thermal comfort for animals, improving animal performance and consequently, generating animal products to supply human food (Giustina et al., Reference Giustina, Carnevalli, Romano, Antonio and Eckstein2017), increasing forage production and quality (Costa et al., Reference Costa, De Mello, Dubeux, Dos Santos, Lira, Oliveira and Apolinário2016) and providing income diversification (Apolinário et al., Reference Apolinário, Dubeux, Lira, Ferreira, Mello, Santos, Sampaio and Muir2015). SPSs may also provide several environmental benefits such as the conservation of biodiversity (Dos Santos et al., Reference Dos Santos, Junior, Dos Santos, De Andrade Lira, De Oliveira Apolinário, Costa, De Lima Coêlho, Peixôto and Da Silva Santos2020), atmospheric carbon sequestration (Hoosbeek et al., Reference Hoosbeek, Remme and Rusch2018), mitigation of greenhouse gas (GHG) emissions (Frota et al., Reference Frota, Carneiro, Pereira, Berndt, Frighetto, Sakamoto, Moreira Filho, Cutrim Júnior and Carvalho2017) and improvements in the water quality (Moreno et al., Reference Moreno, Franca, Correia and Godinho2014).
Successful experiences with SPSs have been documented in Australia, Africa, Asia, Europe, Latin America, Mexico and the United States, where positive impacts on system productivity and ecosystem service delivery have been reported compared to conventional systems (Murgueitio et al., Reference Murgueitio, Calle, Uribe, Calle and Solorio2011; Calle et al., Reference Calle, Murgueitio, Chará, Molina, Zuluaga and Calle2013; Cardona et al., Reference Cardona, Ramírez, Morales, Murgueitio, Orozco, Vera, Sánchez, Flores, Sánchez and Rosales2014; Torralba et al., Reference Torralba, Fagerholm, Burgess, Moreno and Plieninger2016; Lima, et al., Reference Lima, Paciullo, Morenz, Gomide, Rodrigues and Chizzotti2019a, Reference Lima, Paciullo, Silva, Morenz, Gomide, Rodrigues, Bretas and Chizzotti2019b; Castillo et al., Reference Castillo, Tiezzi and Franzluebbers2020; Yang et al., Reference Yang, Zhu, Duan, Lu, Zhang, Wan, Xu, Zhang and Li2020; Schinato et al., Reference Schinato, Munka, Olmos and Bussoni2023; Seidou et al., Reference Seidou, Offoumon, Sanni Worogo, Houaga, Koara Yarou, Azalou, Adambi Boukari, Idrissou, Houinato and Alkoiret Traoré2023). In recent years, satisfactory results in terms of animal performance, soil health or economic viability were reported in Brazil (Apolinário et al., Reference Apolinário, Dubeux, Lira, Ferreira, Mello, Santos, Sampaio and Muir2015; Costa et al., Reference Costa, De Mello, Dubeux, Dos Santos, Lira, Oliveira and Apolinário2016; Lima et al., Reference Lima, Dubeux, Santos, Mello, Lira and Cunha2018; Da Silva et al., Reference Da Silva, Dubeux, De Melo, Da Cunha, Dos Santos, Apolinário and De Freitas2021; Herrera et al., Reference Herrera, De Mello, De Oliveira Apolinário, Dubeux Júnior, Da Cunha and Dos Santos2021; Zambrano et al., Reference Zambrano, Edvan, Oliveira, Da Costa Araujo, Costa, Da Silva, Barros, De Sousa Reis and Dias-Silva2021; De Macêdo Carvalho et al., Reference De Macêdo Carvalho, De Mello, Da Cunha, De Oliveira Apolinário, Da Silva, De Miranda Costa, De Carvalho and Dos Santos2022a, Reference De Macêdo Carvalho, De Mello, Da Cunha, De Oliveira Apolinário, Júnior, Pessoa, De Oliveira Neto and Da Silva2022b), and other countries around the world, such as the United States, Portugal, Nicaragua, Cuba, India and China (Haile et al., Reference Haile, Nair and Nair2010; Guerra and Pinto-Correia, Reference Guerra and Pinto-Correia2016; Hoosbeek et al., Reference Hoosbeek, Remme and Rusch2018; Alonso-Amaro et al., Reference Alonso-Amaro, Lezcano-Fleires and Suris-Campos2019; Yadav et al., Reference Yadav, Gendley, Sahu, Patel, Chandraker and Dubey2019; Yang et al., Reference Yang, Zhu, Duan, Lu, Zhang, Wan, Xu, Zhang and Li2020; Greene et al., Reference Greene, Kazanski, Kaufman, Steinberg, Johnson, Cook-Patton and Fargione2023).
On the other hand, the integration of tree and forage species may also negatively affect the herbage mass and forage accumulation by reducing photosynthetically active radiation reaching the pasture canopy. This occurs under conditions of intense shading or inappropriate selection of species, spacing during establishment or management practices related to pruning and trimming (Lopes et al., Reference Lopes, Paciullo, Araújo, Gomide, Morenz and Villela2017; Lima et al., Reference Lima, Paciullo, Morenz, Gomide, Rodrigues and Chizzotti2019a, Reference Lima, Paciullo, Silva, Morenz, Gomide, Rodrigues, Bretas and Chizzotti2019b). Besides the light competition, tree species can compete for water and nutrients in SPSs. Furthermore, the increasing N cycling with legume trees incorporation into SPSs can lead to greater nitrous oxide (N2O) emissions compared to monoculture pastures (Bretas et al., Reference Bretas, Paciullo, Alves, Martins, Cardoso, Lima, Rodrigues, Silva and Chizzotti2020; Cardoso et al., Reference Cardoso, Alari, de Andrade, Nascimento, van Cleef, Alves, Ruggieri and Malheiros2022), conflicting with one of the major regulating services of agricultural systems. Thus, the complex interaction between soil, forage, trees, animals and the environment may significantly impact SPS feasibility and practical application, underscoring the importance of considering these dynamics when adopting such systems. Several design options for SPS establishment and management have been used worldwide and are determinants of the benefits obtained from the systems. Despite recent advances, the success of integrated systems relies on well-designed planning and establishment practices (Giustina et al., Reference Giustina, Carnevalli, Romano, Antonio and Eckstein2017). Thus, knowledge about SPSs and multidisciplinary support is necessary to overcome possible barriers.
Although previous reviews (Alonso, Reference Alonso2011; Murgueitio et al., Reference Murgueitio, Calle, Uribe, Calle and Solorio2011; Moreno et al., Reference Moreno, Franca, Correia and Godinho2014; Dubeux et al., Reference Dubeux, Muir, Apolinário, Nair, Lira and Sollenberger2017) have significantly contributed to the understanding of environmental benefits from SPSs, none of them has comprehensively delved into provisioning, supporting, regulating and cultural ecosystem services worldwide. Additionally, as research in this area evolves, more recent studies have emerged, providing additional and updated perspectives on these services.
Based on this context, this review aims to fill such gaps by offering a comprehensive analysis of the ecosystem services provided by SPSs in different countries. Our approach seeks to highlight the main benefits of SPSs to the environment and society and discuss potential shortcomings and challenges through a literature review.
Methodology
The review focused on recent studies, mainly those published between 2010 and 2024, with global relevance to SPS and its ecosystem services. We used Google Scholar, Scopus and Web of Science databases for literature screening, using ‘silvopastoral systems’ and ‘agroforestry systems’ combined with the terms ‘ecosystem services’, ‘greenhouse gas’, ‘forage traits’, ‘animal performance’, ‘biofuels’, ‘nutrient cycling’, ‘climate regulation’, ‘biological nitrogen fixation’, ‘biodiversity’, ‘cultural services’, ‘water quality’ and ‘soil health’ as keywords. Both review and research papers published in English between 2010 and 2024 were included in the literature search. For some topics with few recent publications, the date range filter was not applied. The literature was then screened by relevance, adherence to the topic and rigour of evaluations. In total, 200 papers without any restrictions about country, soil type or climatic conditions were recovered and 158 were reviewed. When several relevant publications were found for the same reviewed topic, studies from different countries or ecoclimatic regions were prioritized.
Ecosystem services of SPSs
Ecosystem services can be defined as all benefits directly or indirectly provided by ecosystems to meet the demand for human survival, life and well-being (MEA, 2005; Zhao et al., Reference Zhao, Liu and Wu2020). Nahed-Toral et al. (Reference Nahed-Toral, Valdivieso-Pérez, Aguilar-Jiménez, Cámara-Cordova and Grande-Cano2013) indicated that SPSs provide additional benefits to society at the local/producer level, as well as at the regional/landscape and global levels, compared to conventional pastures dominated by grasses in monocultures. Based on the Millennium Ecosystem Assessment – MEA (2005), ecosystem services can be divided into provisioning, supporting, regulating, and cultural services.
Although some authors criticize the use of the term ‘ecosystem services’ in production systems like SPSs – arguing that it oversimplifies complex ecological interactions into mere outputs for human benefit, prioritizing economic gain over ecological health and overlooking social and cultural values associated with nature – this review takes an integrated approach. It considers the four classes of ecosystem services through a holistic view, emphasizing both the positive and negative aspects of the system. This approach acknowledges the importance of ecological integrity, social equity and the cultural significance of the entire system, fostering a more comprehensive understanding of the relationship between production systems and ecosystems. If well-designed and managed, SPSs can provide multiple ecosystem services (Fig. 1), thus contributing significantly to the sustainability of agricultural systems (Smith et al., Reference Smith, Bentrup, Kellerman, MacFarland, Straight, Ameyaw and Stein2022).

Figure 1. Examples of the different classes of ecosystem services provided by SPSs. The magnitude of all the listed potential ecosystem services depends on the interaction of multiple factors, including system design, choice of species, management and climate conditions.
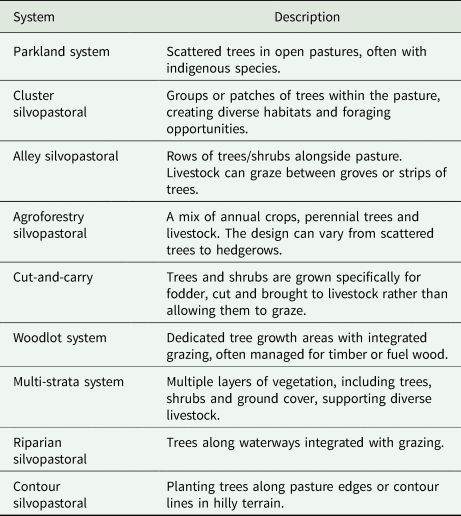
Despite the several benefits provided by SPSs, these benefits and their magnitude might be related to the species, design and management adopted. SPSs are broadly categorized into grazing or tree-fodder systems (Nair, Reference Nair2014a, Reference Nair2014b). Grazing systems are based on grazed pastures under widely spaced or scattered trees such as the extensive Parkland System of sub-Saharan Africa. Tree-fodder systems are based on stall feeding of animals with fodder from trees or shrubs grown in fodder banks (cut-and-carry systems). Each broad category can be divided into several types of SPSs according to their establishment, design and primary purpose. Table 1 presents an overview of the various silvopastoral types/designs and their key characteristics.
Table 1. Overview of the various silvopastoral types/designs worldwide adopted and their summarized descriptions
Provisioning services
Provisioning services include all products obtained from ecosystems, such as wood (Nahed-Toral et al., Reference Nahed-Toral, Valdivieso-Pérez, Aguilar-Jiménez, Cámara-Cordova and Grande-Cano2013; Apolinário et al., Reference Apolinário, Dubeux, Lira, Ferreira, Mello, Santos, Sampaio and Muir2015), human food (Giustina et al., Reference Giustina, Carnevalli, Romano, Antonio and Eckstein2017; Sánchez-Santana et al., Reference Sánchez-Santana, López-Vigoa, Iglesias-Gómez, Lamela-López and Soca-Perez2018), forage or roughage for animal feed (Dos Santos et al., Reference Dos Santos, Junior, Dos Santos, De Andrade Lira, De Oliveira Apolinário, Costa, De Lima Coêlho, Peixôto and Da Silva Santos2020; Yang et al., Reference Yang, Zhu, Duan, Lu, Zhang, Wan, Xu, Zhang and Li2020; Paciullo et al., Reference Paciullo, Fernandes, Carvalho, Morenz, Lima, Maurício and Gomide2021), biofuels (Moreno et al., Reference Moreno, Franca, Correia and Godinho2014; Rade et al., Reference Rade, Cañadas, Zambrano, Molina, Ormaza and Wehenkel2017), natural medicines (Root-Bernstein and Jaksic, Reference Root-Bernstein and Jaksic2013; Olivares-Pérez et al., Reference Olivares-Pérez, Rojas-Hernández, Avilés-Nova, Camacho-Díaz, Jiménez-Guillén and Quiroz-Cardozo2016) and ornamental resources (Dubeux et al., Reference Dubeux, Muir, Apolinário, Nair, Lira and Sollenberger2017).
Wood, fence posts and live fences
The arboreous component in SPSs can simultaneously provide fence posts, live fences, forage and shade to animals, subsequently providing wood. Many tree species can potentially be exploited as live fences, especially the arboreal legume tree ‘gliricidia’ [Gliricidia sepium (Jacq.) Kunth ex Walp], which farmers have used to delimit pastures in the Pernambuco Tropical Region, Brazil (De Macêdo Carvalho, et al., Reference De Macêdo Carvalho, De Mello, Da Cunha, De Oliveira Apolinário, Da Silva, De Miranda Costa, De Carvalho and Dos Santos2022a, Reference De Macêdo Carvalho, De Mello, Da Cunha, De Oliveira Apolinário, Júnior, Pessoa, De Oliveira Neto and Da Silva2022b). At Tabasco State, Mexico, live fences installed around the pastures for cattle production are the main form to cover rural landscapes together with wood-purpose species such as the Latin Rosea (Tabebuia rosea Bertolt. A. DC) and the ‘mafumeira’ (Ceiba pentandra L. Gaertn.) (Nahed-Toral et al., Reference Nahed-Toral, Valdivieso-Pérez, Aguilar-Jiménez, Cámara-Cordova and Grande-Cano2013). In Latin America, many tree species are considered valuable to produce wood aimed at industrial sectors of construction and woodwork, such as the African mahogany (Swietenia macrophylla King) and the tropical cedar (Cedrela odorata L.) (Murgueitio et al., Reference Murgueitio, Barahona, Chará, Flores, Mauricio and Molina2015). In addition, Apolinário et al. (Reference Apolinário, Dubeux, Lira, Ferreira, Mello, Santos, Sampaio and Muir2015) reported that ‘sabia’ trees (Mimosa caesalpiniifolia Benth.) aged 5 years can be sold as fuel or used as great-quality wood in the Brazilian tropical region. In Brazil, Eucalyptus spp. is considered an important option for wood production due to the high growth rate, easy cultivation and wood quality (De Oliveira et al., Reference De Oliveira, Menezes, Gonçalves, De Araújo, Ramirez, Guimarães Júnior, Jayme and Lana2022). Eucalyptus spp. has also been introduced to SPSs in Europe (Báder et al., Reference Báder, Németh, Vörös, Tóth and Novotni2023) and Australia (Francis et al., Reference Francis, Venn, Lewis and Brawner2022) for timber production and additional income due to their great growth rate. Francis et al. (Reference Francis, Venn, Lewis and Brawner2022) evaluated the financial performance of SPSs at four case study sites in southern Australia and reported financial attractivity when silvicultural treatments were implemented to increase timber production.
Forage production for livestock feed and human food supply
Humans can consume tree products such as fruits, but SPSs also supplies human food indirectly through the products from herbivores (Dubeux et al., Reference Dubeux, Muir, Apolinário, Nair, Lira and Sollenberger2017), especially meat and milk. Fruit trees grown on pastures are common in Romania's Western Carpathians (Săndoiu and Cojocariu, Reference Săndoiu and Cojocariu2014). Giustina et al. (Reference Giustina, Carnevalli, Romano, Antonio and Eckstein2017) described well-established fruit trees such as cajá (Spondias mombin L.), red guava (Psidium guajava L.), cashew (Anacardium occidentale L.), acerola (Malpighia glabra L.), dwarf green coconut (Cocos nucifera L.) and banana (Musa spp.) in SPSs at Mato Grosso State, Brazil. Fruit crops, which produce seeds and pods that feed humans, are also traditional in SPSs of the French ‘pre-vergers’, the Dutch ‘boguards’, the Spanish ‘pomaradas’ and the ‘streuobstwiesen’ of Central Europe (Moreno et al., Reference Moreno, Franca, Correia and Godinho2014).
Tree species with a large leaf proportion on their morphological composition or edible fruits may also be satisfactorily incorporated into ruminant feed as roughage or forage sources due to satisfactory energy and protein contents (Ku Vera et al., Reference Ku Vera, Briceño, Ruiz, Mayo, Ayala, Aguilar, Solorio and Ramírez2014). The incorporation of forage legume trees such as Leucaena leucocephala (Lam.) De Wit on the SPSs is an alternative to increase animal performance (Calle et al., Reference Calle, Murgueitio, Chará, Molina, Zuluaga and Calle2013; Carriazo et al., Reference Carriazo, Labarta and Escobedo2020), especially in the driest periods of the year when forage production is reduced due to environmental constraints. Kumar et al. (Reference Kumar, Gautam, Ghosh, Singh, Kumar, Kumar, Alam, Taria, Singh, Yadav and Arunachalam2024) highlighted the crucial role of SPSs in providing year-round fodder supply while restoring degraded landscapes. Murgueitio et al. (Reference Murgueitio, Barahona, Chará, Flores, Mauricio and Molina2015) indicated that the species Samanea saman (Jacq.) Merr., ‘gliricidia’ and arboreal legumes of the genus Prosopis spp. may contribute as forage sources in countries of Latin America. In Brazil, ‘gliricidia’ and ‘sabia’ are considered alternative forage legume trees with the potential to be exploited as forestry (Apolinário et al., Reference Apolinário, Dubeux, Lira, Ferreira, Mello, Santos, Sampaio and Muir2015; Costa et al., Reference Costa, De Mello, Dubeux, Dos Santos, Lira, Oliveira and Apolinário2016; Lima et al., Reference Lima, Dubeux, Dos Santos, De Mello, Lira and Cunha2016; Dos Santos et al., Reference Dos Santos, Junior, Dos Santos, De Andrade Lira, De Oliveira Apolinário, Costa, De Lima Coêlho, Peixôto and Da Silva Santos2020; Da Silva et al., Reference Da Silva, Dubeux, De Melo, Da Cunha, Dos Santos, Apolinário and De Freitas2021; Herrera et al., Reference Herrera, De Mello, De Oliveira Apolinário, Dubeux Júnior, Da Cunha and Dos Santos2021). In Tejupilco, Mexico, non-legume species such as Guazuma ulmifolia Lam. and Crescentia alata Kunth are considered tree species with considerable forage potential by farmers (Olivares-Pérez et al., Reference Olivares-Pérez, Rojas-Hernández, Avilés-Nova, Camacho-Díaz, Jiménez-Guillén and Quiroz-Cardozo2016). In Europe, the potential of ash (Fraxinus spp.), alder (Alnus spp.) and willow (Salix spp.) trees as fodder for livestock in SPSs has also been reported (Luske and Van Eekeren, Reference Luske and Van Eekeren2018). Temperate species such as Salix spp. are also common fodder trees grown in New Zealand, while L. leucocephala and Desmanthus spp. are common in Australia (Vandermeulen et al., Reference Vandermeulen, Ramírez-Restrepo, Beckers, Claessens and Bindelle2018). Tree species such as Quercus spp., Juglans spp. and Robinia pseudoacacia are considered multipurpose trees for SPSs in the United States, offering shade, timber and livestock fodder (Orefice et al., Reference Orefice, Carroll, Conroy and Ketner2017). L. leucocephala, Acacia angustissima, Leucaena diversifolia and Leucaena pallida are important fodder trees in Africa (Franzel et al., Reference Franzel, Carsan, Lukuyu, Sinja and Wambugu2014). It is worth mentioning that forage intake in SPSs does not need to be exclusively grazing, as it can include the cut-and-carry systems widely adopted in sub-Saharan Africa and Asia (Franzel et al., Reference Franzel, Carsan, Lukuyu, Sinja and Wambugu2014; Seruni et al., Reference Seruni, Aguilar, Cai, Gold and Roshetko2021) or harvesting forage for storage and later use as hay or silage during periods of scarcity (Londoño-Carmona et al., Reference Londoño-Carmona, Mahecha-Ledezma and Angulo2020; Yang et al., Reference Yang, Zhu, Duan, Lu, Zhang, Wan, Xu, Zhang and Li2020; Sarabia-Salgado et al., Reference Sarabia-Salgado, Alves, Boddey, Urquiaga, Galindo, Flores-Coello, Santos, Jimenez-Ocampo, Ku-Vera and Sánchez2023).
On the other hand, forage mass and accumulation in open pastures are often greater than in SPSs (Lima et al., Reference Lima, Paciullo, Morenz, Gomide, Rodrigues and Chizzotti2019a, Reference Lima, Paciullo, Silva, Morenz, Gomide, Rodrigues, Bretas and Chizzotti2019b; Dibala et al., Reference Dibala, Jose, Gold, Hall, Kallenbach and Knapp2021; Paciullo et al., Reference Paciullo, Fernandes, Carvalho, Morenz, Lima, Maurício and Gomide2021; Schinato et al., Reference Schinato, Munka, Olmos and Bussoni2023). Reduced solar radiation is generally considered the main limiting factor of forage accumulation in SPSs (Paciullo et al., Reference Paciullo, Pires, Aroeira, Morenz, Maurício, Gomide and Silveira2014; Giustina et al., Reference Giustina, Carnevalli, Romano, Antonio and Eckstein2017; Lima et al., Reference Lima, Dubeux, Santos, Mello, Lira, Cunha, De Freitas and Apolinário2020). Torralba et al. (Reference Torralba, Fagerholm, Burgess, Moreno and Plieninger2016) also observed the negative effect of SPSs on biomass production in a meta-analysis of 53 studies conducted in Europe. However, the interaction between the components within the system will determine the productive capacity and the potential of ecosystem service delivery. Cruz et al. (Reference Cruz, Santos, Silva, Ferreira, Magalhães, Martuscello and Fonseca2020) evaluated the dry matter production of forage peanut (Arachis pintoi) under increasing shading levels (0, 30, 45, 75%) and observed a quadratic effect, with maximum production at 30% of shade and forage peanut adaptation up to 45% of shade. Da Cruz et al. (Reference Da Cruz, Anésio, Santos and Dumont2024) observed no differences in total herbage mass production and tiller density of BRS Tamani (Megathyrsus maximus cv. Tamani) under increasing shading levels (0, 30, 45, 75%), suggesting BRS Tamani as a promising cultivar for SPS adoption. The authors justified it due to the high phenotypic plasticity of BRS Tamani, changing the photosynthetic apparatus and morphological characteristics to adapt to intense shading conditions. However, the shade effect of SPSs on herbage mass might occur in the number of grazing cycles, with shaded plants presenting longer regrowth periods and a lower number of grazing cycles over the year. This was observed by Da Cruz et al. (Reference Da Cruz, Anésio, Santos and Dumont2024) during the winter and autumn seasons. Differently, Da Silva et al. (Reference Da Silva, Dubeux, De Melo, Da Cunha, Dos Santos, Apolinário and De Freitas2021) and Zambrano et al. (Reference Zambrano, Edvan, Oliveira, Da Costa Araujo, Costa, Da Silva, Barros, De Sousa Reis and Dias-Silva2021) found greater herbage mass in SPSs than in monoculture. They justified it due to the capacity of SPSs for soil fertility restoration, greater organic matter supply, greater N-mineralization through litter deposition and maintenance of the soil moisture availability by the shading effect. The time for establishing trees and shrubs and its impact on sunlight availability for the companion forage grass must be considered. Strategies such as appropriate selection of tree species for partial shading, spacing between trees, controlled pruning and species diversification may contribute to reducing light competition and increase nutrient cycling within the system (Gomes et al., Reference Gomes, Pedreira, Santos, Bosi and Pedreira2020; Londoño-Carmona et al., Reference Londoño-Carmona, Mahecha-Ledezma and Angulo2020; Dibala et al., Reference Dibala, Jose, Gold, Hall, Kallenbach and Knapp2021; Schmiedgen et al., Reference Schmiedgen, Komainda, Kayser, Tonn and Isselstein2022). In addition, the choice of forage species plays a vital role in the viability of SPSs due to the variability in phenotypic plasticity among species under shaded conditions (Abraham et al., Reference Abraham, Kyriazopoulos, Parissi, Kostopoulou, Karatassiou, Anjalanidou and Katsouta2014; Lima et al., Reference Lima, Paciullo, Morenz, Gomide, Rodrigues and Chizzotti2019a, Reference Lima, Paciullo, Silva, Morenz, Gomide, Rodrigues, Bretas and Chizzotti2019b).
Regarding animal performance, the well-managed SPSs can potentially increase milk and meat production for the human food supply (Sánchez-Santana et al., Reference Sánchez-Santana, López-Vigoa, Iglesias-Gómez, Lamela-López and Soca-Perez2018). The greater beef cattle average daily gain per animal under SPSs established with signalgrass (Urochloa decumbens Stapf. cv. Basilisk) in a consortium with the tree legumes Acacia mangium Willd., A. angustissima (Mill.) Kuntze, Mimosa artemisiana Heringer & Paula and Eucalyptus grandis W. Hill ex Maiden compared to conventional systems (signalgrass monoculture) was reported by Paciullo et al. (Reference Paciullo, De Castro, Gomide, Maurício, Pires, Müller and Xavier2011) in Minas Gerais State, Brazil. In the same experimental area, Paciullo et al. (Reference Paciullo, Pires, Aroeira, Morenz, Maurício, Gomide and Silveira2014) reported a greater milk yield per animal in the first evaluation year (10.4 kg/cow/day) for the SPSs compared to the monoculture (9.5 kg/cow/day). Barros-Rodríguez et al. (Reference Barros-Rodríguez, Solorio-Sánchez, Ku-Vera, Ayala-Burgos, Sandoval-Castro and Solís-Pérez2012) also reported greater sheep weight gain in SPSs using ‘leucena’ [L. leucocephala (Lam.) De Wit] legume trees in Michoacán, Mexico. Similarly, Da Silva et al. (Reference Da Silva, Dubeux, De Melo, Da Cunha, Dos Santos, Apolinário and De Freitas2021) observed that SPSs using signalgrass and ‘gliricidia’ enhanced livestock weight gains compared with signalgrass in monoculture. Cardona et al. (Reference Cardona, Ramírez, Morales, Murgueitio, Orozco, Vera, Sánchez, Flores, Sánchez and Rosales2014) explain that these greater animal productive responses on SPSs are probably due to the lower seasonality of forage production and the increase in forage nutritional value along the year compared to conventional systems, which increases pasture carrying capacity and animal performance. Additionally, tree canopies may generate a favourable microclimate for forage production and provide natural shading for the animals (Săndoiu and Cojocariu, Reference Săndoiu and Cojocariu2014), contributing to the thermal comfort of grazing animals (Ku Vera et al., Reference Ku Vera, Briceño, Ruiz, Mayo, Ayala, Aguilar, Solorio and Ramírez2014; Paciullo et al., Reference Paciullo, Pires, Aroeira, Morenz, Maurício, Gomide and Silveira2014; Vieira et al., Reference Vieira, Pilatti, Czekoski, Fonsêca, Herbut, Angrecka, De Souza Vismara, De Paulo Macedo, Dos Santos and Paśmionka2021; Schinato et al., Reference Schinato, Munka, Olmos and Bussoni2023), and likely contributing to improving the long-term animal performance. Lemes et al. (Reference Lemes, Garcia, Pezzopane, Brandão, Watanabe, Cooke, Sponchiado, De Paz, Camplesi, Binelli and Gimenes2021) demonstrated that SPSs can improve animal welfare and performance compared to unshaded grazing systems. However, intensive shading can negatively affect animal performance by reducing herbage allowance, requiring suitable management of shading intensity and canopy light interception to ensure provisioning services.
Trees might help to reduce wind speed and extreme temperatures, contributing to water preservation and reducing the seasonality of forage production in the tropics (Cardona et al., Reference Cardona, Ramírez, Morales, Murgueitio, Orozco, Vera, Sánchez, Flores, Sánchez and Rosales2014). Besides, the absorption of nutrients by the plants due to greater cycling in SPSs compared to pasture monoculture can improve forage nutritive value (Herrera et al., Reference Herrera, De Mello, De Oliveira Apolinário, Dubeux Júnior, Da Cunha and Dos Santos2021). The greater forage nutritive value is usually associated with increasing crude protein (CP) concentration in forages under SPSs compared to open pasture (Jose et al., Reference Jose, Walter and Mohan Kumar2019; Dos Santos et al., Reference Dos Santos, Junior, Dos Santos, De Andrade Lira, De Oliveira Apolinário, Costa, De Lima Coêlho, Peixôto and Da Silva Santos2020; Lima et al., Reference Lima, Dubeux, Santos, Mello, Lira, Cunha, De Freitas and Apolinário2020; Dibala et al., Reference Dibala, Jose, Gold, Hall, Kallenbach and Knapp2021). However, the effect of SPSs on neutral detergent fibre (NDF) concentration and forage digestibility is still inconsistent in the literature. Lima et al. (Reference Lima, Paciullo, Morenz, Gomide, Rodrigues and Chizzotti2019a, Reference Lima, Paciullo, Silva, Morenz, Gomide, Rodrigues, Bretas and Chizzotti2019b) and Paciullo et al. (Reference Paciullo, Fernandes, Carvalho, Morenz, Lima, Maurício and Gomide2021) observed lower herbage mass and greater CP concentration, with no differences in forage fibre constituents or digestibility in SPSs compared to monoculture pastures. The increase in CP concentration can be explained by three main mechanisms: (1) greater inorganic N availability in the soil due to greater litter deposition and N cycling, especially when legume trees are incorporated into the system (Wilson, Reference Wilson1996), (2) delay in the ontogenetic development of plants, remaining physiologically younger under shaded conditions (Neel et al., Reference Neel, Felton, Singh, Sexstone and Belesky2016; Geremia et al., Reference Geremia, Crestani, Mascheroni, Carnevalli, Mourão and Da Silva2018; Paciullo et al., Reference Paciullo, Fernandes, Carvalho, Morenz, Lima, Maurício and Gomide2021) and (3) lower tiller density and herbage mass, which reduce the competition for the inorganic N available in the soil based on the theory of N dilution and critical N concentration (Plenet and Lemaire, Reference Plénet and Lemaire1999). However, shade-grown plants usually increase stem and leaf elongation as a strategy to compensate for the light reduction (Cruz et al., Reference Cruz, Borges, Santos, Magalhães, Martuscello, Fonseca and Da Silva2021). The inconsistency in terms of NDF concentration or forage digestibility in SPSs might be associated with the intrinsic characteristics of each forage species and a trade-off effect between the increasing concentration of cell wall constituents in response to stem elongation and the slower physiological development of shaded plants.
Long-term studies (14–19 years) by Lima et al. (Reference Lima, Paciullo, Morenz, Gomide, Rodrigues and Chizzotti2019a, Reference Lima, Paciullo, Silva, Morenz, Gomide, Rodrigues, Bretas and Chizzotti2019b) and Paciullo et al. (Reference Paciullo, Fernandes, Carvalho, Morenz, Lima, Maurício and Gomide2021) demonstrated that SPSs has the potential to maintain animal performance even with reduced herbage mass and accumulation due to greater forage nutritive value and animal well-being compared to the monoculture of signalgrass. Thus, the management of SPSs (e.g. species choice, thinning, pruning, etc.) seems critical for ensuring animal products as an essential provisioning service.
De Oliveira et al. (Reference De Oliveira, Menezes, Gonçalves, De Araújo, Ramirez, Guimarães Júnior, Jayme and Lana2022) conducted a systematic review and meta-analysis of forage traits and animal performance in SPSs integrating Eucalyptus spp. and Brachiaria spp. They demonstrated that SPSs consistently reduce forage mass and forage accumulation while increasing forage CP and lignin concentration with no impact on forage digestibility. In addition, the authors observed a significant reduction in stocking rate and gain per area with large tree populations in SPSs. They concluded that SPSs with less than 99 trees/ha and greater than 28 m between tree rows can maximize forage and beef cattle performance. Lima et al. (Reference Lima, Paciullo, Morenz, Gomide, Rodrigues and Chizzotti2019a, Reference Lima, Paciullo, Silva, Morenz, Gomide, Rodrigues, Bretas and Chizzotti2019b) and Pontes et al. (Reference Pontes, Stafin, Moletta and Porfírio-da-Silva2020) also recommended low density of trees and silvicultural practices to ensure the sustainability of SPSs and greater animal production for human food supply. According to Dos Santos Neto et al. (Reference Dos Santos Neto, Da Silva, Maranhão, Cavalcante, Macedo and Cândido2023), SPSs with up to 30% woody cover can be adopted without compromising forage mass and structural characteristics of Urochloa mosambicensis, M. maximus and Pennisetum ciliare.
Biofuels and other provisioning services
In Europe, there is an increased demand for biomass production for bioenergetics purposes (Moreno et al., Reference Moreno, Franca, Correia and Godinho2014). Root-Bernstein and Jaksic (Reference Root-Bernstein and Jaksic2013) proposed restoring the ‘Espinal’ area, a silvopastoral habitat of Central Chile, with endemic species such as Acacia caven (Mol.), aiming for coal production. In the Amazon region of Brazil, the Attalea maripa (Aubl.) Mart. is a palm tree species that produces fruits with significant potential to produce biodiesel and other industrial products, with the natural occurrence contributing to the establishment of SPSs (Matos et al., Reference Matos, Rosa, Pires, Cabral, Vieira and Silva2017). At Manabí province, Ecuador, the Jatropha curcas L. crop was primarily used as live fences in SPSs and has been used for bioenergy production to increase the system profitability (Rade et al., Reference Rade, Cañadas, Zambrano, Molina, Ormaza and Wehenkel2017).
SPSs can also deliver other provisioning services, such as natural medicine and ornamental plants (Dubeux et al., Reference Dubeux, Muir, Apolinário, Nair, Lira and Sollenberger2017). Olivares-Pérez et al. (Reference Olivares-Pérez, Rojas-Hernández, Avilés-Nova, Camacho-Díaz, Jiménez-Guillén and Quiroz-Cardozo2016) indicated that non-legume species used in SPSs may be used as medicine plants, especially the C. alata Kunth and G. ulmifolia Lam. Besides coal production, the Chilean ‘Espinal’ may be managed to produce medicine plants (Root-Bernstein and Jaksic, Reference Root-Bernstein and Jaksic2013).
Supporting services
Supporting ecosystem services are those necessary to produce the other three types of services (Lamarque et al., Reference Lamarque, Quétier and Lavorel2011). Nutrient cycling (Xavier et al., Reference Xavier, Da Silva Lédo, Paciullo, Urquiaga, Alves and Boddey2014; Apolinário et al., Reference Apolinário, Dubeux, De Andrade Lira, Sampaio, De Amorim, De Miranda E Silva and Muir2016), soil fertility (Lima et al., Reference Lima, Dubeux, Santos, Mello, Lira and Cunha2018; Dibala et al., Reference Dibala, Jose, Gold, Hall, Kallenbach and Knapp2021; Moreno-Galván et al., Reference Moreno-Galván, Romero-Perdomo, Pardo-Díaz, Dávila-Mora, Castro-Rincón, Rojas-Tapias and Estrada-Bonilla2023) and BNF (Xavier et al., Reference Xavier, Da Silva Lédo, Paciullo, Urquiaga, Alves and Boddey2014; Apolinário et al., Reference Apolinário, Dubeux, Lira, Ferreira, Mello, Santos, Sampaio and Muir2015) are the primary services provided in this category.
Nutrient cycling
In SPSs, litter and animal excreta are the main paths of adding or returning nutrients to the ecosystem through microbial decomposition (Dubeux et al., Reference Dubeux, Muir, Apolinário, Nair, Lira and Sollenberger2017). The amount and spatial distribution of litter and animal manure, and consequently the nutrient cycling in the system, depends on the species, tree density, meteorological conditions, stocking method, stocking rate and location of water troughs, mineral troughs, shade and gates (Dubeux et al., Reference Dubeux, Sollenberger, Vendramini, Interrante and Lira2014; Carnevalli et al., Reference Carnevalli, Mello, Shozo, Crestani, Coletti and Eckstein2019; Dubeux and Sollenberger, Reference Dubeux, Sollenberger, Rouquette and Aiken2020).
Trees in SPSs with tap roots may intercept and absorb nutrients from deep soil horizons and recycle them to the soil surface through litter deposition (Dubeux et al., Reference Dubeux, Muir, Apolinário, Nair, Lira and Sollenberger2017; Poudel et al., Reference Poudel, Pent and Fike2024), increasing the availability of nutrients for adjacent herbaceous species. Reis et al. (Reference Reis, Lana, Maurício, Lana, Machado, Borges and Neto2010) showed that the native tree species Zeyheria tuberculosa Vell. Bar. represents considerable N, potassium (K) and calcium (Ca2+) inputs in an SPS in the Cerrado biome, Minas Gerais, Brazil. Torralba et al. (Reference Torralba, Fagerholm, Burgess, Moreno and Plieninger2016) also reported a significant positive effect of agroforestry systems on nutrient cycling and soil fertility in European systems. However, tree species deposit organic materials with different characteristics in SPSs, resulting in variations in litter quality, which may significantly impact decomposition processes, nutrient cycling and the overall soil health and productivity of the ecosystem. Additionally, the distance from legume trees can modify the litter quality and spatial distribution of N in SPSs (Dos Santos et al., Reference dos Santos, Dubeux, dos Santos, Costa, Côelho, Santos, Silva, de Oliveira, Apolinário and Coelho2024). Litter with a lower C:N ratio usually has a greater decomposition rate (Reis et al., Reference Reis, Lana, Maurício, Lana, Machado, Borges and Neto2010). The decomposition is also influenced by other chemical characteristics and environmental factors, such as soil fertility, microorganism activity and diversity and animal stocking rate (Apolinário et al., Reference Apolinário, Dubeux, De Andrade Lira, Sampaio, De Amorim, De Miranda E Silva and Muir2016). Incorporating legume trees in SPSs is an alternative to providing high-quality litter (i.e. lower C:N ratio), increasing soil nutrient mineralization, diversifying nutrient cycling and creating zones with different nutrient return rates along the pasture (Pessoa et al., Reference Pessoa, Da Cunha, De Mello, Dos Santos, Soares, Camelo, De Oliveira Apolinário, Junior and Coelho2024).
Grazing management also affects nutrient cycling in SPSs. Lower grazing intensity can lead to greater deposition and spatial distribution of litter in the system (Dubeux and Sollenberger, Reference Dubeux, Sollenberger, Rouquette and Aiken2020). However, litter accumulation due to low grazing pressure or high tree density, especially poor-quality litter, is often associated with nutrient immobilization and tillering reduction in SPSs and should be avoided (Dubeux and Sollenberger, Reference Dubeux, Sollenberger, Rouquette and Aiken2020).
Regarding animal excreta, the uneven distribution is the main challenge to improving nutrient cycling through this pathway. The excreta deposition is usually concentrated in shade or water areas, reducing the spatial distribution and efficiency of nutrient cycling (Dennis et al., Reference Dennis, Moir, Cameron, Edwards and Di2013; Dubeux et al., Reference Dubeux, Sollenberger, Vendramini, Interrante and Lira2014). Thus, the arboreous component in SPSs can be used to improve the microclimatic conditions in tropical areas, providing better shade distribution for grazing animals (Lira Junior et al., Reference Lira Junior, Fracetto, Ferreira, Silva and Fracetto2020; Dibala et al., Reference Dibala, Jose, Gold, Hall, Kallenbach and Knapp2021; De Macêdo Carvalho et al., Reference De Macêdo Carvalho, De Mello, Da Cunha, De Oliveira Apolinário, Da Silva, De Miranda Costa, De Carvalho and Dos Santos2022a, Reference De Macêdo Carvalho, De Mello, Da Cunha, De Oliveira Apolinário, Júnior, Pessoa, De Oliveira Neto and Da Silva2022b). This contributes to improving the spatial distribution of dung and urine in the pasture, generally improving pasture nutrient recycling due to the decomposition of the residues (Lima et al., Reference Lima, Dubeux, Dos Santos, De Mello, Lira and Cunha2016). de Araújo et al. (Reference de Araújo, Cláudia Rodrigues, Costa, Santos, Junior, de Jesus, Shigaki and Araujo2017) reported better distribution of dung patches in SPSs compared to palisade grass monoculture due to improved thermal comfort provided by the presence of babassu palm trees (Attalea speciosa Mart. ex Sprengel). Short grazing periods and high stocking densities can also lead to a more uniform excreta distribution and nutrient inputs (Dubeux et al., Reference Dubeux, Sollenberger, Mathews, Scholberg and Santos2007). Nitrogen deposition via animal excreta in SPSs, for example, usually ranges from 30 to 60 kg/ha, depending on several factors, such as the type of vegetation present in the SPSs, grazing intensity, management practices, soil characteristics and climatic conditions (Xavier et al., Reference Xavier, Da Silva Lédo, Paciullo, Urquiaga, Alves and Boddey2014; Lima et al., Reference Lima, Dubeux, Dos Santos, De Mello, Lira and Cunha2016; Rivera et al., Reference Rivera, Chará and Barahona2018; Sarabia-Salgado et al., Reference Sarabia-Salgado, Alves, Boddey, Urquiaga, Galindo, Flores-Coello, Santos, Jimenez-Ocampo, Ku-Vera and Sánchez2023).
Therefore, there is a trade-off between increasing litter deposition and reducing excreta deposition when a lower stocking rate and lower forage removal are adopted. Similarly, increasing grazing intensity increases the nutrient return via excreta while reducing litter deposition. In overgrazed areas, the low forage allowance and reduced animal intake can restrict even the excreta deposition. Furthermore, litter accumulation in ungrazed areas or excreta accumulation in overgrazed areas also compromises nutrient cycling and system productivity.
In summary, the herbage allowance and stocking rate adjustments to balance nutrient return via litter and animal manure seem to be the primary strategy to promote nutrient cycling in SPSs. The decomposition rate is greater for animal excreta compared to litter. However, animal excreta are also more susceptible to nutrient loss by gas emissions, which make both pathways essential for nutrient cycling and mineralization (Dubeux and Sollenberger, Reference Dubeux, Sollenberger, Rouquette and Aiken2020).
Soil fertility and quality
SPSs can improve soil fertility, mainly by correcting soil acidity. Reis et al. (Reference Reis, Lana, Maurício, Lana, Machado, Borges and Neto2010) reported that the SPSs formed by the native species Z. tuberculosa Vell. Bur. and the tropical grass Urochloa brizantha Stapf. cv. Marandu, at the Brazilian Cerrado biome, improved soil fertility because of the acidity correction by the high Ca2+ and magnesium (Mg2+) concentrations in the litter while maintaining the soil N, P and K concentrations. However, the authors estimated a density of 160 trees/ha in their study. In systems with lower tree density, the amount of litter deposited might be not enough to increase Ca2+ and Mg2+ concentrations in the soil, while in systems with higher tree density, the forestry component may become a sink of nutrients in the soil.
In Central America, Hoosbeek et al. (Reference Hoosbeek, Remme and Rusch2018) observed that isolated trees in pastures increased contents of C, N and P in the topsoil. The authors attributed the greater soil fertility to the higher above- and below-ground litter inputs from trees. Casals et al. (Reference Casals, Romero, Rusch and Ibrahim2014) also reported that SPSs increased soil K and Ca2+ in the dry tropics compared to open pastures. Accordingly, Lira Junior et al. (Reference Lira Junior, Fracetto, Ferreira, Silva and Fracetto2020) observed that tree legumes incorporated into signalgrass pasture increased soil organic matter (SOM), another essential pool of nutrients in the soil. However, its effects on soil fertility may be affected by tree species, density, age, size and prevalent environmental conditions.
In addition to fertility, SPSs can also improve general soil quality through improvements in the soil's physical, chemical and biological attributes (Barros et al., Reference Barros, Fracetto, Fracetto, Júnior, Araújo and Lira Junior2018; Lima et al., Reference Lima, Dubeux, Santos, Mello, Lira and Cunha2018; Lira Junior et al., Reference Lira Junior, Fracetto, Ferreira, Silva and Fracetto2020). The benefits of SPSs on soil health can be associated with greater water infiltration rate by deeper roots, greater soil microporosity and aeration due to greater litter deposition, and favourable microclimate, including moisture, temperature and vegetation cover to increase soil microfauna (Lima et al., Reference Lima, Dubeux, Santos, Mello, Lira and Cunha2018).
Murgueitio et al. (Reference Murgueitio, Calle, Uribe, Calle and Solorio2011) evaluated microclimate conditions generated by trees in SPSs and their impact on soil physic-hydric characteristics in the tropics. They reported that the shade provided by trees reduced average surface temperatures by up to 3°C and evapotranspiration by 1.8 mm/day. Dibala et al. (Reference Dibala, Jose, Gold, Hall, Kallenbach and Knapp2021) studied three cultivars of guinea grass (Panicum maximum cvs. Massai, Mombaça and Tanzânia) under open, moderate and dense tree canopies (mixed native from Panama species N-fixing and non-N-fixing). They reported bulk density reduction under tree canopies and significant soil quality improvements after reforestation, as tree roots create macropores in the soil favourable to water infiltration. Additionally, SPSs with shrub legumes can affect the chemical fractions of SOM and increase soil C and N stocks (Lima et al., Reference Lima, Dubeux, Santos, Mello, Lira and Cunha2018; Lira Junior et al., Reference Lira Junior, Fracetto, Ferreira, Silva and Fracetto2020).
Barros et al. (Reference Barros, Fracetto, Fracetto, Júnior, Araújo and Lira Junior2018) reported that the structures of the total and ammonium-oxidizing bacterial communities were influenced by the introduction of tree legumes, possibly by its impact on soil chemical attributes. A study conducted in Colombia showed that the implementation of a 9-year-old SPSs consisting of a tree (Alnus acuminata Kunth), two shrubs (Sambucus peruviana Kunth and Sambucus nigra L.) and Kikuyu grass [Cenchrus clandestinus (Hochst. ex Chiov.) Morrone] increased bacterial diversity in the soil and facilitated the absorption of phosphorus by plants (Moreno-Galván et al., Reference Moreno-Galván, Romero-Perdomo, Pardo-Díaz, Dávila-Mora, Castro-Rincón, Rojas-Tapias and Estrada-Bonilla2023).
The availability of nutrients in the soil in SPSs results from a complex interaction of several physical, chemical and biological factors. In the tropical drylands of Colombia, Martínez et al. (Reference Martínez, Cajas, León and Osorio2014) indicated that trees in SPSs increased or maintained soil pH values and nutrient availability (P, K and Ca) relative to pastures with only grasses. Lima et al. (Reference Lima, Dubeux, Santos, Mello, Lira and Cunha2018) reported that the exchangeable Ca2+ in the soil was greater for SPSs with arboreal legume and signalgrass (average of 3.1 mmol/dm3) than grass monoculture (2.0 mmol/dm3). The authors also report that the exchangeable Na in the soil was greater for SPSs with ‘sabiá’ arboreal legume (0.3 mmol/dm3) in comparison to SPSs with gliricidia and grass monoculture (0.1 mmol/dm3) due to greater accumulation of ‘sabia’ litter.
Nitrogen biological fixation
Including tree legumes in SPSs may increase forage production due to BNF compared to unfertilized monoculture pastures (Freitas et al., Reference Freitas, Sampaio, Santos and Fernandes2010). The grass plants benefit from BNF via legume root and nodule degradation, decomposition of legume litter, nutrients recycled through excreta and, to a smaller extent, root nitrogenous exudates and mycorrhizal fungi mycelial networks (Apolinário et al., Reference Apolinário, Dubeux, Lira, Ferreira, Mello, Santos, Sampaio and Muir2015). However, the spacing between trees and tree density will determine the canopy light interception and forage production in SPSs.
Dubeux et al. (Reference Dubeux, Muir, Nair, Sollenberger, Siqueira da Silva and Mello2015) reported BNF in tree and shrub forage legumes varying from 24 to 304 kg N/ha/year in different species and countries, although the typical rate varies from 50 to 150 kg N/ha/year (Dubeux et al., Reference Dubeux, Muir, Apolinário, Nair, Lira and Sollenberger2017). Ledgard and Steele (Reference Ledgard and Steele1992) also reported a wide range of BNF in grass-legume pastures established worldwide (13–682 kg N/ha/year). The wide range of BNF rates of legume species found in the literature might be attributed to differences in estimation methods and to the variability of several factors affecting BNF such as soil fertility, water content, grazing management and climatic conditions (Rosenstock et al., Reference Rosenstock, Tully, Arias-Navarro, Neufeldt, Butterbach-Bahl and Verchot2014; Dubeux et al., Reference Dubeux, Muir, Apolinário, Nair, Lira and Sollenberger2017). General research findings indicate that including forage legumes in SPSs may provide a sufficient amount of N via BNF to maintain the productivity of the pastures (Xavier et al., Reference Xavier, Da Silva Lédo, Paciullo, Urquiaga, Alves and Boddey2014).
BNF is one of the most desirable characteristics in forage legumes, and it can vary according to species, environmental factors and management (Dubeux et al., Reference Dubeux, Muir, Apolinário, Nair, Lira and Sollenberger2017). Tropical forests present a large diversity and participation of legume species with the potential of symbiosis with N-fixing bacteria (Da Silva et al., Reference Da Silva, De Freitas, Costa, Fernandes-Júnior, Martins, Santos, Menezes and Sampaio2017). Native species often obtain more than 80% of N from symbiotic fixation (Freitas et al., Reference Freitas, Sampaio, Santos and Fernandes2010).
Freitas et al. (Reference Freitas, Sampaio, Santos and Fernandes2010) studied the BNF of tree forage legumes from Brazilian Caatinga. They indicated that Mimosa tenuiflora (Willd.) Poir., Mimosa arenosa (Willd.) Poir. and Piptadenia stipulacea (Benth.) Ducke are species with a great capacity for symbiosis with N-fixing bacteria. However, it has been reported that the arboreal tropical legume Bauhinia cheilantha (Bong.) did not ‘fix’ atmospheric N probably because the bacteria cannot infect these plants or due to the low efficiency of N-fixation as related to soil nutrient content (Da Silva et al., Reference Da Silva, De Freitas, Costa, Fernandes-Júnior, Martins, Santos, Menezes and Sampaio2017). Thus, evaluating the potential of N-fixation in tropical legumes is important because it may impact the expected ecosystem service from the system.
Regulating services
Ecosystem regulation services are related to mitigating present and future environmental impacts (Guerra and Pinto-Correia, Reference Guerra and Pinto-Correia2016). MEA (2005) pointed out carbon sequestration, mitigation of GHG emissions, control of soil erosion, climate regulation and pollination as major regulating services. Additionally, the control of pests and diseases (Enriquez-Hidalgo et al., Reference Enriquez-Hidalgo, Gilliland and Hennessy2016; Alonso-Amaro et al., Reference Alonso-Amaro, Lezcano-Fleires and Suris-Campos2019), biodiversity (Tobar and Ibrahim, Reference Tobar and M2009; Olival et al., Reference Olival, De Oliveira, Sais and De Souza2022), residue absorption and control of water quality and availability (Lin et al., Reference Lin, Lerch, Goyne and Garrett2011) can be included in these ecosystem services.
Carbon sequestration and mitigation of GHGs
SPSs play a crucial role in atmospheric carbon sequestration and mitigation of GHG emissions (Montagnini et al., Reference Montagnini, Ibrahim and Murgueitio2013). According to Luedeling et al. (Reference Luedeling, Kindt, Huth and Koenig2014), agroforestry systems offer greater opportunities for C sequestration in biomass and soil than monocultural systems. This is generally attributed to greater litter deposition, nutrient cycling and C stabilization due to deeper root systems within agroforestry systems (Nair, Reference Nair2014a, Reference Nair2014b; Chatterjee et al., Reference Chatterjee, Nair, Chakraborty and Nair2018; Pinheiro et al., Reference Pinheiro, Nair, Nair, Tonucci and Venturin2021). The tree component in SPSs presents a high capacity to sequester C and produce a decomposition-resistant litter, which turns this integrated system into a more efficient system in terms of carbon fixation (Giustina et al., Reference Giustina, Carnevalli, Romano, Antonio and Eckstein2017).
Montagnini et al. (Reference Montagnini, Ibrahim and Murgueitio2013) indicated that the C sink from trees in SPSs varied from 0.42 to 92 Mg/ha, while the soil C sink varied from 58 to 140 Mg/ha. The same authors reported that the C sequestration in the system may vary from 0.08 to 4.6 Mg/ha/year. The wide variation reflects the heterogeneity of SPSs, which differs in its design, species and local conditions. In southwest Nicaragua, the SPSs composed of G. ulmifolia Lam. and C. alata Kunth represented an organic C sink of 70 Mg/ha while pastures under full sun had 45 Mg/ha (Hoosbeek et al., Reference Hoosbeek, Remme and Rusch2018). The total storage of C aboveground on SPSs, including the litter accumulation, can reach 5–7.5 times greater than pastures under full radiation (Aryal et al., Reference Aryal, Gómez-González, Hernández-Nuriasmú and Morales-Ruiz2019). Corroborating this statement, Sarto et al. (Reference Sarto, Borges, Sarto, Rice and Rosolem2020) observed that SPSs stored 68% more C compared to a monoculture pasture due to the C accumulated in the aboveground tree biomass. Besides the C stocks in the above- and below-ground biomass of SPSs, perennial species with deeper root systems (e.g. arboreous species) might contribute to greater C stabilization and, consequently, C sequestration, compared to herbaceous species predominant in monoculture systems (Haile et al., Reference Haile, Nair and Nair2010; Howlett et al., Reference Howlett, Mosquera-Losada, Nair, Nair and Rigueiro-Rodríguez2011; Nair, Reference Nair2014a, Reference Nair2014b; Hoosbeek et al., Reference Hoosbeek, Remme and Rusch2018). Using stable isotope signatures, Haile et al. (Reference Haile, Nair and Nair2010) observed that the C found in the deeper soil profile in Florida SPSs was derived from the tree component, also suggesting greater contributions of C3 plants (e.g. legume trees) than C4 grasses to C sequestration in SPSs. Sarto et al. (Reference Sarto, Borges, Sarto, Rice and Rosolem2020) also observed a greater contribution of C3 trees in SPSs to the C stocks in deep layers (0.6–1.0 m) compared to a monoculture pasture (C4). The greater C stabilization in deeper layers is likely explained by the predominancy of microaggregates, with a greater capacity to protect and stabilize the organic matter compared to top layers, with predominancy of macroaggregates (Pinheiro et al., Reference Pinheiro, Nair, Nair, Tonucci and Venturin2021). In addition, Tonucci et al. (Reference Tonucci, Nair, Nair, Garcia and Bernardino2011) and Pinheiro et al. (Reference Pinheiro, Nair, Nair, Tonucci and Venturin2021) suggested the macroaggregate proportion in the soil as a good indicator of C storage potential in SPSs due to the effect on microaggregate formation and protection. The lower soil disturbance compared to treeless areas may also enhance the formation of microaggregates in SPSs (Angers and Chenu, Reference Angers, Chenu, Lal, Kimble, Follett and Stewart2018).
Chatterjee et al. (Reference Chatterjee, Nair, Chakraborty and Nair2018), in a meta-analysis of 78 studies conducted in different agroecological regions around the world, concluded that SPSs increased C stocks (up to 1 m depth) in arid, semi-arid and Mediterranean regions while decreasing in temperate and lowland humid tropics regions, suggesting that the potential of SPSs to enhance C sequestration is site dependent. In addition, the same authors concluded that SPSs aged between 10 and 20 years are significantly more effective in improving soil C stocks than younger systems. Filho et al. (Reference Filho, De Oliveira, De Souza Barros, Dos Santos and De Oliveira2024) also demonstrated the potential of SPSs (shading level of 25%) to increase soil C stocks (up to 1 m depth) compared to natural vegetation or intensive monoculture pasture in the Cerrado-Amazon ecotone.
On the other hand, Tonucci et al. (Reference Tonucci, Nair, Nair, Garcia and Bernardino2011) found greater C stock up to 1 m depth under pasture compared to other land uses, including SPSs, in Brazilian Cerrado. The authors justified the greater C stock in pastures due to the higher bulk density associated with greater animal trampling in pastures. However, the unequal soil mass might be a bias source in C stock comparisons of soils with different bulk densities (Von Haden et al., Reference Von Haden, Yang and DeLucia2020; Fowler et al., Reference Fowler, Basso, Millar and Brinton2023; Peng et al., Reference Peng, Chahal, Hooker and Van Eerd2024). Seddaiu et al. (Reference Seddaiu, Porcu, Ledda, Roggero, Agnelli and Corti2013) also reported lower C stock in SPSs than monoculture pastures. According to Jobbágy and Jackson (Reference Jobbágy and Jackson2000), the rhizodeposition turnover in pastures is higher than in areas under trees, increasing C stocks. Corroborating previous studies, Pinheiro et al. (Reference Pinheiro, Nair, Nair, Tonucci and Venturin2021) found lower C stock in SPSs established with Eucalyptus hybrid than in open pasture. The same authors highlighted the importance of considering other tree species to establish SPSs in Brazilian Cerrado, suggesting using native N-fixing legumes to increase C stocks while exploring the known biodiversity of the Cerrado biome. This would also contribute to the world's biodiversity conservation goals (Pinheiro and Hunt, Reference Pinheiro and Hunt2020). Most of the SPSs in Brazilian Cerrado involve non-native Eucalyptus trees, which sometimes do not enhance soil C stocks due to the low leaf decomposition rate and root–shoot ratio (Pinheiro et al., Reference Pinheiro, Nair, Nair, Tonucci and Venturin2021).
Despite the often-reported potential of SPSs to mitigate GHG emissions by C sequestration, SPSs can also contribute to GHG emissions from litter decomposition, excreta deposition and enteric fermentation. In addition, Chatterjee et al. (Reference Chatterjee, Nair, Chakraborty and Nair2018) suggested that the potential of SPSs in mitigating GHG emissions through C sequestration could be more relevant in tropical regions, where the soils are usually C-depleted, while soils in temperate regions are usually C-saturated. Nair (Reference Nair2011), in a literature review about C stocks in SPSs, also concluded that the C sequestration potential is affected by climatic conditions, soil type and plant species. De Abreu et al. (Reference De Abreu, De Freitas, Santana, De Almeida Barbosa, Santos, Santos, Sanglard and Frazão2020) highlighted the importance of correct soil sampling and measurements to evaluate soil C variation in SPSs. Thus, the inconsistency found in the literature may be related to different climatic conditions, species, system design, soil type, management practices and methodologies used to access soil C. Compiled studies from 2010 to 2024 demonstrate the global variability in species and system design used for SPS adoption that could justify the inconsistency in the literature regarding the impact of SPSs on GHG emissions (Table 2).
Table 2. Compiled studies from 2010 to 2024 exemplifying the global variability in species and system design for SPS adoptiona
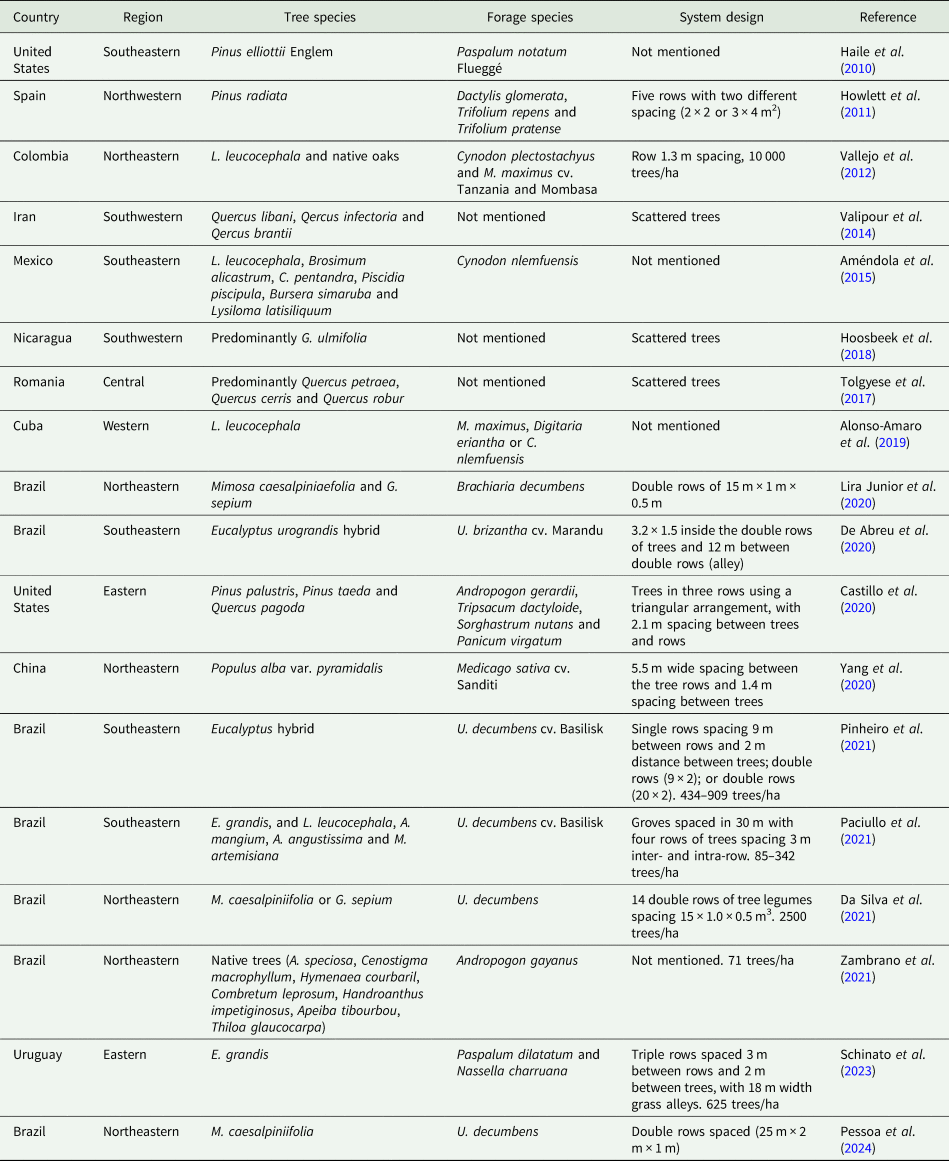
a Several other options of species and system design are worldwide available for SPS adoption.
An approach to minimizing GHG emissions in SPSs involves feeding the cattle high-quality forage that contributes to adequate dry matter intake (Montagnini et al., Reference Montagnini, Ibrahim and Murgueitio2013). The integration of leguminous trees and forages with a considerable condensed tannin concentration can help to reduce animal methane (CH4) emissions in SPSs (Dubeux et al., Reference Dubeux, Muir, Apolinário, Nair, Lira and Sollenberger2017). Tannins are phenolic compounds that interfere with digestion by forming complexes with protein molecules, reducing the activity of ruminal microorganisms, specifically methanogenic microorganisms (Naumann et al., Reference Naumann, Tedeschi, Zeller and Huntley2017). In SPSs with shrubs forage legume L. leucocephala (Lam.) De Wit, the annual CH4 emission per animal may be reduced by 38% (Naranjo et al., Reference Naranjo, Cuartas and Murgueitio2012). In addition, legume trees with high contents of condensed tannins, such as L. leucocephala (Lam.) De Wit could potentially decrease nitrous oxide (N2O) emissions in SPSs by reducing nitrogen excretion in urine through decreased ruminal protein degradation (Van Cleef et al., Reference Van Cleef, Dubeux, Ciriaco, Henry, Ruiz-Moreno, Jaramillo, Garcia, Santos, DiLorenzo, Vendramini, Naumann and Sollenberger2022). Naranjo et al. (Reference Naranjo, Cuartas and Murgueitio2012) highlighted that monoculture pastures, both degraded and improved, subjected to full sun radiation, were identified as significant sources of GHG, with a negative net balance of 3153 and 3259 kg of equivalent CO2/ha/year. On the other hand, the SPS was considered a sink of GHG, with a positive net balance varying from 8800 to 26 565 kg of equivalent CO2/ha/year. Additionally, the N supply from legume trees can enhance diet nutritive value in SPSs, reducing the production cycle and animal product emissions per unit (Lüscher et al., Reference Lüscher, Mueller-Harvey, Soussana, Rees and Peyraud2014; Rivera-Herrera et al., Reference Rivera Herrera, Molina-Botero, Chará, Murgueitio-Restrepo and Barahona Rosales2017; Homem et al., Reference Homem, Borges, De Lima, Guimarães, Spasiani, Ferreira, Meo-Filho, Berndt, Alves, Urquiaga, Boddey and Casagrande2024). Furthermore, some of the enteric fermentation gases produced by grazing animals in SPSs might be used by trees and grasses, increasing tree biomass production, resulting in more rapid grass regrowth and reducing even more GHG emissions (Zambrano et al., Reference Zambrano, Edvan, Oliveira, Da Costa Araujo, Costa, Da Silva, Barros, De Sousa Reis and Dias-Silva2021).
Despite the potential benefits, it is important to mention that including N-fixing species in SPSs may contribute to other paths of GHG emissions in the system. Bretas et al. (Reference Bretas, Paciullo, Alves, Martins, Cardoso, Lima, Rodrigues, Silva and Chizzotti2020) observed greater N2O and CH4 fluxes in SPSs compared to monoculture pasture and justified it due to greater N concentration in the manure from animals grazing in SPSs, greater inorganic-N availability in SPSs (litter and excreta richer in N), and maintenance of favourable soil conditions (e.g. soil moisture, microbial activity, etc.) for a more extended period in SPS systems. On the other hand, Rivera et al. (Reference Rivera, Villegas, Chará, Durango, Romero and Verchot2024) showed lower CH4 and N2O emissions in SPSs compared to monoculture pastures.
The site preparation required for SPS establishment should also be considered because, in some cases, land management can result in C loss. Most of the C in biodegradable form is likely to be lost by soil disturbance during land use changes (Nair, Reference Nair2011). Thus, the adoption of SPSs as a strategy to mitigate GHG emissions should be carefully evaluated. Generally, well-managed SPSs can offset the N2O and CH4 emissions from soil and animal excreta by the C sequestration. Naranjo et al. (Reference Naranjo, Cuartas and Murgueitio2012) estimated the balance of GHG in an SPS and concluded that the C sequestration in the system compensated the emitted N2O. Therefore, the tree density, species, soil preparation, grazing management, litter accumulation, forage accumulation, herbage allowance, animal performance and GHG emissions are directly related and should be considered as a system to optimize the regulating services in SPSs.
Control of soil erosion
Soil erosion is a natural process, often accelerated by human activities, which involves the progressive removal and loss of soil particles from the surface. This removal occurs mainly due to the action of water and wind, resulting in soil degradation and a reduction in its quality (Lal, Reference Lal2001; Issaka and Ashraf, Reference Issaka and Ashraf2017). The trees in the SPSs present ecological functions, including soil protection, contributing to reducing the adverse effects of wind and water percolation. According to Nahed-Toral et al. (Reference Nahed-Toral, Valdivieso-Pérez, Aguilar-Jiménez, Cámara-Cordova and Grande-Cano2013), the trees associated with pastures can contribute to mitigating soil erosion through their radicular systems. Bayala et al. (Reference Bayala, Sanou, Teklehaimanot, Kalinganire and Ouédraogo2014) also reported the potential of extensive SPSs known as parklands in West Africa to reduce soil erosion and mitigate climate. These systems are characterized by pastures grown under scattered trees and shrubs that provide soil cover, reduce soil erosion and offer green fodder.
One of the main benefits of trees on agroecosystems is soil conservation, maintaining or increasing SOM, and improving soil structure, porosity and water-holding capacity. Trees may contribute to reducing surface runoff, increasing water infiltration into the soil (Benegas et al., Reference Benegas, Ilstedt, Roupsard, Jones and Malmer2014; Lima et al., Reference Lima, Dubeux, Santos, Mello, Lira and Cunha2018) and improving water uptake by plants, including the water in the deeper soil layers, which may contribute to increasing total water consumption (Pezzopane et al., Reference Pezzopane, Bosi, Nicodemo, Santos, Cruz and Parmejiani2015; Bosi et al., Reference Bosi, Pezzopane and Sentelhas2020). These associated factors might contribute to improving plant growth and soil cover, reducing soil erosion.
However, all the above-mentioned benefits rely on soil cover. If incompatible species, high density of trees, intense shading and high grazing intensity are adopted, the forage production might reduce significantly, exposing the soil to an erosive process. Thus, the species choice, shading management and grazing management will determine the persistency of the forage species in SPSs and the potential of soil cover over time, playing a key role in soil erosion control by SPSs.
Microclimate regulation
The tree shading may influence the pastoral microenvironment. Tree canopies can protect from winds and reduce soil surface temperature and canopy evapotranspiration, which may increase soil water availability (Benegas et al., Reference Benegas, Ilstedt, Roupsard, Jones and Malmer2014; Dibala et al., Reference Dibala, Jose, Gold, Hall, Kallenbach and Knapp2021). Pezzopane et al. (Reference Pezzopane, Bosi, Nicodemo, Santos, Cruz and Parmejiani2015) observed that rows of native trees on the SPSs reduced 46% of wind speed and changed the incident pattern of photosynthetic active radiation on pastures (reduction of about 40%). In SPSs, the temperatures can be 2–5°C lower than in pastures under full solar radiation (Murgueitio et al., Reference Murgueitio, Calle, Uribe, Calle and Solorio2011). In Mexico, the average temperatures of integrated systems were reduced by 8.6°C compared to traditional pastures (Cardona et al., Reference Cardona, Ramírez, Morales, Murgueitio, Orozco, Vera, Sánchez, Flores, Sánchez and Rosales2014). In the Southeastern USA, Castillo et al. (Reference Castillo, Tiezzi and Franzluebbers2020) described the potential of different tree species to mitigate changes in temperature, relative humidity and temperature–humidity index in SPSs. Frota et al. (Reference Frota, Carneiro, Pereira, Berndt, Frighetto, Sakamoto, Moreira Filho, Cutrim Júnior and Carvalho2017), evaluating an SPS with a density of 67 Babaçu (A. speciosa Mart.) trees/ha in the Brazilian Amazon, estimated a shaded area of 26% of the total study area. According to Paciullo et al. (Reference Paciullo, De Castro, Gomide, Maurício, Pires, Müller and Xavier2011), moderate shade level (up to 30–40%) does not affect signalgrass growth in SPSs. Similarly, Cruz et al. (Reference Cruz, Santos, Silva, Ferreira, Magalhães, Martuscello and Fonseca2020) observed forage peanut (A. pintoi) adaptation up to 45% of shade level. Dos Santos Neto et al. (Reference Dos Santos Neto, Da Silva, Maranhão, Cavalcante, Macedo and Cândido2023) also suggested U. mosambicensis, M. maximus and P. ciliare adaptation up to 30% of shade level in an SPS in the Brazilian semi-arid region. Besides, shaded pastures may also increase soil moisture because of the lower evapotranspiration or maintain the soil moisture for longer periods after rain events due to the lower soil and air temperature (Bretas et al., Reference Bretas, Paciullo, Alves, Martins, Cardoso, Lima, Rodrigues, Silva and Chizzotti2020).
Vieira et al. (Reference Vieira, Pilatti, Czekoski, Fonsêca, Herbut, Angrecka, De Souza Vismara, De Paulo Macedo, Dos Santos and Paśmionka2021) compared the thermal environment as well as the bio-thermal and behavioural responses of grazing animals under an SPS with 23% shade level and open pastures (full sun). Regarding the thermal environment, the authors observed lower air temperature, ground surface temperature, black globe-humidity index and radiant heat load in SPSs compared to open pastures. Regarding the animal responses, lower respiratory rates, rectal temperature and hair coat surface temperature were observed. Additionally, animals in SPSs presented greater grazing and walking time, while reducing the resting time. These findings suggest higher thermal comfort of animals in SPSs compared to open pastures and might impact animal performance and system productivity. Similarly, changes in the thermal environment may also affect the herbage mass and forage nutritive value as previously discussed.
Additionally, the above-mentioned potential of SPSs to increase C sequestration and mitigate GHG emissions also contributes to climate regulation by reducing the global warming associated with significant global climate changes.
Residue absorption and control of water quality and availability
Nutrient losses, livestock residues, use of agricultural defensives, bacteria and soil and water protozoa contamination are concerns due to their impact on water quality. Lin et al. (Reference Lin, Lerch, Goyne and Garrett2011) reported that vegetation may reduce 58–72% of the transport of dissolved herbicides linked to sediments and veterinary antibiotics in the surface flow. According to the authors, tall fescue (Festuca arundinacea Scherb.) can be an efficient barrier to reduce the transportation of dissolved tylosin and enrofloxacin in the soil. Additionally, Moreno et al. (Reference Moreno, Franca, Correia and Godinho2014) and Torralba et al. (Reference Torralba, Fagerholm, Burgess, Moreno and Plieninger2016) pointed out the capacity of the deep root system of trees to absorb nutrients from deeper horizons, which reduces the nutrient lixiviation in SPSs. The residue absorption from the high aboveground biomass and deep roots from trees might contribute to improving the water quality in SPSs.
Incorporating trees in livestock systems can also enhance infiltration rates and reduce runoff losses, improving soil water storage and availability (Nair et al., Reference Nair, Kumar, Nair, Nair, Kumar and Nair2021). However, Pezzopane et al. (Reference Pezzopane, Bosi, Nicodemo, Santos, Cruz and Parmejiani2015) and Bosi et al. (Reference Bosi, Pezzopane and Sentelhas2020) reported that soil water availability was reduced in sampling sites near the tree rows, mainly due to the deep roots of the trees, especially during the dry season. Competitive interaction may be viewed more as a disservice rather than a benefit (Nair et al., Reference Nair, Kumar, Nair, Nair, Kumar and Nair2021). Thus, tree spacing, choice of species, topography, silvicultural practices and climate conditions may be the key factors driving soil water availability in SPSs. Further studies are required to define an optimal tree density and best management practices in each specific SPS to maximize soil water storage, avoiding the risk of depletion in stored water (Ilstedt et al., Reference Ilstedt, Bargués Tobella, Roméo, Bayala, Elke, Nyberg, Sanou, Benegas, Murdiyarso, Laudon, Sheil and Malmer2016; Dibala et al., Reference Dibala, Jose, Gold, Hall, Kallenbach and Knapp2021).
Pollination and biodiversity
Insects are pollinators often present in almost all ecosystems. The level of presence may indicate environmental impacts. Livestock intensification, including using nitrogen fertilizers and excessive defoliation, has been considered a critical factor affecting the population of pollinators essential to sustaining food and feed production (Dubeux et al., Reference Dubeux, Muir, Apolinário, Nair, Lira and Sollenberger2017). However, increasing plant diversity affects plant–pollination interactions by changing the environmental conditions, botanical composition and flower availability (Centeno-Alvarado et al., Reference Centeno-Alvarado, Lopes and Arnan2023). In a global review, Centeno-Alvarado et al. (Reference Centeno-Alvarado, Lopes and Arnan2023) concluded that agroforestry systems, including SPSs, promote pollination services compared to conventional systems.
Auad et al. (Reference Auad, Resende, Da Silva and Das Graças Fonseca2012) collected and identified 5841 species distributed into 11 families of the Hymenopter order in a pasture area of U. decumbens under SPSs in Minas Gerais State, Brazil. Among them, species from Formicidae, Ichneumonidae, Vespidae, Brachonidae, Chalcididae, Pompilidae, Apidae and Sphecidae families were frequently collected. Species from these families present the potential to disperse seeds and pollinate flowers, indicating the potential of SPSs for biodiversity conservation. Additionally, biodiversity may be used as a biological indicator. In the western region of Cuba, Alonso-Amaro et al. (Reference Alonso-Amaro, Lezcano-Fleires and Suris-Campos2019) made an ecological catalogue of the entomofauna and weed species present on SPSs. The authors reported 34 weeds and 79 insect species. The African bee (Apis mellifera L.) stood out among the four catalogued pollinators. In India, Yadav et al. (Reference Yadav, Gendley, Sahu, Patel, Chandraker and Dubey2019) reported that SPS areas increased the number and variety of native bird species that enhance environmental services related to pollination. The number of bird species increased by 200%, and a significant increase in butterflies occurred.
Extensive livestock farming is one of the leading causes of deforestation and loss of biodiversity in agricultural frontier regions in Brazil (Olival et al., Reference Olival, De Oliveira, Sais and De Souza2022). The tree component in SPSs may provide many gradients in light radiation, nutrients, soil moisture, herbage mass and refuge to different species, even with certain modification levels caused by grazing animals (Moreno et al., Reference Moreno, Franca, Correia and Godinho2014). This creates a habitat mosaic, which is a critical factor for the increase in biodiversity in these ecosystems.
In the Central Region of Costa Rica, Tobar and Ibrahim (Reference Tobar and M2009) reported 2782 insects from 75 butterfly species in SPSs with live fences. They reported most frequent species were Anartia fatima Fabricius, Eurema daira Godart, Eurema nise Cramer, Hermeuptychia hermes Fabricius, Junonia evarete Cramer and Phoebis philea Linnaeus. According to the authors, the SPSs supported 56% of the total species observed in secondary and riparian forests, indicating that it may play an essential role in butterfly conservation in livestock areas. Moreno et al. (Reference Moreno, Franca, Correia and Godinho2014) reported greater species richness for vessel plants, bees, spiders and worms in SPSs compared to traditional pastures in the Iberia Dehesas, Spain. Extensive SPSs from western Africa (parklands) are also reported as biodiverse ecosystems with a high potential for biodiversity conservation due to the wide diversity of wood species, habitats and resources for bird species (Muruts and Birhane, Reference Muruts and Birhane2018).
Studies have shown that small farmers can also contribute to reintroducing native tree species and native biodiversity into pastures in SPSs (Olival et al., Reference Olival, De Oliveira, Sais and De Souza2022). In addition, SPSs may provide conditions to maintain a diversity of amphibians, reptiles, fish, aquatic arthropods, molluscs and other organisms that can find habitat and resources in these systems. Overall, SPSs are potential alternatives to contribute to biodiversity increase and ecosystem stability due to the rich diversity of pollinators in response to the integration between at least two different plant species and functional groups.
Control of pests and diseases
If adequately managed, SPSs may also contribute to increasing wildlife diversity, which has the potential to control pests and diseases. The presence of the animals in pastures throughout the year associated with the high forage allowance and great nutritive value may contribute to the acquisition of resistance to internal and external parasites due to possible improvement of nutrition and immune response (Murgueitio et al., Reference Murgueitio, Calle, Uribe, Calle and Solorio2011; Cardona et al., Reference Cardona, Ramírez, Morales, Murgueitio, Orozco, Vera, Sánchez, Flores, Sánchez and Rosales2014). It has been reported that SPSs may contribute to reducing the internal parasites by 40% due to possible break in the parasite life cycle when associated with intermittent stocking and effects of secondary metabolites from tree species like L. leucocephala (Lam.) De Wit (Cardona et al., Reference Cardona, Ramírez, Morales, Murgueitio, Orozco, Vera, Sánchez, Flores, Sánchez and Rosales2014). It might occur due to substances such as tannins and alkaloids in its leaves (Cardona et al., Reference Cardona, Ramírez, Morales, Murgueitio, Orozco, Vera, Sánchez, Flores, Sánchez and Rosales2014). Additionally, the high aboveground biomass in intensively managed SPSs can favour predators such as birds, ants and fungi involved in the biological control of tick populations (Murgueitio et al., Reference Murgueitio, Calle, Uribe, Calle and Solorio2011).
Enriquez-Hidalgo et al. (Reference Enriquez-Hidalgo, Gilliland and Hennessy2016) evaluated the effect of tannin-rich plants on the control of gastrointestinal nematodes in zebu cows grazing in subtropical SPSs. They reported more eggs per g of faeces were found in cows in the conventional pasture system than in SPSs. The authors justified this effect by several environmental changes in the survivability of free-living parasite stages in SPSs and higher tannin concentration of legume trees, suggesting the potential of SPSs for controlling parasitic nematode infection in bovines. The main gastrointestinal nematodes identified in culture for both systems were Oesophagostomum spp., Haemonchus contortus, Trichostrongylus and Cooperia spp. Corroborating that study, Flota-Bañuelos et al. (Reference Flota-Bañuelos, Rivera-Lorca and Candelaria-Martinez2019) reported that sheep consuming more L. leucocephala in SPSs had lower parasite loads and greater haematocrit contents compared to animals grazing in a monoculture system due to the high iron (Fe) concentration in this legume, promoting accelerated growth; increasing resistance to infection and absence of anaemia (reflected in the haematocrit). The same authors also mentioned the presence of plant secondary metabolites as an additional benefit.
Alonso-Amaro et al. (Reference Alonso-Amaro, Lezcano-Fleires and Suris-Campos2019) reported 34 weed species in an SPS of the West Region from Cuba. The authors indicated that five species are considered able to host insects, and 27 of these insects may provide benefits for the SPSs (11 predators, 12 parasites and four pollinators). Among them, in the first group, three ladybugs and two beetles were identified as bioregulators of the Heteropsylla cubana Crawford, the primary pest of L. leucocephala (Lam.) De Wit. In the second group, many limestone wasps are considered regulators of fall armyworm [Spodoptera frugiperda (J. E. Smith)], an essential pest for agricultural fields.
Despite the mentioned benefits of SPSs to control pests and diseases in agricultural systems, some might be related to grass or tree species. Most revised studies demonstrated the benefits of L. leucocephala (Lam.) De Wit., requiring further studies with different tree species and system designs for better understanding.
Cultural services
Cultural ecosystem services refer to the non-material benefits that individuals derive from nature, encompassing spiritual, aesthetic, educational and recreational values (MEA, 2005; Kosanic and Petzold, Reference Kosanic and Petzold2020). SPSs may provide a variety of cultural ecosystem services that enhance the social and cultural fabric of communities, extending beyond economic and ecological benefits. These systems are essential for sustainable rural development by fostering cultural heritage, enhancing community cohesion, promoting mental and physical well-being and preserving traditional practices.
Recognizing and valuing these cultural services is crucial for policymaking and resource management aimed at supporting resilient communities. However, most studies have focused on provisioning and supporting services of SPSs (Hernández-Morcillo et al., Reference Hernández-Morcillo, Plieninger and Bieling2013; Cheng et al., Reference Cheng, Van Damme, Li and Uyttenhove2019). Cultural ecosystem services are often neglected because they are viewed as ‘intangible’, ‘non-material’ and ‘invisible’ in contrast to more tangible services (Cheng et al., Reference Cheng, Van Damme, Li and Uyttenhove2019). This lack of attention undermines their importance and the potential benefits they provide to communities and ecosystems. Thus, future research should focus on quantifying these services and exploring the mechanisms through which SPSs contribute to cultural sustainability.
Cultural heritage, identity and spiritual services
SPSs often reflect long-standing agricultural traditions, preserving community cultural heritage (Moreno et al., Reference Moreno, Franca, Correia and Godinho2014). The integration of trees and livestock management embodies local knowledge and practices, maintaining a sense of identity and continuity. Specific trees may hold symbolic meanings and be integral to local rituals and ceremonies, reinforcing social cohesion.
The ecological diversity supported by SPSs not only benefits agricultural productivity but also enriches the cultural landscape (Nair, Reference Nair1993; Daniel et al., Reference Daniel, Muhar, Arnberger, Aznar, Boyd, Chan, Costanza, Elmqvist, Flint, Gobster, Grêt-Regamey, Lave, Muhar, Penker, Ribe, Schauppenlehner, Sikor, Soloviy, Spierenburg, Taczanowska, Tam and Von Der Dunk2012; Isaac et al., Reference Isaac, Sinclair, Laroche, Olivier and Thapa2024). The presence of diverse flora and fauna is often integral to local culture, providing resources for traditional crafts, medicine and culinary practices. This biodiversity sustains cultural identity and heritage.
In addition, SPSs offer significant spiritual services that enhance community well-being and cultural identity. These services are rooted in the connections people establish with the landscape and the natural environment. The integration of livestock and trees can play a role in traditional rituals, linking agricultural cycles with spiritual beliefs and practices, thereby reinforcing cultural heritage.
Aesthetic and recreational value
The diverse landscapes created by SPSs provide aesthetic value, enhancing the quality of life for local populations (Chan et al., Reference Chan, Goldstein, Satterfield, Hannahs, Kikiloi, Naidoo, Vadeboncoeur, Woodside, Kareiva, Tallis, Ricketts, Daily and Polasky2011). These integrated systems contribute to the scenic beauty of rural areas, attracting visitors and fostering pride among residents. Furthermore, they offer recreational opportunities such as walking, hunting, bird observation, bike rides, horsemanship, painting and nature-based tourism, promoting mental well-being and community engagement (Acácio and Holmgren, Reference Acácio and Holmgren2014).
Social cohesion and community engagement
The collaborative nature of managing SPSs fosters social interactions and strengthens community bonds (Ali et al., Reference Ali, Kyatappanavar, Kabeer and Kale2024). Decision-making processes often involve local stakeholders, fostering a sense of ownership and responsibility towards shared resources. Knowledge-sharing regarding traditional practices and sustainable management further enhances social ties, creating a network of mutual support (Garrity et al., Reference Garrity, Akinnifesi, Ajayi, Weldesemayat, Mowo, Kalinganire, Larwanou and Bayala2010; Isaac et al., Reference Isaac, Sinclair, Laroche, Olivier and Thapa2024).
Educational opportunities
SPSs also serve as valuable educational platforms, offering opportunities to learn about sustainable agriculture and ecological stewardship. They provide a context for transmitting traditional ecological knowledge to younger generations, promoting environmental awareness and responsible resource management (Berkes, Reference Berkes2017). Incorporating these systems into educational programmes can highlight their importance in biodiversity conservation.
Mental and physical well-being
Access to nature and green spaces is associated with improved mental health and well-being (Kaplan and Kaplan, Reference Kaplan and Kaplan1989). SPSs, by providing tranquil environments, can reduce stress and promote relaxation. The connection to nature fosters a sense of place and belonging, which is crucial for psychological health (Barton and Pretty, Reference Barton and Pretty2010). Additionally, the availability of diverse natural resources enhances nutrition and physical health, aligning with cultural dietary practices (Garrity et al., Reference Garrity, Akinnifesi, Ajayi, Weldesemayat, Mowo, Kalinganire, Larwanou and Bayala2010).
Final considerations
Integrating different components in SPSs provides a wide range of ecosystem services due to the more efficient use of natural resources and interactions between agricultural elements. Overall, SPSs have demonstrated worldwide potential to enhance human food and forage production, nutrient cycling, soil fertility, BNF, carbon sequestration, GHG mitigation, erosion control, microclimate regulation, pollination, control of pests and diseases, biodiversity, residue absorption, water quality, spiritual enrichment, recreation and aesthetic experiences. However, most ecosystem services provided by SPSs depend on climate conditions, soil type, choice of species, system design and management practices to ensure that such benefits are not provided at the cost of other important services. The interaction among components is the major challenge for system management and ecosystem services evaluations. Long-term studies exploring the complex interactions within SPSs are needed to assess their impacts on ecosystem services fully.
Most of the research on SPSs is geographically concentrated in certain specific regions, especially in South and Central America, despite the recognized importance of such systems in Europe, Australia, New Zealand, sub-Saharan Africa (Parklands), India and Asia (‘cut-and-carry’ systems). This highlights the need for comprehensive studies considering a variety of geographic contexts to capture the nuances of interactions between SPS components under different designs and environments.
Conclusion
Adopting SPSs is a valuable strategy to promote more sustainable and diversified agriculture while providing valuable ecosystem services. Public policies, including evaluating ecosystem services, should be developed to promote the adoption of SPSs.
Acknowledgement
The authors acknowledge the National Council for Scientific and Technological Development (CNPq, Brazil).
Author contributions
C. B. M. Carvalho: writing – original draft. A. C. L. de Mello: conception, writing-review and editing. M. V. Cunha, V. X. O. Apolinário, J. C. B. Dubeux Jr., V. J. Silva, A. S. Medeiros, J. L. P. S. Izidro and I. L. Bretas: writing – review and editing.
Funding statement
No funding was received to assist with the preparation of this manuscript.
Competing interests
None.




 Open access
Open access