



Management Implications
Ventenata dubia (ventenata) is a nonnative, annual grass that has spread rapidly across the inland Pacific Northwest and eastward to the Northern Great Plains, potentially causing substantial ecological and economic impacts in perennial grasslands. Ventenata dubia’s accessibility and palatability as forage for livestock and wildlife is questionable, and controlling it with a herbicide may lead to improved forage quality and quantity. Indaziflam (Rejuvra®, formerly Esplanade®) is a preemergence herbicide that has shown great potential for annual grass control; however, its effect on V. dubia is less known than for the annual grass Bromus tectorum (downy brome). Furthermore, the herbicide’s effect on V. dubia and other aspects of vegetation beyond 3 yr is lacking. We applied indaziflam, other annual grass herbicides (glyphosate, imazapic, propoxycarbazone-sodium, rimsulfuron), indaziflam combined with each of those herbicides, and a non-sprayed control to an improved pasture in southwestern Montana in fall 2016. For 7 yr, we sampled canopy cover of V. dubia and the rest of the plant community. In the fifth year, we sampled the seedbank. Our results indicated that V. dubia cover was zero or close to zero in treatments that included indaziflam, and this trend held for 6 yr. However, by the seventh year, V. dubia had decreased to very low cover across the pasture, including the non-sprayed control, suggesting factors other than herbicide application were playing a role in vegetative dynamics. Perennial grasses and forbs were generally unaffected by treatment and did not increase in cover, even though V. dubia decreased. Annual forb cover and species richness were lowest in indaziflam treatments at 2 yr after treatment, but were similar in all treatments by 7 yr. Five years after herbicide application, the number of total seedlings (dicot and monocot combined) emerging from the seedbank was 12 to 33 times lower in indaziflam-treated soil than soil treated with other herbicides or not treated at all. Our study suggests that a single application of indaziflam along with abiotic and biotic environmental variables provided long-term (i.e., 7 yr) control of V. dubia and appeared to have depleted the seedbank across all species, but did not result in a release in desired vegetation as has been seen in some studies.
Introduction
Non-native annual grasses, for example, downy brome (Bromus tectorum L.) and ventenata [Ventenata dubia (Leers) Coss.], are associated with decreases in livestock and wildlife forage (Hart and Mealor Reference Hart and Mealor2021) and native species diversity (Jones et al. Reference Jones, Davis and Prather2020) on rangeland in the western United States. Herbicides can be used to minimize impacts from annual grass invasions and promote desired vegetation (Monaco et al. Reference Monaco, Mangold, Mealor, Mealor and Brown2017). Several herbicides labeled for rangeland use, including the active ingredients glyphosate, imazapic, rimsulfuron, and sulfosulfuron, provide control of annual grasses for 1 and sometimes 2 yr with minor injury to resident, desirable plants (Hirsch et al. Reference Hirsch, Monaco, Call and Ransom2012; Kyser et al. Reference Kyser, Ditomaso, Doran, Orloff, Wilson, Lancaster, Lile and Porath2007; Mangold et al. Reference Mangold, Parkinson, Duncan, Rice, Davis and Menalled2013; Sebastian et al. Reference Sebastian, Sebastian, Nissen and Beck2016b, Reference Sebastian, Nissen, Sebastian, Meiman and Beck2017c; Wallace and Prather Reference Wallace and Prather2016). Indaziflam, a preemergence cellulose biosynthesis–inhibiting herbicide, provides 2 to 3 yr (Clark et al. Reference Clark, Sebastian, Nissen and Sebastian2019, Reference Clark, Sebastian, Nissen and Sebastian2020; Koby et al. Reference Koby, Prather, Quicke, Beuschlein and Burke2019; Sebastian et al. Reference Sebastian, Sebastian, Nissen and Beck2016b) and possibly more years (Courkamp et al. Reference Courkamp, Meiman and Nissen2022a) of annual grass control. This long duration of control and potential depletion of the seedbank (Courkamp et al. Reference Courkamp, Meiman and Paschke2022b; Meyer-Morey et al. Reference Meyer-Morey, Lavin, Mangold, Zabinski and Rew2021) due to indaziflam’s persistence, may improve our ability to mitigate impacts of annual grasses (Sebastian et al. Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a).
Ventenata dubia was first reported in the 1950s in Washington and Idaho (Scheinost et al. Reference Scheinost, Stannard and Prather2008). Since then, it has invaded the intermountain Pacific Northwest and has spread eastward to the Northern Great Plains (Garner and Lakes Reference Garner and Lakes2019; Hart and Mealor Reference Hart and Mealor2021). It is associated with up to 50% yield loss in croplands across eastern Idaho and Washington (Wallace and Prather Reference Wallace and Prather2016) and substantial ecological and economic impacts in perennial grass habitats across the inland Pacific Northwest (Wallace et al. Reference Wallace, Pavek and Prather2015). Its accessibility and palatability as a source of forage for livestock and wildlife is questionable (Brummer et al. Reference Brummer, Morris and Laarman2023), and its removal with herbicide has been shown to improve forage quality and quantity (Hart and Mealor Reference Hart and Mealor2021). The shallow root system creates the potential for soil erosion in areas of monotypic invasions, thus decreasing land productivity and value (Scheinost et al. Reference Scheinost, Stannard and Prather2008). Ventenata dubia is known to invade areas previously inhabited by B. tectorum and medusahead [Taeniatherum caput-medusae (L.) Nevski] (Jones et al. Reference Jones, Norton and Prather2018; Wallace et al. Reference Wallace, Pavek and Prather2015; Wallace and Prather Reference Wallace and Prather2016). Seed longevity is estimated to be less than 4 yr, based upon a field study in which seeds of V. dubia were buried 2-cm deep and retrieved at various intervals; a small fraction of seeds was germinable at 3 yr but none were germinable at 4 yr (Wallace et al. Reference Wallace, Pavek and Prather2015).
Knowledge about the effectiveness of indaziflam on V. dubia and any impacts on desirable vegetation over an extended period is limited. Indaziflam and other commonly used active ingredients have been tested on V. dubia and associated vegetation in the intermountain Pacific Northwest over a 2-yr period (Koby et al. Reference Koby, Prather, Quicke, Beuschlein and Burke2019). Additionally, the effect of indaziflam on V. dubia and other vegetation, especially perennial grasses, has been evaluated across multiple sites in Wyoming for 1 yr posttreatment (Hart and Mealor Reference Hart and Mealor2021). The objective of our study was to compare effectiveness of indaziflam and other commonly used annual grass herbicides for V. dubia control, while accounting for effects on other vegetation over a 7-yr period at a site in southwestern Montana. We also assessed the effect of herbicide treatments on the seedbank at 5 yr after application. Response of V. dubia and desired vegetation by functional group, species richness, rank abundance, and number of seedlings (total, dicot, and monocot) emerging from the seedbank was assessed across 10 herbicide treatments: a non-sprayed control, indaziflam (Esplanade®; Anonymous 2016), glyphosate (Accord® XRT II; Anonymous 2014), imazapic (Plateau®; Anonymous 2011), propoxycarbazone-sodium (Lambient™; Anonymous 2009), and rimsulfuron (Matrix®SG; Anonymous 2017), and indaziflam combined with each of the other four active ingredients. Based on previous studies (Sebastian et al. Reference Sebastian, Nissen, De and Rodrigues2016a, Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a), we hypothesized that indaziflam applied alone and in combination with the other herbicides would control V. dubia for three or more years, while the other herbicides applied alone would control V. dubia for a year, and that a similar effect would be observed in the seedbank (i.e., emergence of fewer seedlings). Further, we sought to quantify any changes in species richness or abundance (i.e., cover) of vegetation other than V. dubia (perennial grass, perennial forb, and annual forb), expecting an increase in non-target vegetation as has been seen with other indaziflam studies (Clark et al. Reference Clark, Sebastian, Nissen and Sebastian2020; Hart and Mealor Reference Hart and Mealor2021; Koby et al. Reference Koby, Prather, Quicke, Beuschlein and Burke2019; Sebastian et al. Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a).
Materials and Methods
The study was carried out in an improved pasture near Bozeman in southwestern Montana (45.8808°N, 111.0567111°W). Mean annual precipitation is 47 cm and temperature is 6 C. Soil (0 to 15 cm) is a Sawicki cobbly loam (41% sand, 38% silt, 21% clay) with 6.2 pH and 2.7% organic matter. The site was dominated by smooth brome (Bromus inermis Leyss.) interspersed with western wheatgrass [Pascopyrum smithii (Rydb.) Á. Löve] and mountain brome (Bromus marginatus Nees ex Steud.), and a uniform infestation of V. dubia. The pasture is lightly grazed by sheep and cattle; however, grazing was excluded from the study area while the study was conducted by using an electric fence.
Ten herbicide treatments (Table 1) were replicated three times and arranged in a randomized complete block design using 9.1 m by 3.1 m plots. Herbicide treatments were applied on November 4, 2016, using a CO2 backpack sprayer that delivered 182 L water ha−1 at 290 kPa. The non-ionic surfactant Induce® (Helena Chemical Company, Billings, MT) was applied at a 0.25% v/v rate along with each herbicide treatment. At the time of treatment, V. dubia was in the 1- to 2-leaf growth stage and appeared alive and free of any frost injury.
Table 1. Herbicide treatments applied to Ventenata dubia–infested study site near Bozeman in southwestern Montana in November 2016.

Foliar canopy cover (hereafter referred to as cover) for each species was measured using three randomly located 20 cm by 50 cm Daubenmire (Reference Daubenmire1959) frames per plot in early to mid-July from 2017 through 2023. Cover (%) of all observed species was estimated to the nearest 1%, trace values were replaced with 0.1 for analyses, and total cover could exceed 100. Species other than V. dubia were later combined into functional groups of perennial grasses, perennial forbs, or annual forbs for analyses.
Five years posttreatment (August 2021), the germinable seedbank was sampled by compositing five cores (5.2 cm [w] by 5.6 cm [d] by 10.9 cm [l], 40 cm3 total volume) within each plot. The composited samples were stored for 3 mo at 5 C. Samples were sieved (4-mm and 2-mm sieves) to remove all litter, rocks, and root material, and any visible seeds were removed with tweezers but immediately returned to their respective samples. Each sample was spread over pasteurized (70 C for 60 min) potting soil in a 12 cm (w) by 5.7 (d) by 16.5 cm (l) tray within Montana State University’s Plant Growth Center (MSU PGC) (45.6681°N, 111.0533°W). The PGC potting soil is equal parts (by volume) of loam soil, washed concrete sand, and Canadian sphagnum peat moss with AquaGro 2000 G (Aquatrols, Paulsboro, NJ) wetting agent blended in at 454 g 0.76 cm−3 of soil mix. Samples were kept moist by misting with water at least once daily, and trays were monitored twice weekly for seedling emergence from November 3 to 1 December 1, 2021. Seedlings were counted and identified as monocot or dicot.
All analyses and graphical interpretations were conducted using R software v. 4.3.2 (R Core Team 2023). Specifically, we used routines within BiodiversityR, dplyr, emmeans, lme4, lmertest, multcomp, ggpatterns, and ggplot2 packages (Bates et al. Reference Bates, Mächler, Bolker and Walker2015; Mike et al. Reference Mike and Davis2022; Hothorn et al. Reference Hothorn, Bretz and Westfall2008; Kindt and Coe Reference Kindt and Coe2005; Kuznetsova et al. Reference Kuznetsova, Brockhoff and Christensen2017; Wickham Reference Wickham2016; Wickham et al. Reference Wickham, Romain, Henry and Müller2018). Ten treatments replicated three times and sampled over 7 yr precluded us from analyzing all treatments over all years due to a lack of statistical power. Therefore, we analyzed all treatments over 4 yr (2017 to 2019, 2023). Years were selected based upon expected persistence of the tested herbicides ranging from 1 to 3 yr and to capitalize on the relatively long-term nature of our data set at 7 yr.
Ventenata dubia, perennial grass, perennial forb, and annual forb cover over 4 yr were evaluated using separate linear mixed-effect models with the interaction between herbicide treatment and year as a fixed effect and sampling plot as a random effect. The interaction was significant and retained in all models except the perennial forb model, where herbicide treatment and year were retained as additive. Due to a violation of the assumption of heteroscedasticity, a square-root transformation was needed on all response variables. A type II ANOVA was used to determine whether each parameter indicated significant impact on the response variable (α = 0.05). A post hoc Tukey Kramer HSD pairwise comparison was used to elucidate differences between the herbicide treatment and year combinations using the glht function of the multcomp package (Hothorn et al. Reference Hothorn, Bretz and Westfall2008). A similar method was repeated for species richness; however, this model used a Poisson distribution and did not require transformation. Ventenata dubia was not included in species richness calculations. Bromus tectorum and houndstongue (Cynoglossum offinale L.) were the only other two invasive species found at the site, and they occurred infrequently (Supplementary Table S1).
Current stewardship guidelines indicate that indaziflam should be applied before germination of the target invasive annual grass, and if that window is missed, indaziflam should be combined with a postemergence herbicide, like imazapic (Envu Environmental Science US Inc. 2023). Therefore, we also analyzed V. dubia cover as affected by indaziflam and imazapic + indaziflam treatments and compared it to the non-sprayed control for all 7 yr of the study (2017 to 2023) using the same methods as described earlier. No other treatments were analyzed for all 7 yr of the study.
Rank and relative abundance of all species were also evaluated for the non-sprayed control, indaziflam, and imazapic + indaziflam treatments over 4 yr (2017 to 2019, 2023). For the top five species, abundance and proportion were calculated. Abundance represents the sum of cover estimates for a species across all replications of the treatment, and proportion relativizes the species to all other species found in the treatment.
For the seedbank analysis, a type II ANOVA was used to determine whether herbicide treatment affected total, dicot, and monocot seedlings that emerged (α = 0.05). Post hoc Tukey Kramer HSD pairwise comparisons were made to separate means.
Results and Discussion
Ventenata dubia Cover
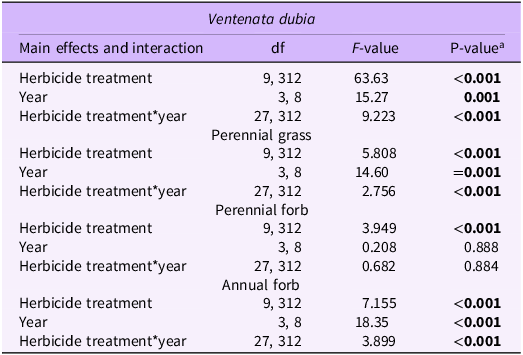
Differences in V. dubia cover were evident across treatments for the first 3 yr (2017 to 2019) but not the final year (2023) (P < 0.001) (Table 2; Figure 1). In 2017, all treatments, except glyphosate alone, had less V. dubia than the control. Indaziflam, imazapic, and propoxycarbazone-sodium applied alone reduced cover to a lesser extent than rimsulfuron alone or indaziflam combined with the other active ingredients. Cover of V. dubia in the indaziflam, imazapic, propoxycarbazone-sodium, and rimsulfuron treatments was 5.6 ± 2.0%, 8.0 ± 2.4%, 16.4 ± 3.4%, and 0.1 ± 0.2%, respectively, with the indaziflam, imazapic, and rimsulfuron treatments reducing V. dubia by at least 90% compared with the non-sprayed control, arguably a satisfactory outcome from a management perspective. Imazapic and rimsulfuron applied alone continued to reduce V. dubia cover in 2018 but were less effective by 2019. Treatments that included indaziflam provided the highest and most persistent reduction in V. dubia cover in 2018 and 2019, with V. dubia being absent or nearly absent from plots treated with either indaziflam alone or in combination with the other active ingredients. Our results are consistent with other studies that evaluated similar herbicides for V. dubia control (Davies and Hamerlynck Reference Davies and Hamerlynck2019; Koby et al. Reference Koby, Prather, Quicke, Beuschlein and Burke2019; Sebastian et al. Reference Sebastian, Nissen, De and Rodrigues2016a; Wallace and Prather Reference Wallace and Prather2016) and suggest multiple options for controlling V. dubia.
Table 2. Herbicide treatment effects on Ventenata dubia, perennial grass, perennial forb, and annual forb cover across 4 yr (2017–2019, 2023) for a site near Bozeman in southwestern Montana, using ANOVA.
a Bold numbers are significant at α = 0.05.

Figure 1. Ventenata dubia canopy cover (%) for 10 herbicide treatments (C, non-sprayed control; In, indaziflam; Gl, glyphosate; Gl+In, glyphosate + indaziflam; Im, imazapic; Im+In, imazapic + indaziflam; Pr, propoxycarbazone-sodium; Pr+In, propoxycarbazone-sodium + indaziflam; Ri, rimsulfuron; Ri+In, rimsulfuron + indaziflam) over 4 yr (2017–2019, 2023) applied in 2016 at a site near Bozeman in southwestern Montana. Boxes represent interquartile range, with line at center indicating median; whiskers are 95% range of the data; semi-transparent points are raw data points, with solid points indicating extreme outliers. Red asterisks (*) indicate treatments that were significantly different (P < 0.05) from that year’s non-sprayed control (C). Ventenata dubia cover ranged from 0% to 80%.
As stated earlier, current stewardship guidelines indicate that indaziflam should be mixed with a postemergence herbicide like imazapic if applied after annual grass emergence. When we analyzed the indaziflam and imazapic + indaziflam treatments compared with the non-sprayed control for each of 7 yr (2017 to 2023), we saw that these two treatments reduced V. dubia cover for 6 yr (through 2022), χ2 (12, N = 216) = 256.2, P < 0.001) (Figure 2). The two treatments performed similarly to each other, except in 2017, when V. dubia cover in the imazapic + indaziflam treatment was about 4% lower than in the indaziflam treatment (P < 0.01). The 2017 result was not surprising, as the herbicide treatments were applied postemergence in 2016, and we expected the combination treatment to outperform the indaziflam-only treatment, as the combination would have had activity on both emerged and yet-to-emerge seedlings. More surprising is that the indaziflam-only application reduced V. dubia compared with the non-sprayed control to the extent that it did, and that these treatments influenced V. dubia cover for 6 yr. Our study was longer than others (Davies and Hamerlynck Reference Davies and Hamerlynck2019; Koby et al. Reference Koby, Prather, Quicke, Beuschlein and Burke2019; Sebastian et al. Reference Sebastian, Nissen, De and Rodrigues2016a; Wallace and Prather Reference Wallace and Prather2016) and offers encouraging evidence for invasive plant managers regarding how long a single application of indaziflam can control V. dubia. Where rangeland and pastures are highly degraded and revegetation is necessary, our study also offers encouraging evidence that less-persistent herbicides like imazapic and rimsulfuron can provide satisfactory reductions in V. dubia for a year or two. Invasive plant managers could integrate these herbicides with seeding, allow seeded species a year or two to establish, then follow with an application of indaziflam to control any reinvading V. dubia if necessary.

Figure 2. Ventenata dubia canopy cover (%) for three herbicide treatments (C, non-sprayed control; In, indaziflam; Im+In, imazapic + indaziflam) over 7 yr (2017–2023) applied in 2016 at a site near Bozeman in southwestern Montana. Boxes represent interquartile range, with line at center indicating median; whiskers are 95% range of the data; semi-transparent points are raw data points, with solid points indicating extreme outliers. Red asterisks (*) indicate treatments that were significantly different (P < 0.05) from that year’s non-sprayed control.
By 2023, V. dubia cover was similarly low across all treatments, including the non-sprayed control (Figures 1 and 2); cover in the non-sprayed control decreased from 61% in 2017 to 0.7% in 2023, and there was a decrease in V. dubia each year, with levels <10% for the fifth year on. Interannual variability in annual grass abundance, including V. dubia, is common (Courkamp et al. Reference Courkamp, Meiman and Nissen2022a; Orloff et al. Reference Orloff, Mangold and Menalled2015; Wallace and Prather Reference Wallace and Prather2016) and can be strongly influenced by precipitation and temperature (Adhikari et al. Reference Adhikari, Mangold, Mainali and Rew2023; Bradley et al. Reference Bradley, Curtis, Fusco, Abatzoglou, Balch, Dadashi and Tuanmu2018; Bradley and Mustard Reference Bradley and Mustard2005; Clinton et al. Reference Clinton, Potter, Crabtree, Genovese, Gross and Gong2010). Spring through midsummer (April 1 to July 31) precipitation varied greatly during the study from a low of 12 cm in 2021 to a high of 23 cm in 2018 (NOAA 2023). Years 4 (2020), 5 (2021), and 6 (2022) were generally drier than the first 3 yr or the last year of the study. Year 5 (2021) was very dry, and the entire county where the study site was located (Gallatin County) was in severe to extreme drought by the end of July (NDMC 2023). However, V. dubia cover remained low despite the final summer (2023) being wet (22 cm).
Cover of Other Functional Groups
Perennial grass cover was influenced by the interaction of herbicide treatment and year (P < 0.001; Table 2) with the effect of glyphosate in 2017 being primarily responsible for this interaction (Figure 3). Cover in the glyphosate treatment was 5.2 ± 1.8%, while mean cover in the other treatments, including the non-sprayed control, ranged from 12.6% to 33.8%. For years other than 2017, there were no differences in perennial grass cover relative to the non-sprayed control and among treatments, and cover remained mostly consistent within each treatment over the duration of the study. Two exceptions to this were the glyphosate treatment, which increased in cover from 2017 to 2023 (P < 0.01), and the imazapic + indaziflam treatment, which declined in perennial grasses from 2017 to 2018 (P < 0.01) and then increased from 2018 to 2023 (P < 0.01).

Figure 3. Perennial grass canopy cover (%) for 10 herbicide treatments (C, non-sprayed control; In, indaziflam; Gl, glyphosate; Gl+In, glyphosate + indaziflam; Im, imazapic; Im+In, imazapic + indaziflam; Pr, propoxycarbazone-sodium; Pr+In, propoxycarbazone-sodium + indaziflam; Ri, rimsulfuron; Ri+In, rimsulfuron + indaziflam) over 4 yr (2017–2019, 2023) applied in 2016 at a site near Bozeman in southwestern Montana. Boxes represent interquartile range, with line at center indicating median; whiskers are 95% range of the data; semi-transparent points are raw data points, with solid points indicating extreme outliers. Red asterisks (*) indicate treatments that were significantly different (P < 0.05) from that year’s non-sprayed control.
Some research has shown an increase in perennial grass abundance following herbicide applications to control V. dubia or other invasive annual grasses (Clark et al. Reference Clark, Sebastian, Nissen and Sebastian2020; Courkamp et al. Reference Courkamp, Meiman and Nissen2022a; Hart and Mealor Reference Hart and Mealor2021; Koby et al. Reference Koby, Prather, Quicke, Beuschlein and Burke2019; Sebastian et al. Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a), and on range and pasture where livestock production is the primary management objective, this increase is touted as an economic benefit of controlling invasive annual grasses that can help to offset the cost of applying herbicide (Hart et al. Reference Hart, Ritten and Mealor2023). We did not see an increase in perennial grass cover as we had hypothesized, which was disappointing, considering grazing was deferred for the duration of the study. We did not measure perennial grass biomass over the 7-yr study, and it is possible that meaningful increases in biomass were not reflected in cover; however, biomass was not higher at 2 yr posttreatment (Harvey Reference Harvey2019). The dominant perennial grass species at our site were B. inermis, B. marginatus, and P. smithii, all species considered competitive, resilient, and stress tolerant (USDA-NRCS 2023a, 2023b, 2023c). Ventenata dubia appeared to grow under the canopy of these species, especially the Bromus species, and our results suggest that V. dubia was not affecting the abundance of these species, as its removal did not coincide with an increase in perennial grass.
Perennial forb cover was affected by herbicide treatment (P < 0.001), but not year (P = 0.888) or the interaction of herbicide treatment and year (P = 0.884; Table 2). Mean cover ranged from 0.3% to 4.1% across herbicide treatments, and only propoxycarbazone-sodium differed from the non-sprayed control (Figure 4). Comparing across treatments other than the non-sprayed control, perennial forb cover was higher in the glyphosate treatment (4.1 ± 1.1%) than in the indaziflam (0.9 ± 0.5%), imazapic (0.6 ± 0.4%), propoxycarbazone-sodium (0.3 ± 0.3%), and rimsulfuron + indaziflam (0.8 ± 0.5%) treatments; and it was higher in the glyphosate + indaziflam treatment (3.4 ± 1.0%) than in the propoxycarbazone-sodium treatment.

Figure 4. Perennial forb canopy cover (%) for 10 herbicide treatments (C, non-sprayed control; In, indaziflam; Gl, glyphosate; Gl+In, glyphosate + indaziflam; Im, imazapic; Im+In, imazapic + indaziflam; Pr, propoxycarbazone-sodium; Pr+In, propoxycarbazone-sodium + indaziflam; Ri, rimsulfuron; Ri+In, rimsulfuron + indaziflam) combined over 4 yr (2017–2019, 2023) applied in 2016 at a site near Bozeman in southwestern Montana. Boxes represent interquartile range, with line at center indicating median; whiskers are 95% range of the data; semi-transparent points are raw data points, with solid points indicating extreme outliers. There were no significant differences (P < 0.05) relative to the control treatment; however, there were differences among treatments.
Annual forb cover was affected by the interaction between herbicide treatment and year (P < 0.001; Table 2), but there was no evidence of a difference in cover between a herbicide treatment and the non-sprayed control within a year (Table 2; Figure 5). Instead, the interaction was due to differences among herbicide treatments (other than the non-sprayed control) in the second year postapplication (2018), when cover levels tended to be higher in the treatments without indaziflam. In 2018, which was 2 yr after herbicide application, imazapic (5.1 ± 1.2%), propoxycarbazone (5.4 ± 1.3%), and rimsulfuron (5.0 ± 1.2%) applied alone had higher annual forb cover than glyphosate (1.8 ± 0.7%) applied alone or indaziflam and any of the active ingredients combined with indaziflam (0.0 to 1.3%). By 2023, annual forbs were rarely found at the study site regardless of herbicide treatment.

Figure 5. Annual forb canopy cover (%) for 10 herbicide treatments (C, non-sprayed control; In, indaziflam; Gl, glyphosate; Gl+In, glyphosate + indaziflam; Im, imazapic; Im+In, imazapic + indaziflam; Pr, propoxycarbazone-sodium; Pr+In, propoxycarbazone-sodium + indaziflam; Ri, rimsulfuron; Ri+In, rimsulfuron + indaziflam) combined over 4 yr (2017–2019, 2023) applied in 2016 at a site near Bozeman in southwestern Montana. Boxes represent interquartile range, with line at center indicating median; whiskers are 95% range of the data; semi-transparent points are raw data points, with solid points indicating extreme outliers. There were no significant differences (P < 0.05) relative to the non-sprayed control.
Perennial and annual forbs may respond differently to an indaziflam application. Based on previous research, perennial forbs can increase (Sebastian et al. Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a), decrease (Meyer-Morey et al. Reference Meyer-Morey, Lavin, Mangold, Zabinski and Rew2021), or be unaffected (Clark et al. Reference Clark, Sebastian, Nissen and Sebastian2020; Sebastian et al. Reference Sebastian, Sebastian, Nissen and Beck2016b), while annual forbs appear to decrease (Courkamp et al. Reference Courkamp, Meiman and Paschke2022b; Meyer-Morey et al. Reference Meyer-Morey, Lavin, Mangold, Zabinski and Rew2021), which makes sense, given the life histories of the two functional groups and the characteristics of indaziflam as a preemergence herbicide. In our study, there was no response of perennial forbs, but annual forbs during the second year after application decreased when indaziflam was applied alone or in combination with other active ingredients compared with when those active ingredients were applied singly.
Species Richness
Over the course of the study, 51 species were recorded at the site (not including V. dubia) (Supplementary Table S1). Species richness was affected by the interaction of herbicide treatment and year, χ2 (27, N = 360) 41.01, P = 0.041), which was due largely to the second year after herbicide application (2018). In this year, there were differences between the non-sprayed control (6.5 ± 0.9 species) and any treatment that included indaziflam (2.4 to 3.7 species) (Figure 6). In that same year, all treatments that included indaziflam resulted in lower species richness than any treatment that was a singularly applied active ingredient. Species richness increased from 2017 to 2018 in all treatments that did not include indaziflam, which may have been due to higher precipitation in 2018, for example, June precipitation in that year was two times what it was in 2017 (8.3 cm vs. 4.0 cm) (NOAA 2023). By 2019, species richness declined in those treatments that peaked in 2018 so that all treatments had similar species richness, a result that held into 2023.

Figure 6. Species richness (species per 0.1 m2) for 10 herbicide treatments (C, non-sprayed control; In, indaziflam; Gl, glyphosate; Gl+In, glyphosate + indaziflam; Im, imazapic; Im+In, imazapic + indaziflam; Pr, propoxycarbazone-sodium; Pr+In, propoxycarbazone-sodium + indaziflam; Ri, rimsulfuron; Ri+In, rimsulfuron + indaziflam) over 4 yr (2017–2019, 2023) applied in 2016 at a site near Bozeman in southwestern Montana. Boxes represent interquartile range, with line at center indicating median; whiskers are 95% range of the data; semi-transparent points are raw data points, with solid points indicating extreme outliers. Red asterisks (*) indicate treatments that were significantly different (P < 0.05) from that year’s control treatment.
Previous research has indicated increases (Clark et al. Reference Clark, Sebastian, Nissen and Sebastian2019; Sebastian et al. Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a), decreases (Meyer-Morey et al. Reference Meyer-Morey, Lavin, Mangold, Zabinski and Rew2021), and no change (Hart and Mealor Reference Hart and Mealor2021) in species richness in response to applications of indaziflam for annual grass control. These studies and our own occurred in different geographic locations that ranged from sites with low disturbance and high richness and diversity (e.g., Meyer-Morey et al. Reference Meyer-Morey, Lavin, Mangold, Zabinski and Rew2021) to more disturbed and depauperate sites (e.g., Sebastian et al. Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a)—our current site was an improved pasture dominated by perennial grasses. We saw a short-term decrease in species richness, possibly due to loss of annual forbs, but there was no effect on species richness by the end of the study. Our results indicate indaziflam controlled V. dubia with minimal impacts to non-target vegetation. However, there is still limited knowledge of the effects of cellulose biosynthesis–inhibiting herbicides like indaziflam on monocot and dicot proteins (Brabham et al. Reference Brabham, Lei, Gu, Stork, Barrett and DeBolt2014; Sebastian et al. Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a). Given that there are thousands of species present on rangeland in the western United States (FNA 1993+), more investigation of non-target species’ tolerances and their long-term responses to indaziflam is needed.
Rank and Relative Abundance
With few exceptions, B. inermis, B. marginatus, and P. smithii were among the top five most abundant species in the non-sprayed control, indaziflam, and indaziflam + imazapic treatments over the duration of the study (Table 3). Thickspike wheatgrass [Elymus lanceolatus (Scribn. & J.G. Sm.) Gould], prairie Junegrass [Koeleria macrantha (Ledeb.) Schult], and intermediate wheatgrass [Thinopyrum intermedium (Host) Barkworth & D.R. Dewey] were three additional perennial grasses that appeared in the top five most abundant as the study progressed. Common sheep sorrel (Rumex acetosella L.) was the most consistently abundant forb, appearing in the non-sprayed control 2017 to 2019 and 2023; it did not appear in the indaziflam, imazapic, or indaziflam + imazapic treatments, except for the indaziflam treatment in 2023.
Table 3. Relative abundance of the top five species in the non-sprayed control (C), indaziflam (In), and imazapic + indaziflam (Im+In) treatments for the first 3 yr (2017–2019) and 7 yr (2023) after treatment application at a study site near Bozeman in southwestern Montana.

a Abundance represents the sum of cover estimates for a species across all replications of the treatment.
b Proportion relativizes the species to all other species found in the treatment.
Seedbank
In addition to herbicides affecting aboveground vegetation dynamics for up to 6 yr, herbicide treatments affected the total (all species) (P < 0.001), dicot (P < 0.001), and monocot (P = 0.047) seedlings emerging from the seedbank 5 yr after application (Table 4). For total seedlings, glyphosate alone had more seedlings than any other treatment at 165 ± 7.4 seedlings per sample (Table 4). Glyphosate was followed by propoxycarbazone-sodium, the non-sprayed control, rimsulfuron, and imazapic, with seedlings per sample ranging from 60 to 90. Indaziflam applied alone and in combination with the other herbicides resulted in five or fewer total seedlings per sample. Dicot and monocot seedlings trended similarly to total seedlings, but differences among treatments were not as distinct, and the effects of indaziflam were not as clearly evident as they were with total seedlings. For dicot and monocot seedlings separately, none of the treatments reduced seedlings compared with the non-sprayed control (Table 4).
Table 4. Number of total, dicot, and monocot seedlings (mean ± SE) of any species emerging from the seedbank (per 40-cm3 sample) 5 yr after herbicide treatments were applied at a site near Bozeman in southwestern Montana a .

a The same letter following means indicates no difference between treatments within total seedlings (α = 0.05), dicot seedlings (α = 0.05), and monocot seedlings (α = 0.10).
Seedbank depletion of target species is critical for invasive annual grass management (Sebastian et al. Reference Sebastian, Nissen, Sebastian and Beck2017b), and indaziflam remains active at low concentrations in the upper couple of centimeters of soil for at least 2 to 3 yr (Sebastian et al. Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a). Ventenata dubia seed longevity is estimated to be less than 4 yr (Wallace et al. Reference Wallace, Pavek and Prather2015), and our results provide evidence of seedbank depletion up to 5 yr after a single indaziflam application. At a site in Wyoming, a single application of indaziflam provided 5 yr of B. tectorum control (Courkamp et al. Reference Courkamp, Meiman and Nissen2022a). Courkamp et al. (Reference Courkamp, Meiman and Nissen2022a) speculated that the maximum single-use application rate of indaziflam (Indaziflam GRZ; USEPA 2023), the rate used in our study, could have depleted the seedbank of B. tectorum, which is estimated to be up to a year longer than that of V. dubia (Thill et al. Reference Thill, Beck and Callihan1984).
We did not identify seedlings to species in the seedbank analysis, so we are unable to say whether the indaziflam treatments were selecting for V. dubia over other species in the seedbank. However, we differentiated between monocots and dicots, and dicots generally made up a greater proportion of total seedlings emerging from the seedbank than monocots. Additionally, indaziflam treatments tended to result in much lower dicot or monocot seedling emergence when compared with the treatments that yielded the highest number of seedlings (e.g., glyphosate). Indaziflam is reported to have more activity on monocots than dicots (Sebastian et al. Reference Sebastian, Fleming, Patterson, Sebastian and Nissen2017a), contributing to its value for invasive annual grass management. At the same time, there is increasing evidence that indaziflam reduces native annual forbs (dicots) in the seedbank (Courkamp et al. Reference Courkamp, Meiman and Paschke2022b; Meyer-Morey et al. Reference Meyer-Morey, Lavin, Mangold, Zabinski and Rew2021). Whether the reduction of the annual forb seedbank is reflected in the aboveground plant community has been mixed. Meyer-Morey et al. (Reference Meyer-Morey, Lavin, Mangold, Zabinski and Rew2021) observed an absence of native annual forbs emerging from the seedbank ex situ and in situ from high elevation, diverse sagebrush steppe communities 2 yr following indaziflam application. In contrast, Courkamp et al. (Reference Courkamp, Meiman and Paschke2022b) observed native annual forbs at their field site 3 yr after an indaziflam application, even though those same species were absent from the seedbank.
Our study, which analyzed the seedbank 5 yr after indaziflam application, provides longer-term evidence relative to other studies (Courkamp et al. Reference Courkamp, Meiman and Paschke2022b; Meyer-Morey et al. Reference Meyer-Morey, Lavin, Mangold, Zabinski and Rew2021) of indaziflam’s depauperating effect on the seedbank. We saw parallels between the seedbank and aboveground vegetation in terms of annual species, which is consistent with other seedbank studies where annual species are typically well-represented using a greenhouse germination method (Ball and Miller Reference Ball and Miller1989; Chiquoine and Abella Reference Chiquoine and Abella2018). The methods used for our seedbank analysis were similar to those of Sebastian et al. (Reference Sebastian, Nissen, Sebastian and Beck2017b) but shorter in duration (about 1 mo) than most studies (e.g., 4 to 17 mo) (Ball and Miller Reference Ball and Miller1989; De Villiers et al. Reference De Villiers, Van Rooyen and Theron2003; Meyer-Morey et al. Reference Meyer-Morey, Lavin, Mangold, Zabinski and Rew2021; Plue et al. Reference Plue, Colas, Auffret and Cousins2017; Price et al. Reference Price, Wright, Gross and Whalley2010). Furthermore, our seedbank analysis did not employ methods such as stirring (Courkamp et al. Reference Courkamp, Meiman and Paschke2022b; Price et al. Reference Price, Wright, Gross and Whalley2010), stratification (Meyer-Morey et al. Reference Meyer-Morey, Lavin, Mangold, Zabinski and Rew2021; Plue et al. Reference Plue, Colas, Auffret and Cousins2017), or addition of gibberellic acid (Chiquoine and Abella Reference Chiquoine and Abella2018) to stimulate additional germination following the initial germination period and therefore may have underrepresented the total number of seeds. However, our seedbank analysis offers a cursory evaluation of the effects of indaziflam on the seedbank 5 yr after application; further and more rigorous examination of how impacts to the seedbank influence aboveground plant community composition over time is warranted.
We observed variability in V. dubia cover over time. Cover was highest in the non-sprayed control in 2017, and by the seventh year after treatments were applied, V. dubia was nearly absent from the study site, regardless of treatment. Interannual variability in annual grass abundance is common (Courkamp et al. Reference Courkamp, Meiman and Nissen2022a; Ehlert et al. Reference Ehlert, Engel and Mangold2015; Orloff et al. Reference Orloff, Mangold and Menalled2015; Wallace and Prather Reference Wallace and Prather2016) and can be strongly influenced by precipitation and temperature (Adhikari et al. Reference Adhikari, Mangold, Mainali and Rew2023; Bradley et al. Reference Bradley, Curtis, Fusco, Abatzoglou, Balch, Dadashi and Tuanmu2018; Bradley and Mustard Reference Bradley and Mustard2005; Clinton et al. Reference Clinton, Potter, Crabtree, Genovese, Gross and Gong2010). Spring through midsummer precipitation varied during our study with the middle years (2020 and 2021) being drier than average. These variations in precipitation appear to have been particularly detrimental to annual species at our site, as both V. dubia and annual forb cover declined to near zero, while perennial grasses and forbs maintained their presence. Perennial grasses like B. inermis and B. marginatus, generally regarded for their competitive vigor (USDA-NRCS 2023a, 2023b), became proportionately more dominant as the study progressed. Our results support the view that plant communities change in response to invasive annual grass management but, importantly, also in response to abiotic and biotic interactions, and monitoring beyond a year or two after management may reveal unexpected vegetation dynamics.
Given the decline in V. dubia at our improved pasture site regardless of herbicide treatment, a rangeland manager may question the need to invest in V. dubia management. Variability in precipitation coupled with competition dynamics among species within a plant community adds to the challenge of making management decisions regarding V. dubia. While there is still much to learn about V. dubia, we argue that its control is advised, given evidence for its recent (Jones et al. Reference Jones, Davis and Prather2020; Ridder et al. Reference Ridder, Morris, Day and Kerns2022) and predicted future (Adhikari et al. Reference Adhikari, Mangold, Mainali and Rew2023) success. For example, V. dubia cover increased by 30% and frequency by 50% in a 15-yr period on undisturbed (no fire, no grazing) grasslands in the inland Pacific Northwest (Ridder et al. Reference Ridder, Morris, Day and Kerns2022); furthermore, suitable habitat for V. dubia is predicted to increase under future climate scenarios in the same county where this study occurred, especially along roads and in agricultural lands (Adhikari et al. Reference Adhikari, Mangold, Mainali and Rew2023). Ventenata dubia’s lack of value as forage for livestock or wildlife (Brummer et al. Reference Brummer, Morris and Laarman2023) is yet another reason to control it. On sites like ours, with perennial grass cover approximately 20%, monitoring before management would be a viable option, with particular focus on the abundance of perennial species that can reproduce vegetatively to help ensure satisfactory aboveground cover. Our study occurred at a single site, which can limit its applicability to other areas where V. dubia is problematic.
Rather than confounding decision making, we hope our results are encouraging to rangeland managers in that a single application of indaziflam along with abiotic and biotic environmental variables, like variable precipitation over time, an exclusion of grazing, and competitive perennial grasses dominating the site, provided numerous years of V. dubia control and appear to have depleted the seedbank. Treatment did not result in an increase of any other desired species, though, and this potential outcome should be considered when making management decisions.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/inp.2024.20
Acknowledgments
We would like to thank landowners Sas and Stuart Weber for granting us space and patience and sharing their experiences with V. dubia as we conducted the study over the years.
Funding statement
We would like to thank Harry Quicke and Shannon Clark with Envu (formerly Bayer CropSciences) for funding that partially supported this project along with funding that supported students, including the Alfred P. Sloan Indigenous Graduate Partnership, Montana Noxious Weed Trust Fund, and Western Sustainable Agriculture Research and Education. LJR and JMM are supported by NIFA-USDA Hatch MONB000363 and MONB00359, respectively.
Competing interests
No conflicts of interest have been declared.













 Open access
Open access