



Social media summary: Worldwide, millions of women stop using contraceptives. Why? Novel hypotheses from evolutionary public health
1. Introduction
Uncovering the processes underpinning the dynamics of contraceptive behaviour is relevant for understanding the demographic transition, defined as the shift from high to low fertility with increasing levels of socioeconomic development in recent and contemporary history (Bongaarts & Watkins, Reference Bongaarts and Watkins1996; Borgerhoff Mulder, Reference Borgerhoff Mulder1998). Of course, the availability of modern contraception is neither necessary nor sufficient for fertility decline: the European demographic transition started around two centuries before the contraceptive pill was commercialised (Livi-Bacci, Reference Livi-Bacci, and S and Watkins1986) and despite modern contraception being accessible in many parts of sub-Saharan Africa, fertility remains high on average. Yet, as contraceptive technology has been introduced recently in some areas, it is possible to study the cultural evolution of contraceptive use, i.e. the change in the prevalence of contraceptive use at the population level over time. Note that by cultural evolution we mean the process of change over time rather than any specific theoretical framework aiming at explaining the evolutionary process (i.e. behavioural ecology or cultural evolution approaches). Given that contraceptive use is linked to reduced infant and maternal mortality, as well as improved socioeconomic opportunities (Ahmed et al., Reference Ahmed, Li, Liu and Tsui2012; Canning & Schultz, Reference Canning and Schultz2012; Cleland et al., Reference Cleland, Conde-Agudelo, Peterson, Ross and Tsui2012; United Nations, Department of Economic and Social Affairs, 2015), uncovering the mechanisms linking individual behaviour to population change in contraceptive use has important public health implications.
Thus far, evolutionary research on contraceptive behaviour has focused on why people adopt low-fertility ideals and begin using fertility regulation methods, with relatively little attention given to processes underpinning the maintenance of contraceptive behaviour past its initial adoption. Yet, reducing fertility is more than an idea to be accepted once or a process to be initiated. It is an ongoing practice with much variation in execution over time. For instance, contraceptive use can involve a mix of so-called ‘traditional’ and ‘modern’ contraceptive techniques including abstinence, withdrawal, calendar rhythm, condoms, injectables, IUDs, pills, emergency pills and herbal remedies (Marston et al., Reference Marston, Renedo, Nyaaba, Machiyama and Tapsoba2016). In turn, methods differ in their length of action (e.g. short-term vs. long-term), efficacy (e.g. withdrawal is less efficacious than IUD) and perceived cost for future fertility (e.g. non-hormonal vs. hormonal methods leading to side-effects). While family planning studies initially researched the determinants of contraceptive ever-use across methods, there is now a noticeable shift from a focus on ‘adopters’ to a concern over ‘discontinuers’, i.e. women who stop using a given method of contraception after initiating use (Jain, Reference Jain2014; Castle & Askew, Reference Castle and Askew2015). There are multiple reasons for discontinuation, and sometimes multiple reasons per discontinuation event, but discontinuation due to side-effects is the most important cause of discontinuation while in need (i.e. stopping despite the desire to continue limiting or spacing their fertility; Ali et al., Reference Ali, Cleland and Shah2012; Castle & Askew, Reference Castle and Askew2015). The possibility for individuals to both learn from their experience of contraceptive techniques and socially transmit knowledge gained from their positive or negative experiences might have important implications for both contraceptive and fertility dynamics. Further, women's decision to discontinue using contraception (because of side-effects or other reasons) is likely to be shaped by socio-ecological and individual factors (e.g. age, parity, marital and socio-economic status) that influence variation in life-history trade-offs between current fertility and other components of fitness. Thus far, however, the evolutionary human sciences have not yet engaged with the issue of contraceptive discontinuation at the individual level.
This paper discusses the strengths, limitations and future of studies researching the cultural evolution of contraception. While our discussion focuses primarily on modern methods, a similar reasoning could also be applied to traditional methods, even though traditional methods are less efficacious than modern ones for avoiding unplanned pregnancies. We start by briefly outlining evolutionary perspectives on fertility and contraceptive dynamics in high fertility or transitioning populations with widespread availability of family planning services, including behavioural ecology and cultural evolutionary approaches. Second, we suggest ways to harness evolutionary frameworks for understanding variation in the propensity to discontinue contraception due to side-effects (Table 1), the main reason for discontinuation whilst in need. Third, we illustrate how an integration of both adoption and discontinuation dynamics may provide a fuller understanding of the evolution of fertility-regulating behaviour, the transition from high to low fertility and the impact of family programmes on the demographic transition. We conclude by pointing to the importance of individual experience and the flexibility of practices for modelling cultural evolutionary processes.
Table 1. Evolutionary approaches to understanding patterns of discontinuation of modern contraception due to side-effects. We focus on discontinuation whilst in need, for which the most common reason is side-effects

C, Contraception; CU, contraceptive use; D, Discontinuation; DSE, discontinuation due to side-effects; DOR, discontinuation due to other reasons; SE, side-effects; S, switch between methods. We propose an evolutionary ecological approach, identifying the ultimate (i.e. evolutionary history and function utility) and proximate (i.e. mechanisms underlying behaviour) factors responsible for patterns of variation.
2. Current evolutionary perspectives on the use of modern contraception
Why do people start using modern contraception? Is fertility change driven by a change of ideas and/or a change of economies? Much work has been done in demography and family planning, where two models have been proposed to explain the adoption of fertility-regulating technologies: (a) demand theory (Easterlin, Reference Easterlin1975), whereby contraceptive adoption is driven by levels of socioeconomic development and thus responds to a previously unmet need (when a woman does not want to get pregnant but is not using contraception); and (b) diffusion theory (Casterline, Reference Casterline and Casterline2001), whereby a new idea, technology or behaviour spreads in the population independently of the level of socioeconomic development. A review of 63 developing countries found that, at the aggregate level, family planning programmes both decrease unmet need for contraception by removing ‘barriers’ and increase the demand for contraception through educating people about the benefits of small family size (Bongaarts, Reference Bongaarts2014), thus providing support for both theories. Yet standard quantitative population-level models of demographic change do not typically consider within-population variation in behaviour that affects fecundity and mortality, nor aspects of cultural transmission that can facilitate the spread of fertility-reducing behaviours (Creanza et al., Reference Creanza, Kolodny and Feldman2017), limiting the scope of their projections. It has thus been suggested that evolutionary approaches should be integrated into demography (Colleran, Reference Colleran2016), as they focus on what has changed in the environment and what evolved mechanisms underpin fertility and contraceptive dynamics. In this section, we briefly present current evolutionary frameworks, highlighting areas of debate for their integration, and review the empirical evidence for each framework for the adoption (emergence and spread) and the maintenance of modern contraception in a population.
(a) Toward an integrated evolutionary framework
To understand why a low-fertility variant emerges and spreads, evolutionary scholars have used principles from primarily two frameworks: behavioural ecology (Nettle et al., Reference Nettle, Gibson, Lawson and Sear2013) and cultural evolution (Mesoudi, Reference Mesoudi2016). Broadly, a behavioural ecology approach is concerned with the fitness consequences of a given behaviour for the individual and assume that mechanisms do not constrain adaptive responses to the ecology (Winterhalder & Smith, Reference Winterhalder and Smith2000). In contrast, a cultural evolution approach focuses on the social transmission of a cultural variant, often independently of its impact on individual fitness (but see El Mouden et al., Reference El Mouden, André, Morin and Nettle2014). Both approaches assume the dynamics of cultural change over ecological times to be constrained by evolved cognitive dispositions at the individual level, but while BE models are concerned with the impact of behaviour on individual fitness, CE models are concerned with the impact of social transmission biases on the fitness of the cultural trait (e.g. the prevalence of contraceptive uptake at the population level).
Behavioural ecology theory advances that individuals adjust their fertility as a response to an increased perceived trade-off between fertility and investment in their own and their offspring's embodied capital (Kaplan, Reference Kaplan1996; Borgerhoff Mulder, Reference Borgerhoff Mulder1998; Kaplan et al., Reference Kaplan, Lancaster, Tucker and Anderson2002). As the risk of death in early childhood decreases, the importance of parental investment for an individual's ability to outcompete their peers increases, especially in skill-based competitive labour markets (Kaplan, Reference Kaplan1996), leading to increased trade-offs between fertility and investment in their own and their offspring's capital and ultimately, fertility decline (Mace, Reference Mace2008). While empirical studies in low-fertility populations have not found lowered fertility rates to be optimal (Kaplan et al., Reference Kaplan, Lancaster, Johnson and Bock1995; Goodman et al., Reference Goodman, Koupil and Lawson2012), there is some empirical support for a link between variation in fertility trade-offs and inter-individual diversity in fertility (Shenk, Reference Shenk2009; Shenk et al., Reference Shenk, Towner, Kress and Alam2013) and age at marriage in India (Shenk, Reference Shenk2009) or the timing of adoption of modern contraception in Ethiopia (Alvergne et al., Reference Alvergne, Gibson, Gurmu and Mace2011; see full review in Colleran, Reference Colleran2016), suggesting that changing fertility trade-offs can promote fertility change. Yet it is also well known to demographers that socioeconomic circumstances alone cannot fully explain the dynamics of fertility behaviour (Cleland, Reference Cleland and Casterline2001), although the issue remains debated and economic concerns may be of high importance in certain contexts (Colleran & Snopkowski, Reference Colleran and Snopkowski2018).
Cultural evolution theorists have proposed that the worldwide phenomenon of fertility decline can be explained by evolved cognitive dispositions for social learning (Richerson & Boyd, Reference Richerson and Boyd2006). The leading theory posits that if people who delay reproduction become more educated and/or wealthier, and if people imitate the most successful and prestigious, then a fertility-reducing variant could spread (Richerson & Boyd, Reference Richerson and Boyd1984; Ihara & Feldman, Reference Ihara and Feldman2004). While direct causal evidence for the role of prestige-biased transmission in spreading fertility reducing behaviour is not yet available, studies at a later stage of the demographic transition found stronger effects for community-level and social-network variables than individual factors in predicting fertility (Colleran et al., Reference Colleran, Jasienska, Nenko, Galbarczyk and Mace2014) or contraceptive use (Colleran & Mace, Reference Colleran and Mace2015). Thus, a comprehensive evolutionary account of fertility decline should incorporate insights from a cultural evolution (CE) approach to traditional behavioural ecology (BE) models (Colleran, Reference Colleran2016).
There is some debate as to whether BE and CE models are compatible, limiting the scope for their integration. Cultural models can be seen both as mechanistic models of how fertility declines or as causal models for why fertility declines (Mattison & Shenk, Reference Mattison, Shenk, Burger, Lee and Sear2020), but there is significant debate as to whether CE offers a proximate (Scott-Phillips et al., Reference Scott-Phillips, Dickins and West2011) or an ultimate (Laland et al., Reference Laland, Sterelny, Odling-Smee, Hoppitt and Uller2011, Reference Laland, Uller, Feldman, Sterelny, Müller, Moczek and Strassmann2014; Bateson & Laland, Reference Bateson and Laland2013) explanation. To evaluate the conditions under which CE and BE models are compatible it is helpful to (a) distinguish between the impact of behaviour on individual fitness from its impact on the dynamics of a cultural trait, (b) separate out the phases of the dynamics of the cultural evolutionary process (emergence, spread and maintenance) (Alvergne et al., Reference Alvergne, Gibson, Gurmu and Mace2011; Colleran et al., Reference Colleran, Jasienska, Nenko, Galbarczyk and Mace2014) and (c) interact rather than add socioeconomic and cultural factors. First, while CE models are not concerned with the impact of behaviour on individual fitness, both BE and CE models can be used to explore the forces underpinning the dynamics of behaviour change over ecological scales (Mattison et al., Reference Mattison, Moya, Reynolds and Towner2018). Second, while social learning dispositions might not be directly relevant to explaining why a ‘low-fertility variant’ emerges in the population in the first place, as there is no existing model to imitate, CE models are not incompatible with BE models showing that, in environments where wealth is heritable and a better predictor of long-term fitness than fertility, material motivations could be selected over pure reproductive motivations (Rogers, Reference Rogers1995). Indeed, CE models view the emergence of a negative correlation between fertility and wealth as the starting point for the spread of low fertility. Third, with regards to the spread and maintenance of behaviour, BE and CE models could be offering alternative, independent or complementary explanations depending on whether the process of cultural transmission interacts with individual fertility trade-offs. While both approaches are often considered alongside each other (Borgerhoff Mulder, Reference Borgerhoff Mulder2009; Shenk, Reference Shenk2009; Alvergne et al., Reference Alvergne, Gibson, Gurmu and Mace2011; Shenk et al., Reference Shenk, Towner, Kress and Alam2013; Colleran et al., Reference Colleran, Jasienska, Nenko, Galbarczyk and Mace2014; Snopkowski & Kaplan, Reference Snopkowski and Kaplan2014; Colleran & Mace, Reference Colleran and Mace2015; Howard & Gibson, Reference Howard and Gibson2017; Mattison et al., Reference Mattison, Moya, Reynolds and Towner2018), integration remains limited.
(b) The adoption and spread of modern contraception
Most evolutionary accounts of contraceptive dynamics have focused on identifying the forces underpinning the adoption, or first use, of modern contraception. Studies have been conducted in various settings, from high-fertility to transitioning to low-fertility populations, and predictions from both BE and CE are helpful in advancing knowledge of contraceptive dynamics. From a BE perspective, it is not clear that the initial adoption of modern contraception is driven by the life-history trade-off between fertility and child survival (i.e. so-called quantity-quality trade-off, reviewed by Lawson & Borgerhoff Mulder, Reference Lawson and Borgerhoff Mulder2016): women working in the cash economy predominate among ever-users among the Pimbwe, Tanzania (Borgerhoff Mulder, Reference Borgerhoff Mulder and Rauch2009) and contraceptive use does not improve child survival among the Arsi Oromo, Ethiopia (Alvergne et al., Reference Alvergne, Lawson, Clarke, Gurmu and Mace2013). Some data rather suggest that contraceptive adoption and use are a response to strong life-history trade-offs between current and future reproduction, which can be increased owing to, e.g., maternal somatic depletion or changes in age at first birth, maternal education or maternal marital status. Modern contraception is often used to space births in high-fertility and African contexts (Mace et al., Reference Mace, Allal, Sear and Prentice2006; Borgerhoff Mulder, Reference Borgerhoff Mulder and Rauch2009; Alvergne et al., Reference Alvergne, Lawson, Clarke, Gurmu and Mace2013), and in rural Gambia, the technology is used to recuperate between births and improve body resilience for future reproduction (Bledsoe et al., Reference Bledsoe, Banja and Hill1998). Further, early adopters are more likely to be fertile women (Mace et al., Reference Mace, Allal, Sear and Prentice2006; Alvergne et al., Reference Alvergne, Lawson, Clarke, Gurmu and Mace2013) with higher parity for their age (Mace & Colleran, Reference Mace and Colleran2009; Alvergne et al., Reference Alvergne, Gibson, Gurmu and Mace2011, but see Borgerhoff Mulder, Reference Borgerhoff Mulder and Rauch2009), possibly suggesting that early adopters are more physiologically ‘depleted’ than others and/or on the highly fertile end of the spectrum.
From a CE standpoint, the slow spread of modern contraception in the early stages of the demographic transition points towards the absence of prestige-biased social transmission (Alvergne et al., Reference Alvergne, Gibson, Gurmu and Mace2011), suggesting that contraceptive uptake does not take off if the accumulation of cultural wealth (e.g. education) is independent from fertility (e.g. reducing fertility does not allow the gaining of wealth). Yet it is difficult to conclude the absence of social transmission when only semi-complete networks are available. There is also little evidence that kin influence goes against contraceptive uptake (the ‘kin influence hypothesis’; Newson et al., Reference Newson, Postmes, Lea and Webley2005): contraceptive use is either more likely among women with matrikin (in Tanzania; Borgerhoff Mulder, Reference Borgerhoff Mulder and Rauch2009), with extended kinship ties (in Thailand; Godley, Reference Godley2001) and whose female kin are users (in Poland; Colleran & Mace, Reference Colleran and Mace2015), or is independent from the presence of kin (in Gambia; Mace & Colleran, Reference Mace and Colleran2009). Non-parental transmission of contraceptive choices is found to be important further along the demographic transition, in rural Poland (Colleran & Mace, Reference Colleran and Mace2015) and Bangladesh (Gayen & Raeside, Reference Gayen and Raeside2010) for instance, and with modernisation (Kohler et al., Reference Kohler, Behrman and Watkins2001). Thus, evolutionary frameworks offer a strong theoretical backdrop against which to propose causal factors of change in the prevalence of contraceptive ever-users in each population.
(c) The discontinuation of modern contraception
There has been little attempt to harness evolutionary perspectives for understanding patterns of diversity and changes in the prevalence of contraceptive discontinuation within and among populations (Vitzthum & Ringheim, Reference Vitzthum and Ringheim2005). However, the discontinuation of contraceptive use is increasingly becoming one of the most significant concerns for family planning programmes: one in five ‘ever-users’ discontinue modern methods of contraception while still in need of family planning worldwide (Jain et al., Reference Jain, Obare, RamaRao and Askew2013). At the individual level, discontinuation while in need puts women at risk of unwanted pregnancies and unsafe abortion (Sedgh et al., Reference Sedgh, Singh and Hussain2014). At the population level, contraceptive discontinuation produces a ‘leaking bucket phenomenon’ (Jain, Reference Jain2014), whereby programmes aiming at increasing the prevalence of contraceptive use are hindered by contraceptive discontinuation. The reproductive consequences of contraceptive discontinuation while in need are substantial: more than 30 million unintended pregnancies occur among women not using contraception (Ali et al., Reference Ali, Cleland and Shah2012). Thus, following Jain's claim (Jain, Reference Jain2014) that family planning programmes would do better to concentrate on retaining existing users rather than focus on recruiting new clients, recent research and policy reports have suggested that tackling contraceptive discontinuation is central to achieving public health goals (Ali et al., Reference Ali, Cleland and Shah2012; Jain et al., Reference Jain, Obare, RamaRao and Askew2013; Castle & Askew, Reference Castle and Askew2015).
Possible reasons for the lack of discussion of contraceptive discontinuation in the evolutionary human sciences are: (a) studies of contraceptive use test hypotheses relevant to explaining the Darwinian puzzle of fertility decline rather than take contraceptive technologies as an object of study in its own right, and indeed theoretical work has researched the importance of biased social transmission for explaining the maintenance of low fertility in industrial populations (Ihara & Feldman, Reference Ihara and Feldman2004; Kendal and Ihara, Reference Kendal and Ihara Y2005; Ghirlanda & Enquist, Reference Ghirlanda and Enquist2007; Kolk et al., Reference Kolk, Cownden and Enquist2014); and (b) cultural evolution models are adapted from population genetics models (Cavalli-Sforza & Feldman, Reference Cavalli-Sforza and Feldman1981), in which a trait, once inherited or acquired through social transmission, is not typically lost at the individual level. Such models appear well adapted to long-acting, less easily reversible contraceptive methods (sterilisation, IUDs, implants) used for fertility limitation, but do not apply well to the use of short-acting methods, such as the pill or injectable, used for fertility timing/spacing; (c) Cultural evolutionary studies of fertility change tend to focus primarily on the social transmission of information rather than information obtained from one's experience (e.g. of contraceptive use). From a theoretical perspective, however, both CE and BE models can provide predictions for why and under which conditions women might discontinue contraception (Table 1). While some individuals might discontinue contraception in order to reproduce in line with their reproductive ideals, for others, contraceptive discontinuation while in need (e.g. because of side-effects) is likely to be at odds with a woman's fertility intentions. Below we outline how public health data on contraceptive discontinuation whilst in need can be analysed within an evolutionary framework.
3. Towards an understanding of the evolutionary dynamics of contraceptive discontinuation whilst in need
Why do individuals discontinue contraception despite wanting to delay or prevent pregnancy (i.e. whilst in need)? Women report multiple reasons for discontinuation whilst in need including method-related dissatisfaction due to concerns about side-effects (hormonal methods) and efficacy (condoms, withdrawal), fears or rumours that hormonal contraceptives cause cancer and/or infertility, partner's desired fertility, issues of cost and availability, as well as community-level contraceptive prevalence and attitudes (Campbell et al., Reference Campbell, Sahin-Hodoglugil and Potts2006; Bradley et al., Reference Bradley, Schwandt and Khan2009; Ali et al., Reference Ali, Cleland and Shah2012; Castle & Askew, Reference Castle and Askew2015). Whilst issues of cost, availability and partner's desires do influence women's decision to discontinue, they are much less frequently given as reasons for discontinuation for almost all method types than side-effects and health concerns (Bellizzi et al., Reference Bellizzi, Mannava, Nagai and Sobel2020), suggesting that the latter reasons are the most salient drivers of discontinuation for the majority of women. In this line, data from low- and middle-income countries show that contraceptive side-effects and health concerns are the leading cause of discontinuation whilst in need (Ali & Cleland, Reference Ali and Cleland1995; Blanc et al., Reference Blanc, Curtis and Croft1999; Castle & Askew, Reference Castle and Askew2015). Reported physiological side-effects from hormonal contraception include heavy bleeding, irregular bleeding, nausea, depression and loss of libido, amongst others. Injectable contraceptives are consistently cited as the method with highest rates of discontinuation due to side-effects and health concerns; however, qualitative studies show that women also report significant burdens of side-effects from pills, implants and IUDs (Meskele & Mekonnen, Reference Meskele and Mekonnen2014; Chebet et al., Reference Chebet, McMahon, Greenspan, Mosha, Callaghan-Koru, Killewo and Winch2015; Jain et al., Reference Jain, Reichenbach, Ehsan and Rob2017). A large body of qualitative literature show that women find side-effects highly problematic (Samandari & O'Connell, Reference Samandari and O'Connell2011; Meskele & Mekonnen, Reference Meskele and Mekonnen2014; Ochako et al., Reference Ochako, Mbondo, Aloo, Kaimenyi, Thompson, Temmerman and Kays2015; Capurchande et al., Reference Capurchande, Coene, Schockaert, Macia and Meulemans2016; Jain et al., Reference Jain, Reichenbach, Ehsan and Rob2017), as exemplified by the quote from a Ugandan woman: ‘I was bleeding so much, I thought I was going to die’ (Kibira et al., Reference Kibira, Muhumuza, Bukenya and Atuyambe2015).
From an evolutionary perspective, discontinuation due to the experience and/or fear of side-effects can be reframed as the outcome of ultimate causes and associated proximate mechanisms, in particular (a) evolved physiological mechanisms adjusting reproductive function to one's socioecological circumstances and causing variation in susceptibility to side-effects, (b) evolved cognitive dispositions adjusting reproductive behaviour, including discontinuation owing to side-effects, to context-specific fitness costs and benefits and (c) evolved cognitive dispositions underpinning social learning. Further, these forces might act independently or interact, e.g. the experience of side-effects, such as heavy bleeding or lack of bleeding, may fuel rumours about infertility. Below we outline how an evolutionary approach can shed new light on the dynamics of discontinuation due to side-effects (Table 1).
(a) Patterns of diversity in the experience of biological side-effects
With regards to contraceptive discontinuation, the question of why some women experience more side-effects than others has, to date, attracted little attention from either clinical trials (Inoue et al., Reference Inoue, Barratt and Richters2015), which focus on overall burden rather than interindividual variation, or public health, which concentrates efforts on dispelling misconceptions (Path, 2015). In contrast, reproductive ecologists have suggested that the risk of experiencing side-effects may be higher for women living in socioecological settings characterised by poverty, owing to low endogenous reproductive hormone levels relative to the contraceptive doses received (Bentley, Reference Bentley1994; Vitzthum & Ringheim, Reference Vitzthum and Ringheim2005). Indeed, previous anthropological studies have shown that levels of reproductive hormones vary significantly across cycles, between women and across populations (Jasienska & Jasienski, Reference Jasienska and Jasienski2008), with lower levels often (but not always) found in less affluent contexts (Vitzthum, Reference Vitzthum2009). In this line, an analysis of Demographic and Health Survey (DHS) data from Ethiopia shows that the risk of contraceptive abandonment is 42% higher among women living in poverty – an effect also reported in qualitative interviews e.g. ‘Only women having access to better diet [egg, meat, butter …] should take pills or injectables as it does not work for those having poor diet’ (Alvergne et al., Reference Alvergne, Stevens and Gurmu2017; see also Meskele and Mekonnen, Reference Meskele and Mekonnen2014). The perceived relationships between wealth and discontinuation owing to side-effects has also been shown in other settings such as Morocco, where women state they will use the pill when they can ‘afford to have a banana and glass milk from time to time’ (Mernissi, Reference Mernissi1975). In the same lines, using DHS data from Ethiopia, we found that anaemia increases the risk of discontinuing using contraception owing to side-effects (DSE), but not the risk to discontinue owing to other reasons (DOR), suggesting that poor somatic condition predicts the experience of negative side-effects (Stevens et al., Reference Stevens, Malbos, Gurmu, Riou and Alvergne2020). Yet the hypothesis that current doses of hormonal contraceptives may be maladjusted to some women's bodies has not been directly tested to date (Jasienska et al., Reference Jasienska, Bribiescas, Furberg, Helle and Núñez-de la Mora2017).
(b) Patterns of diversity in the experience of social side-effects
The experience of physiological side-effects can in turn elicit social side-effects, i.e. additional consequences that have a negative impact on social relations. The nature and severity of these effects depend on the local cultural context. For instance, variation in libido and mood changes have various implications depending on the cultural expectations of a woman's intimate relationships. In Kenya, women report that injectables make users ‘cold’, leading to a lack of sexual arousal and raising suspicions of infidelity from husbands (Ochako et al., Reference Ochako, Mbondo, Aloo, Kaimenyi, Thompson, Temmerman and Kays2015). Another side-effect that can have serious social consequences is disrupted menstruation, often subject to many local taboos and norms. In Mali, menstrual disruption can lead to accusations of witchcraft or immoral behaviour and can prevent a woman from preparing food and praying (Castle, Reference Castle2011). In Bengali society in Bangladesh, after menstruation, a woman must wait 7 days before she can resume her prayers. She must avoid sexual intercourse when menstruating, as there is a belief that contact with menstrual blood can jeopardise a man's sexual and spiritual integrity (Rashid, Reference Rashid2001). Such social side-effects can lead to reduced indirect paternal investment (e.g. investment in a child through support to the mother; Geary, Reference Geary2015), as well as loss of social support, which in turn decreases the potential fitness benefits of delaying reproduction. If side-effects are perceived to entail costs for one's future ability to conceive and/or in terms of loss of resources owing to social exclusion or decreased paternal investment, then the short-term costs of continuing contraceptive use might be perceived to outweigh the long-term benefits of using hormonal contraceptives to delay or space fertility. This is particularly relevant in some high-fertility contexts where hormonal contraception is primarily used to space births and conserve strength (Bledsoe et al., Reference Bledsoe, Banja and Hill1998; Alvergne et al., Reference Alvergne, Lawson, Clarke, Gurmu and Mace2013). Thus, physiological side-effects must be considered given the socio-cultural contexts of contraceptive users.
To understand diversity in the severity of social rejection owing to contraceptive use and associated side-effects, sexual conflict theory (Borgerhoff Mulder & Rauch, Reference Borgerhoff Mulder2009) might prove helpful. When the reproductive interests of men and women do not align, with men favouring higher fertility than women, conflict over contraceptive use and side-effects might ensue. One would predict that social side-effects might be more important in social systems associated with sexual conflict over fertility (stratified or unequal groups as compared to egalitarian groups; Borgerhoff Mulder & Rauch, Reference Borgerhoff Mulder2009). Further, sexual conflict over reproductive interests extends from husbands and wives to their kin, and thus one would expect the risk of discontinuation due to side-effects (DSE) to be modulated by the presence of kin. A promising avenue could be to compare DSE risk across and within social groups varying in their kinship ecologies and in degree of sexual conflict over fertility.
(c) From side-effects to contraceptive discontinuation: patterns of diversity in the tolerance of side-effects
From a behavioural ecology perspective, much of contraceptive discontinuation whilst in need can be seen as the outcome of the impact of side-effects on perceived fertility trade-offs. First, individuals face a life-history trade-off between current reproductive effort and future survival and reproduction. Thus, DSE can be understood as a form of investment in future survival and reproduction at the expense of current effort in offspring and own capital. Within this framework, DSE is expected to be more likely for women suffering the highest cost of side-effects for future survival and reproduction, for instance women at the beginning of their reproductive career: women who have never had a baby will be more likely to discontinue while older women will be more likely to tolerate side-effects. Conversely, if side-effects are perceived to threaten one's immediate chances of survival, which is more likely to be the case for women suffering the strongest side-effects, then the cost of continuing contraceptive use will outweigh the benefits of delaying or spacing fertility. In a study we conducted in Ethiopia, the experience and fear of physiological side-effects of the injectable contraceptive in the form of irregular and heavy bleeding led women to worry about significant threats to their health (the words used for heavy bleeding were synonymous with those used for ‘loss of life’) and their fertility (resulting from a delay to return to fertility after use or amenorrhea and/or a change in bleeding patterns while using). Thus, women experience various trade-offs depending on their circumstances and life-histories, and those need to be considered for predicting the risk to DSE.
Second, individuals face a life-history trade-off between fertility and survival of existing offspring (Lawson & Borgerhoff Mulder, Reference Lawson and Borgerhoff Mulder2016). If DSE is a form of investment in fertility at the expense of offspring quality, it might be more likely if alloparental care is available. Alloparental care is key to increasing family size (Sear & Coall, Reference Sear and Coall2011) and correlates with earlier reproduction among the Mosuo (Mattison et al., Reference Mattison, Scelza and Blumenfield2014), suggesting that alloparental care reduces the cost of reproduction. DSE is also potentially influenced by female reproductive competition. If side-effects interfere with marital relationships, and thus jeopardise women's ability to compete for paternal investment, then DSE might be more likely among polygynous households. Alternatively, if women use contraception to avoid the cost of a reproductive overlap with other women in the household (i.e. mother-in-law and co-wives, mother and sisters, depending on the social system), then tolerance to side-effects might be increased.
Third, the strength of such trade-offs is likely to vary across populations, between individuals and over the lifespan, depending on context (Uggla, Reference Uggla, Burger, Lee and Sear2020). For instance, individuals are expected to adjust their reproductive scheduling to the local rate of extrinsic mortality (i.e. a risk independent from individual phenotype and applying equally to all members of a population (Charnov, Reference Charnov1993), which vary across countries or neighbourhoods; reviewed in Uggla, Reference Uggla, Burger, Lee and Sear2020). When the risk of extrinsic mortality is high, the fitness benefits to delaying fertility are reduced and thus all else being equal, DSE might be more likely as compared with environments where child survival is highly dependent on parental investment. In addition, if the fitness benefits associated with investing in one own's capital depend on the subsistence economy (i.e. subsistence economy vs. competitive labour market economy), tolerance for side-effects in order to delay age at first reproduction is likely to be increased when education positively correlates with wealth. Macro-level features of the socioecology are thus also relevant to explaining patterns of DSE.
(d) Diversity in the fear of side-effects
Public health does not often make the distinction between women who experience side-effects and those who fear them. Rather, fear of side-effects is often referred to as misinformation, misconceptions or rumours, with an underlying assumption that many fears are not founded in real experience (Chipeta et al., Reference Chipeta, Chimwaza and Kalilani-Phiri2010; Hindin et al., Reference Hindin, McGough and Adanu2014; Path, 2015). In turn, dispelling myths (e.g. contraceptives lead to health problems; Gueye et al., Reference Gueye, Speizer, Corroon and Okigbo2015) through education remains the preferred strategy for tackling discontinuation. However, one must acknowledge that experience and rumour are not mutually exclusive and can often interact (Diamond-Smith et al., Reference Diamond-Smith, Campbell and Madan2012). Women's discussions of their own true experience of contraceptive side-effects end up creating fears based in the real experience of previous users. These rumours can then have significant impact on the rates of discontinuation. In this line, one study from three low- and middle-income countries (Nigeria, Kenya, Senegal) found a positive association between the aggregate level of method use and the prevalence of negative myths in a community in Nigeria (odd ratio, 1.6; Gueye et al., Reference Gueye, Speizer, Corroon and Okigbo2015) suggesting that myths originate from experiences. From an evolutionary perspective, this is not suprising, as humans (and other animals) rely on a complex mix of individual and social learning mechanisms to make decisions (Mesoudi, Reference Mesoudi2016). Understanding which mechanisms are at play in shaping cultural transmission in a given ecology is critical for predicting the dynamics of discontinuation over space and time.
Fear of side-effects when using hormonal contraception do appear to be prevalent for all methods and all groups of society (Path, 2015), suggesting that the motivation underpinning contraceptive discontinuation is culturally transmitted. In this line, the transmission of side-effect experiences and their impact on contraceptive behaviour has been documented in qualitative studies conducted in Kenya (Rutenberg & Watkins, Reference Rutenberg and Watkins1997; Kohler et al., Reference Kohler, Behrman and Watkins2001; Ochako et al., Reference Ochako, Mbondo, Aloo, Kaimenyi, Thompson, Temmerman and Kays2015), Egypt (DeClerque et al., Reference DeClerque, Ong Tsui, Abul-Ata, Abul-Ata and Barcelona1986), the Dominican Republic (Porter, Reference Porter1984) and Ethiopia (Alvergne et al., Reference Alvergne, Stevens and Gurmu2017). Women learn about side-effects from their social networks, and peers and other community members act as their main sources of information over and above health providers and other educational sources (Porter, Reference Porter1984; Rutenberg & Watkins, Reference Rutenberg and Watkins1997; Castle & Askew, Reference Castle and Askew2015; Ochako et al., Reference Ochako, Mbondo, Aloo, Kaimenyi, Thompson, Temmerman and Kays2015). Of course, the social transmission of contraceptive behaviour may be limited in some contexts owing to the high level of stigma associated with the practice (Alvergne et al., Reference Alvergne, Stevens and Gurmu2017). Yet, how fears about contraceptive side-effects and subsequent discontinuation behaviour may transmit through a network will vary based on social and ecological contexts. An evolutionary framework can be useful for making predictions about the dynamics of contraceptive discontinuation behaviour and transmission mechanisms in different contexts. To advance this research agenda and disentangle which processes are at play, we have suggested below questions needing investigation.
First, fears about infertility may persist as a cultural trait owing to pre-existing cognitive biases (Sperber, Reference Sperber1996). For instance, fears about contraception as a threat to future fertility is one of the most persistent and ubiquitous fears about modern contraception worldwide. This continuity may not be simply coincidental and may possibly be based on an existing cognitive bias to avoid risks to future fertility. Sperber (Reference Sperber1996) has argued that the stability of a cultural trait may emerge as people independently reconstruct representations based on pre-existing cognitive biases, or ‘attractors’ (Sperber, Reference Sperber1996; Morin, Reference Morin2015). This concept has been used across the disciplines of anthropology, psychology and linguistics to explain the surprising consistency of cultural traits (Buskell, Reference Buskell2017). Fears of contraception-caused infertility may be a cultural attractor, which may explain its ease of social transmission and surprising persistence across time and geography, despite education efforts within family planning programmes. This suggests that in order to counteract the spread of rumours, efforts should be directed at identifying which specific side-effects are associated with a fear of infertility and reducing them.
Second, the mechanisms underpinning the social transmission of contraceptive behaviours might vary as a function of local desired fertility norms. The saliency of fear of possible infertility will be stronger where the local cultural importance assigned to fertility is high, and where social influence mechanisms (e.g. frequency-biased social learning) might favour the spread and maintenance of discontinuation. This may be the case among women living in sub-Saharan Africa where the ideal family size is high, and where fertility decline appears to have stalled (Bongaarts & Casterline, Reference Bongaarts and Casterline2013). Further, if non-users or discontinuers are perceived to achieve higher wealth (unpublished interviews in Ethiopia reveal that irregular heavy bleeding owing to contraceptive use prevents some women from attending university or employment) and fertility, they may become seen as prestigious individuals and the spread of discontinuation through prestige-bias transmission might ensue.
Third, cultural transmission is also driven by social learning, where an individual's circumstances will interact with the process of diffusion. For instance, payoff-biased social learning, where an individual copies the behaviour with the highest observable payoff, leads to the quickest spread of the most successful techniques in a wild primate species (Barrett et al., Reference Barrett, McElreath and Perry2017). Learning about contraceptive side-effects is not independent from fitness related information as qualitative studies have shown that fears about infertility from contraceptive use are particularly present among young women with no children in contexts which place high value on fertility (Castle, Reference Castle2003; Adongo et al., Reference Adongo, Tabong, Azongo, Phillips, Sheff, Stone and Tapsoba2014; Hindin et al., Reference Hindin, McGough and Adanu2014; Ochako et al., Reference Ochako, Mbondo, Aloo, Kaimenyi, Thompson, Temmerman and Kays2015). Social network studies interacting individual characteristics with cultural transmission networks might be able to evaluate variation in how individuals make use of various learning strategies.
Applying evolutionary models to the study of contraceptive discontinuation can help provide explanations for patterns of discontinuation across cultural, social and other ecological contexts. Such models can be used to predict whether and when women discontinue hormonal contraception. Indeed, the timing of fertility – age at first birth, age at last birth and inter-birth intervals – is an important component to fitness as fertility poorly predicts fitness in a non-stationary population (Jones & Bird, Reference Jones and Bird2014). To date, however, scholarship on the cultural evolution of fertility decline has not sufficiently considered (1) that behavioural change is an incremental and dynamic process (e.g. adopt, stop, re-adopt), rather than the outcome of a binary choice, and (2) that the experience of contraceptive use, be it positive or negative, can be socially transmitted, bringing contraceptive adoption, discontinuation and switching in full circle. In Table 1, we draw on and integrate evolutionary approaches to propose future research questions and data to be collected for understanding patterns of discontinuation whilst in need, particularly due to side-effects. Note that before this can be done, however, a tool for measuring the severity of side-effects in a non-clinical setting is required for each context.
4. From contraceptive to fertility dynamics
Whilst contraceptive continuation is neither necessary nor sufficient to cause demographic transitions, and other methods of fertility regulation are important to fertility decline, contraceptive behaviour does have the potential to impact the pace of fertility decline. In this section we (a) review existing models of the relationship between the cultural evolution of contraceptive behaviour and demography, (b) sketch a new framework for modelling the evolution of contraceptive behaviour to consider change within an individual's lifetime and (c) discuss the implications of considering contraceptive dynamics and its impact on fertility for family planning policy. We argue that understanding how contraceptive uptake, discontinuation and switching between methods interact to shape fertility dynamics is necessary to provide a better understanding of how contraceptive behaviour influences fertility (Figure 1).

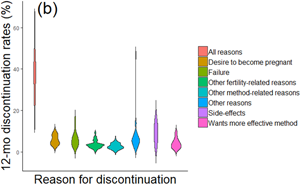
Figure 1. Contraceptive dynamics and fertility. (a) Scatterplot of total fertility rate and ever use of modern methods of contraception. (b) Violin plot of 12 month discontinuation rates by reason for all women using any modern method. Discontinuation due to side-effects is one of the top reasons for discontinuation. (c) Scatterplot of total fertility rate and 12 month discontinuation rate due to all reasons. (d) Scatterplot of total fertility rate and 12 month discontinuation rate due to side-effects. There is a strong positive correlation between total fertility rate (TFR) and (d) discontinuation due to side-effects, stronger than that between (c) TFR and both discontinuation due to all reasons and (a) contraceptive ever-use. Note that these are crude associations and additional multivariate analysis would help elucidate these effects further. Data is taken from aggregate DHS results, accessed via https://www.statcompiler.com/en/ on 21/04/20. A total of 31 low- and-middle-income countries were included in the analysis, with surveys spanning the years 1990–2015. For all countries included, only the most recent survey which had available data on all variables used was included. All data is for all (married and unmarried, results are the same for married only) women and only refers to modern methods. The R script and data are available in Online Appendix 1. R values indicate the results of Pearson's product-moment correlation tests. *** p < 0.001. ** p < 0.01.
(a) Existing models on the demographic impact of the cultural evolution of fertility
In the field of cultural evolution, previous studies of the interaction between demography and cultural evolution have aimed at understanding how changes in population size alter the evolution of cumulative culture and the maintenance of cultural complexity (Henrich, Reference Henrich2004; Ghirlanda & Enquist, Reference Ghirlanda and Enquist2007; Enquist et al., Reference Enquist, Ghirlanda, Jarrick and Wachtmeister2008; Ghirlanda et al., Reference Ghirlanda, Enquist and Perc2010). Only a few studies have proposed a mechanistic model to study the impact of cultural evolution on fertility and the demographic transitions (Ghirlanda & Enquist, Reference Ghirlanda and Enquist2007; Fogarty et al., Reference Fogarty, Creanza and Feldman2013; Kolk et al., Reference Kolk, Cownden and Enquist2014). Those have been concerned with explaining the maintenance of low fertility because of rapid cultural change that increases the amount of non-parental transmission. In those models, however, contraceptive dynamics at the individual level (adoption, switching between methods, discontinuation) are not considered. More generally, while discontinuation or ‘culture loss’ has been previously modelled (e.g. Henrich, Reference Henrich2004), it usually occurs at the population level and results from a cultural transmission error. Here we argue for the need to consider contraceptive dynamics at the individual level, which echoes other calls for a ‘systems approach to cultural evolution’ (Buskell et al., Reference Buskell, Enquist and Jansson2019).
In the public health literature, only a few statistical models have formalised the evolving relationship between individual contraceptive behaviour and population level outcomes (contraceptive prevalence and total fertility rate). Some focused on the impact of method mix on switching and contraceptive prevalence (reviewed in Blanc et al., Reference Blanc, Curtis and Croft1999). If increasing method mix enables more switching between methods and leads to less discontinuation, contraceptive prevalence is expected to increase (Castle & Askew, Reference Castle and Askew2015). Others investigated the reproductive consequences of contraceptive discontinuation (Blanc et al., Reference Blanc, Curtis and Croft1999, Reference Blanc, Curtis and Croft2002; Jain et al., Reference Jain, Obare, RamaRao and Askew2013). A study of DHS data from 15 countries (Blanc et al., Reference Blanc, Curtis and Croft1999) estimated that the total fertility rate would decrease by 20–48% in the absence of discontinuation while in need of contraceptives. However, those predictions do not consider the possibility either that negative experiences of contraceptive use are socially transmitted or that the risk to discontinue depends on individual circumstances. More generally, the mechanisms linking individual to group levels patterns are poorly understood.
(b) The evolution of contraceptive strategies
To better understand the mechanisms underpinning observed patterns of contraceptive discontinuation given socioecological contexts, it might be helpful to extend adaptive models of human reproductive decision-making to human contraceptive decision-making. When should one change contraceptive behaviour? Behavioural ecology and cultural evolution models potentially make different and complementary predictions for observed patterns. Following Mace (Reference Mace1998), one could consider a behavioural ecology state-dependent dynamic model on the assumption that contraceptive behaviour is adaptive. In such a model, parents would decide when to have another baby as a function of the environment they currently live in. Decisions would depend on the state of the parent, which could be described at any time by three variables: the type of contraceptive used (a proxy for the fitness cost of increasing the interbirth interval if the type of contraceptive causes side-effects perceived to have a reproductive health or fertility cost), the level of reproductive hormones (a proxy for the risk of experiencing side-effects) and the number of existing offspring. Optimal decision rules or contraceptive strategies would be those that maximise the number of grandchildren. The study of the discrepancy between observed and predicted patterns of discontinuation would inform on the importance of individual learning for contraceptive behaviour.
From a cultural evolution perspective, a classic population genetics model could be extended following Boyd and Richerson (Reference Boyd and Richerson1985) and Newson et al. (Reference Newson, Postmes, Lea, Webley, Richerson and Mcelreath2007) to understand the evolution of cultural norms given the impact of natural selection on the transmission of cultural variants. For instance, one could consider a polymorphic cultural trait, e.g. ‘belief that using modern contraception leads to infertility’ and ‘belief that using modern contraception improves fertility potential’ and investigate the impact of various transmission biases (e.g. cultural selection) during social interactions on the evolution of norms over successive generations. Which learning biases lead to the maximisation of individual fitness? Depending on the transmission mechanism at play, what is the shape of the diffusion curve? The study of the discrepancy between observed and predicted patterns of discontinuation would inform on the importance of social learning for contraceptive behaviour.
(c) The impact of contraceptive strategies on demography
Once evolutionary processes have been studied and mechanisms better understood, ecological processes at the timescale of a generation should be considered if one is to consider the impact of contraceptive behaviour on population demography. To that end, epidemiological-like compartment models, such as SIS (susceptible-infected-susceptible) models (Anderson & May, Reference Anderson and May1991), could present an interesting alternative to population genetics-like models (Cavalli-Sforza, L.L. & Feldman, Reference Cavalli-Sforza and Feldman1981) for the study of the evolution of culture and its ecological impacts (Cavalli-Sforza & Feldman, Reference Cavalli-Sforza and Feldman1981; Lewens, Reference Lewens2015). Compartments models would allow the consideration of contraceptive behaviour as a process, i.e. that individuals can change state throughout their lives, and if contraceptive behaviour is linked to vital dynamics (e.g. birth and death rates), an estimation of the impact of contraceptive dynamics on demography would be possible. A ‘multiple strains’ SIS model should be favoured to account for the diversity of contraceptive methods and estimate their ‘virulence’, i.e. how well they might spread. However, compartment models, in their simplest forms, are deterministic, and thus do not allow for the consideration of interindividual variability in ‘infection’ risk (i.e. adoption), ‘recovery’ (i.e. discontinuation) and interactions rates. To address those issues, epidemiologists have various tools at their disposal, including (a) the shift from deterministic to stochastic compartmental models, which introduce variability or white noise in parameters and (b) network models, which are also stochastic but offer a flexible framework for repeated contacts and for considering micro-level behaviour such as partner choice based on sex or age. Within network models, the classic ‘metapopulation’ approach, where interactions are more frequent within than between ‘patches’, can be used to divide the population according to sociological parameters (e.g. age, experience of side-effects, income, opinion; Kitchovitch & Liò, Reference Kitchovitch and Liò2011; Gargiulo & Huet, Reference Gargiulo and Huet2010). Another possibility is to use agent-based models, which probably offer the most flexible approach. Previous research suggests that stochastic epidemic metapopulation model and agent-based models give similar results, but the relevance of each approach depends on the particular aspect of the evolutionary dynamics to be studied and the availability of real-world data (Chen et al., Reference Chen, Small, Tao and Fu2018).
(d) Policy strategies
Modelling contraceptive dynamics as an evolutionary and ecological process provides a starting point to formally investigate the outcomes of various family planning strategies. This approach could be used to compare the efficacy of different strategies to promote the demographic transition such as encouraging switching (e.g. increasing the number of methods available) and improving contraceptive technology (e.g. to reduce side-effects) compared with investing in recruiting new adopters (e.g. acquiring new clients). The current public health default strategy is to increase modern contraceptive prevalence (mCPR) by focusing on counselling. For instance, the UNFPA cites the lack of knowledge and incorrect perceptions about the health risk of contraceptive methods as the main causes for discontinuation (UNFPA, 2019). In turn, many family planning programmes assume that providing full and consistent information about the safety of contraception will lead to 'rational' decisions (from the point of view of family planning programmes), leading people to ignore their fears of side-effects and take up contraception (Kaler, Reference Kaler2009). However, a strategy that focuses on health education about the safety of contraceptive use might not be effective if it contradicts users’ experiences.
One strategy often cited to reduce unmet need includes increasing switching between methods through augmenting method mix. FP2020 cites commitment to improving access to long-acting reversible methods (LARMs) (FP2020, 2017). Yet this strategy is not independent from the common metric used for evaluating FP programmes, couple years protection, which might favour LARMs (Darroch & Singh, Reference Darroch and Singh2011). Some authors have questioned the motives of LARM promotion in trying to achieve policy objectives over thinking about women's choice (Gomez et al., Reference Gomez, Fuentes and Allina2014). Recent studies also reveal that health providers sometimes push for LARMS despite women requesting using short-acting reversible methods (Yirgu et al., Reference Yirgu, Wood, Karp, Tsui and Moreau2020). Such a strategy might discourage women taking up contraceptives altogether, as it is often difficult to get long-acting methods removed (e.g. IUDs and implants).
An alternative strategy would be to improve current methods. It has been suggested that reducing side-effects may be achieved by changing either the dose of contraceptives or women's physiology. Vitzthum and Ringheim (Reference Vitzthum and Ringheim2005) note that there may be a biological basis (perhaps related to diet, nutrition or other metabolic factors) for variation in women's experience of side-effects and tolerance of hormonal contraception. In this line, women given much lower doses of oral contraceptives in both Latin America and Thailand had lower rates of irregular bleeding and few side effects, leaving them less likely to discontinue (Koetsawang et al., Reference Koetsawang, Charoenvisal, Banharnsupawat, Singhakovin, Kaewsuk and Punnahitanont1995). The recent roll out of Sayana Press, a lower-dose short-term acting injectable, might thus be a promising avenue in tackling discontinuation.
While many countries and family planning programmes focus on increasing mCPR, shifting the focus to tackling unmet need might prove both more efficacious and more ethical. Indeed, women who do not express a desire to limit family size or prefer to space births may be making ‘rational’ choices, from an evolutionary or biocultural perspective, given their local circumstances and the impact of the technology on their immediate health, resources and/or social circumstances. Future public health efforts should aim at providing women who need contraceptives with the right method for their goal (space, stop, delay) and their sociocultural and biological contexts (Cates & Maggwa, Reference Cates and Maggwa2014). As compared with pushing the family planning agenda to increase new adopters or promote only LARMs in order to achieve mCPR targets, providing the conditions and the acceptable technologies for women to invest in themselves and their offspring might be more effective in achieving the demographic transition or other policy goals such as decreasing infant mortality and improving child growth and maternal health.
5. Conclusions
Understanding patterns of contraceptive discontinuation has implications for predicting fertility dynamics and the demographic transition. Evolutionary scholars have yet to consider this issue when modelling the spread of low-fertility ideals, and public health models, aimed at estimating progress towards reducing unmet need for contraception (Kantorová et al., Reference Kantorová, Wheldon, Ueffing and Dasgupta2020), place the focus heavily on understanding adoption rather than continuation. In this paper, we call for evolutionary demography (Sear et al., Reference Sear, Lawson, Kaplan and Shenk2016; Sear & Burger, Reference Sear and Burger2020) and public health studies to go beyond adoption dynamics when modelling behaviour, because contraceptive discontinuation while in need opposes and possibly changes fertility ideals with demographic consequences. Indeed, fertility ideals are often renegotiated in light of changing circumstances (e.g. schooling of women; Behrman, Reference Behrman2015), and thus fertility ‘strategies’ are perhaps best understood as flexible (Trinitapoli & Yeatman, Reference Trinitapoli and Yeatman2018; Hruschka et al., Reference Hruschka, Sear, Hackman and Drake2019). We suggested ways in which an evolutionary framework can be harnessed to uncover the mechanisms underpinning patterns of discontinuation across groups, time and space. One of the most pressing types of data needed for this is a measurement tool for the prevalence and severity of side-effects in a non-clinical setting, which previously have only been documented qualitatively. We also outlined possible pathways for modelling both the biological evolution of flexible contraceptive behaviour strategies and the cultural evolution of modern contraception as a technology. While public health models aimed at assessing the demographic impact of alternative family planning strategies might gain from situating contraceptive behaviour within an evolutionary ecological framework, the evolutionary human sciences might also benefit from considering ‘culture loss’ or the reversibility of behaviour at the individual level more explicitly when modelling the cultural evolution of low fertility.
Considering behavioural discontinuation can also more widely inform evolutionary debates over the spread of low fertility. For instance, it has been proposed that institutions associated with monogamous marriage suppress intra-sexual competition and reduce total fertility, which lead them to be favoured by group selection because they promote success in inter-group competition (Henrich et al., Reference Henrich, Boyd and Richerson2012). This paper shows that individual learning from bodily experience (also referred to as embodied, somatic or visceral learning; Lewens, Reference Lewens2015), may, given individual socio-ecological circumstances, produce a transmission bias against the cultural evolution of low fertility. If individual learning constantly fuels within-group variability in fertility behaviour, then the conditions for cultural group selection (according to the Price equation, between-group variability should be larger than within-group variability; El Mouden et al., Reference El Mouden, André, Morin and Nettle2014) might not be met. Empirical data on the social transmission of discontinuation are needed to evaluate this possibility further.
Following others who have argued that cultural evolution need not imply group selection (Amir et al., Reference Amir, Jordan and Randb2016), we find that culture is more than socially transmitted information and can evolve following changes in life-history trade-offs (owing to e.g. the use of a particular technology, a change in individual or socio-ecological circumstances). This is not to say that social transmission is not crucial (see Whiten et al., Reference Whiten, Caldwell and Mesoudi2016 for a review of cultural diffusion in humans and other species). The idea that an individual negative experience can counteract cultural pressure has been raised by others in a formal model showing that cultural evolution that reduces genetic fitness can be evolutionarily unstable (El Mouden et al., Reference El Mouden, André, Morin and Nettle2014). Considering behavioural traits as practices that can be discontinued, and understanding the conditions under which discontinuation occurs, might have implications for understanding the evolution of other costly behaviours or Darwinian puzzles. The commonality of discontinuation behaviour remains to be investigated, but we hope that this paper will encourage future studies within the fields of cultural evolution and behavioural ecology to go beyond adoption dynamics when studying the evolutionary process of cultural change.
Conflicts of interest
AA is on the editorial board of Evolutionary Human Sciences.
Authors’ contributions
AA conceived the initial version of the paper. AA and RS wrote the paper. All authors gave final approval for publication.
Acknowledgements
We thank two anonymous referees for very detailed and insightful feedbacks on this manuscript. We are also grateful to Rebecca Sear for her encouragement, Sylvain Billiard for being an original force behind the emergence of this paper and for stimulating discussions around cultural ecology, and Nichola Raihani for helpful feedbacks on an earlier version of this manuscript. ISEM no. 2021-023.
Financial support
AA was funded by a Returning Carer Fund from the University of Oxford VC Fund. RS is funded by a joint ESRC and Clarendon PhD scholarship.
Research transparency and reproducibility
The data and R script for producing Figure 1 are available from https://figshare.com/s/d3492feab9aceddd2001



 Open access
Open access