


INTRODUCTION
Campylobacteriosis is the most frequently reported human bacterial enteric disease in most developed countries [1]. The symptoms of campylobacteriosis in humans include mild to severe diarrhoea, abdominal pain, nausea, malaise, with the occasional complication of Guillain–Barré syndrome [1]. Many domesticated and wild animals, including birds and insects can carry several species within the genera Campylobacter. Campylobacter jejuni subsp. jejuni (C. jejuni) is the most frequently isolated species in chickens (and also frequently isolated from sheep and cattle faeces) with Campylobacter coli being most frequently isolated from pigs [Reference Munroe, Prescott and Penner2]. However, broiler chickens from industrial poultry production are regarded as being the main reservoir for C. jejuni, which is the most frequent Campylobacter subtype isolated from infected humans [3]. The transmission from animals to humans is via the oral route, either by consumption of faecally contaminated meat or by direct contact with feces by handling animals, soil or water (occupational hazard). Therefore, control of Campylobacter during primary broiler production is expected to have a greater benefit for public health than control measures later in the production chain, as spread from animals to humans can occur within the farm by other ways than poultry meat [3].
Worldwide, the incidence of human campylobacteriosis has increased considerably during the last decade. Many studies were conducted to understand the pattern and cause of this increased incidence in humans. An increased interest for Campylobacter in the scientific and medical community might have led to the increase in number of reported human cases. The rise in consumption of fresh chicken (and thereby also handling of raw meat) in Western countries was seen by many as the main reason for increased incidence in humans [4, Reference Guerin5]. Due to its importance for public health, several countries have taken large- or small-scale protective measures to control Campylobacter in chickens and hence in humans. Emphasis has been placed on reducing the prevalence of positive flocks pre-harvest and on reducing the number of bacteria on positive carcasses by applying measures like heat treatment and freezing [Reference Rosenquist6].
Campylobacter is a bacteria living in the intestinal tract of poultry, without harming them (commensal). There is frequently a lag-time between introduction (time) and spread/detection of the infection in a flock. However, transmission within a flock is regarded as being very quick, the speed depending on whether Campylobacter is introduced by one, few or several vectors (flies, drinking water, people). By the time a colonized flock is sent for slaughter, the majority of the chickens are colonized [Reference Newell and Fearnley7]. Before introducing a new flock, the houses are cleaned and disinfected to the extent that no Campylobacter should survive.
As mentioned above, many domesticated and wild animals could be regarded as a source of campylobacteriosis in humans [3]. An available control option is therefore to block transmission of Campylobacter from the environment into naive flocks. Many risk-factor studies have therefore been conducted in order to shed light on the spatial, temporal and management factors involved in Campylobacter colonization of broiler flocks. Several spatial and management factors have been evaluated and identified, including poor hygiene practices on the farm, contaminated water, presence of other farm animals on the farm, more than one house on the farm, surroundings with a grove (small group of trees) close to the farm, presence of pig herds on neighbouring farms and season [Reference Kapperud8–Reference Hansson11]. The temporal variation in Campylobacter infection in humans and animals is well documented with the highest prevalence during the summer period [Reference Kapperud8, Reference Meldrum12–Reference Jore14]. The higher prevalence in summer has been suggested to be due to better survival conditions for Campylobacter in the environment at high temperatures [3, Reference Jore14]. However, the spatial dependence of Campylobacter colonization in broilers is not well documented as temporal dependence.
Methodology commonly used to investigate spatial dependence includes analysis of spatial and spatio-temporal clustering and geostatistics. Scan statistics is a widely used technique for identification of local clusters (an area in space with higher or lower disease occurrence compared to expected numbers if the disease was randomly distributed in space). The K function is another commonly used technique for estimating global spatial clustering (identifying second-order spatial effects). Semivariogram models (a technique within geostatistics) can be used to estimate the range of spatial dependence (range of influence). These methods are particularly useful to generate hypotheses, i.e. investigating whether the distance between chicken flocks, at which transmission of Campylobacter is possible, is related to the dispersal capability of vectors like flies. Some studies have observed spatial variation in Campylobacter colonization on broiler farms in Norway [Reference Jonsson15], without being able to link spatial clustering to any factors being handled particularly different in the different regions. However, this information could be combined with other information from sources like the Quality Assurance System in Danish Broiler Production (KIK system) database in slaughter chickens and other gathered information about high- and low-risk areas. Examples of this kind of information are the presence of other animals on the farm or on neighbouring farms, cooperation or common ownership, and using a common processing plant.
In Denmark, a voluntary national surveillance programme for Campylobacter in broiler flocks has been active since 1998 based on faecal swabs collected from the floor of the chicken house (mandatory since 1 January 2010). At the processing-plant level, cloacal swab samples are collected from ten randomly selected birds from each flock and then pooled in the laboratory. Since 2003, the collection of these swab samples has been mandatory [Reference Rosenquist6]. By testing the pooled samples, the overall status of Campylobacter for each flock is seen as being efficiently estimated [Reference Hansson11]. Subtyping of positive isolates is not routinely conducted. In 2008, a new 5-year action plan against Campylobacter was implemented by the Danish Government with the aim of reducing the prevalence of Campylobacter in Danish broiler flocks and meat further down the production chain. There has been a decrease of Campylobacter-positive broiler flocks in Denmark from 38% in 2002 to 29% in 2006 [16]. From 2007, however, there has been no further reduction in the prevalence of Campylobacter-positive flocks.
The aim of this study was to contribute to the further reduction of Campylobacter colonization in broiler farms in Denmark. In this context we began by exploring the spatial and temporal variation of Campylobacter colonization in broilers in Denmark. The knowledge gained on the spatial patterns will then be used, together with risk-factor data collected from other sources, to categorize the farms into different risk profile groups. Advice specifically tailored for the risk profile of each farm can then be given.
The objectives of this study were therefore to study the spatial and temporal patterns in order to identify potential high-risk areas, spatio-temporal clusters and to estimate whether there was any statistically significant range of influence between the farms. The latter would indicate if vectors, sharing of equipment and/or labour would be likely spatial-explanatory factors for the introduction of Campylobacter into broiler flocks.
Spatio-temporal patterns in Campylobacter colonization in Danish broilers remain unexplored. Together with hypothesis generation on explanatory factors that might be responsible for colonization, identification of high-risk areas will facilitate a more focused surveillance and increased safety measures. Furthermore, producers can be advised regarding safety precautions (e.g. strengthening of on-farm biosecurity) with regard to the presence of colonized farms within the estimated range of influence.
MATERIALS AND METHODS
Study design
At the Danish broiler processing plants cloacal swab samples were collected from ten randomly selected birds of each flock. These ten samples were pooled in the laboratory into one sample and analysed by polymerase chain reaction (PCR) as described by Lund et al. [Reference Lund17] to detect the presence of Campylobacter in the sample. All flocks sent for slaughter were classified as Campylobacter positive or negative. The dataset used for the present study included Campylobacter test results at the flock level from 2007 to 2009, laboratory test date, season (winter, spring, summer, autumn) and geographical coordinates at the farm level. The original dataset included test results from all flocks of 236 farms in 3 years (January 2007 to December 2009). Ten farms sent less than five flocks for slaughter in the 3 years and most of them were clustered in time (i.e. were sent for slaughter within the same year). To avoid the influence of this artificial temporal clustering of sample submission, those ten farms were excluded from the analysis. For some flocks there was more than one test result (due to partial slaughtering or using more than one processing plant for large flocks). To avoid double counting of a flock, laboratory results were aggregated to one test result if there were fewer than 29 days between slaughters. The aggregated test result was positive if at least one of the tests was positive. The final dataset included 226 farms with 10 680 flocks.
Analytical methods
Visualization of spatio-temporal pattern
Risk mapping was used to visualize spatio-temporal patterns of Campylobacter cases in the four different seasons across years and during summer in different years separately. The prevalence in four different seasons across years was calculated as the number of test-positive flocks out of the flocks submitted by each farm in each season across the period of study. The prevalence in summer in different years was calculated as the number of test-positive flocks out of the flocks submitted by each farm in each year during summer. The kernel density estimation (KDE) method was used to derive a smooth surface over the point locations. The kernel density function generates a point density surface over the point locations of the events [Reference Pfeiffer18]. The kernel density was calculated using a fixed-sized window centred on each data point. A bivariate probability density function was applied to determine the concentration (mean number of events per unit area) of a spatial point process. The degree of smoothing depends on the size of the window as more information will be drawn from the adjacent areas using a bigger window. In the present study, different-sized bandwidths and grid cells were assessed by visual inspection and it was found that using a 15-km bandwidth and 1-km resulting grid cell created the best surface to describe the underlying point pattern. A bandwidth >15 km produced a very smooth surface which calculated density estimations over spaces with no population. A bandwidth <15 km created less smooth surface which resembled the point locations. The analysis and visualization were performed using ArcMap (ArcGIS version 9.1, ESRI Inc., USA).
Global clustering
K function analysis was performed in order to detect global clustering. K function analysis is testing if the spatial distribution of the colonized locations estimated by the empirical K function deviates from complete spatial randomness. Complete spatial randomness is often modelled using a homogeneous or inhomogeneous Poisson process. However, if the spatial distribution of the farm location (independently of colonization status) does not follow a Poisson process the test will fail. Initially a test was performed to evaluate if the spatial distribution of farm locations deviates from a homogeneous Poisson distribution [Reference Ersbøll and Ersbøll19]. This was done by testing if the number of flocks within each county follows a Poisson distribution. The country is divided in regions (i.e. islands and a peninsula). Each region is divided into counties. As the counties are different in size, the intensity of the Poisson distribution is proportional to the size of the area of the county:
 $$N_{{\rm producers\comma }i} \in P\lpar \alpha \vert A_{i} \vert \rpar \comma \hfill$$
$$N_{{\rm producers\comma }i} \in P\lpar \alpha \vert A_{i} \vert \rpar \comma \hfill$$where α is the average density of farms (number of farms/km2) and |A i| is the size of county i. For each county i, the quantile corresponding to the actual number of farms in a Poisson distribution given by equation (1) is recorded. A QQ plot of quantiles for all counties should follow a uniform distribution. The analysis was performed for one region, the Jutland peninsula.
A modified K function was used to detect global clustering as described by Ersbøll and Ersbøll [Reference Ersbøll and Ersbøll19]. The modified K function overcomes the assumption of a specific distribution characterizing complete spatial randomness of the point locations of the event (e.g. a homogeneous or inhomogeneous Poisson distribution). The null-hypothesis K function describing complete spatial randomness is derived using Monte Carlo simulation. The analysis was performed for the summer season each year separately. A farm was defined as positive for the season of the year if at least one flock of the farm tested positive in that season. The K function was estimated as:
 $${\widehat{K}}\lpar h\rpar \equals \sum\limits_{i \equals \setnum{1}}^{n} {\sum\limits_{j \equals \setnum{1}}^{n} {I_{h} \lpar d_{ij} \rpar \quad \lpar h \gt 0\comma i \ne j\rpar } } \hfill$$
$${\widehat{K}}\lpar h\rpar \equals \sum\limits_{i \equals \setnum{1}}^{n} {\sum\limits_{j \equals \setnum{1}}^{n} {I_{h} \lpar d_{ij} \rpar \quad \lpar h \gt 0\comma i \ne j\rpar } } \hfill$$where h is the distance, n is the number of colonized farms, I h(d ij) is an indicator function taking the value 1 if d ij<h and 0 otherwise. The empirical K function, K 1(h) was calculated using equation (2) with the actual location of n test-positive farms. A simulated null-hypothesis K function, K 0(h), was derived from a random sample of n farms among the total number of farms, assuming these n farms were test-positive. A total of 999 simulations was used. The difference D(h) between the empirical and simulated K function was plotted vs. the distance, h, together with the 95% simulation envelop. If the D function crosses the 95% envelop, it indicates global clustering. SAS version 9.1 (SAS institute Inc., USA) was used to perform the analysis.
Local spatio-temporal clusters
Presence of local spatio-temporal clusters was evaluated using spatio-temporal scan statistics. Analysis was performed for each year, separately. A farm was considered positive in a month in a specific year when one or more flocks from that farm were found positive during that month in the specific year. Spatio-temporal scan statistics as implemented in SaTScan v. 8.2.1 [Reference Kulldorff20] was used. A series of cylindrical windows with different radius and height (from 0 to a specified percentage of the total population and time) was created over the study area and period of interest. The base of the cylinder centred on each farm location represents space and the height of the cylinder represents time. The risk of the disease within each window was compared with the risk outside the window [Reference Kulldorff20]. A Bernoulli model was used as colonization status of each of the data locations was regarded as either positive or negative. The maximum size of the temporal window and population at risk included in a cluster varied from 10% to 50%. No geographical overlapping between clusters was allowed in the analysis. Clusters obtained by Monte Carlo hypothesis testing with 999 permutations were considered significant when P⩽0·05.
Range of influence
To estimate the range of influence of test-positive farms, semivariogram analysis was applied. The empirical semivariogram was calculated using a robust model suggested by Cressie & Hawkins [Reference Cressie and Hawkins21] as:
 $$\eqalign{\tab 2\bar{\gamma }\lpar h\rpar \equals \cr \tab \left\{ {{1 \over {\vert N\lpar h\rpar \vert }}\sum\limits_{N\lpar h\rpar } {\vert Z\lpar S_{i} \rpar \minus Z\lpar S_{j} \rpar \vert ^{{\setnum{1} \over \setnum{2}}} } } \right\}^{\setnum{4}} \sol \lpar 0{\cdot}457 \plus 0{\cdot}494\sol \vert N\lpar h\rpar \vert \rpar \comma}$$
$$\eqalign{\tab 2\bar{\gamma }\lpar h\rpar \equals \cr \tab \left\{ {{1 \over {\vert N\lpar h\rpar \vert }}\sum\limits_{N\lpar h\rpar } {\vert Z\lpar S_{i} \rpar \minus Z\lpar S_{j} \rpar \vert ^{{\setnum{1} \over \setnum{2}}} } } \right\}^{\setnum{4}} \sol \lpar 0{\cdot}457 \plus 0{\cdot}494\sol \vert N\lpar h\rpar \vert \rpar \comma}$$where Z(S i) and Z(S j) are the Campylobacter prevalence for farms i and j at locations S i and S j separated by the distance h. N(h) denotes all pairs of neighbouring farms at distance h and |N(h)|denotes the number of distinct pairs in N(h).
An exponential semivariogram model was fitted using nonlinear regression to obtain estimates for the range of influence (a) of the prevalence of Campylobacter colonization between farms as:
 $$\widehat\gamma \lpar h\rpar \equals c_{\setnum{0}} \plus c_{\setnum{1}} \exp \left( {{h \over {3a}}} \right)\quad {\rm for\ }h{\rm \gt 0} \comma$$
$$\widehat\gamma \lpar h\rpar \equals c_{\setnum{0}} \plus c_{\setnum{1}} \exp \left( {{h \over {3a}}} \right)\quad {\rm for\ }h{\rm \gt 0} \comma$$where 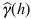 $\widehat\gamma \lpar h \rpar$ is the fitted semivariogram, h is the distance, c 0 and c 1 are the nugget effect and partial sill, respectively. The practical range of influence (a′=3 a) was calculated as three times the estimated range of influence. The prevalence of Campylobacter colonization during summer for each farm per year was used for fitting the semivariogram model. The nugget effect (c 0) is an estimate of sampling error and short-scale variability. The semivariogram usually increases up to a limit where it becomes stable. The maximum variance that can be attained by the variogram is the partial sill (c 1). The lag distance at which the sill occurs is called the range of influence (a′) which indicates the average distance between locations at which Campylobacter colonization is not correlated. Semivariograms were modelled for prevalence each month and each season of the year separately. We used SAS version 9.1 (SAS institute Inc., USA) to conduct this analysis.
$\widehat\gamma \lpar h \rpar$ is the fitted semivariogram, h is the distance, c 0 and c 1 are the nugget effect and partial sill, respectively. The practical range of influence (a′=3 a) was calculated as three times the estimated range of influence. The prevalence of Campylobacter colonization during summer for each farm per year was used for fitting the semivariogram model. The nugget effect (c 0) is an estimate of sampling error and short-scale variability. The semivariogram usually increases up to a limit where it becomes stable. The maximum variance that can be attained by the variogram is the partial sill (c 1). The lag distance at which the sill occurs is called the range of influence (a′) which indicates the average distance between locations at which Campylobacter colonization is not correlated. Semivariograms were modelled for prevalence each month and each season of the year separately. We used SAS version 9.1 (SAS institute Inc., USA) to conduct this analysis.
RESULTS
Descriptive analysis
In this study, the total number of flocks sent to processing plants by each farm in the 3-year period ranged from six to 393 (median 44 flocks).The number of test-positive flocks for each farm having positive flocks in summer ranged from one to 64 (median four flocks). The number of farms with at least one test-positive flock in different seasons across the 3 years was 99 (44%), 81 (36%) 199 (88%) and 177 (78%) in winter, spring, summer and autumn, respectively. During the summer season the number of farms with at least one positive flock was 158 (72%), 141 (66%) and 118 (57%) in 2007, 2008 and 2009, respectively. Flock-level prevalence of Campylobacter for each farm was used to visualize the temporal pattern in Figure 1.

Fig. 1. Kernel-smoothed maps of the spatio-temporal patterns regarding prevalence of Campylobacter colonization in Danish broilers 2007–2009. (a) Winter, (b) spring, (c) summer, (d) autumn. Darker shading indicates higher prevalence.
The location of the farms and the spatial distribution of the density of farms are shown in Figure 2. Visual evaluation of the map indicates differences in the density of the distribution of the farms. Among 226 farms, eight (4%) farms never produced any positive flock during the study period. Four out of these eight farms are located in two different areas with high farm density but low prevalence. The locations of the remaining four farms are scattered outside high farm-density areas.

Fig. 2. Kernel-smoothed map of the spatial distribution of broiler farms in Denmark 2007–2009. Darker shading indicates higher density.
The spatial and temporal variation of the prevalence of Campylobacter colonization is shown in the kernel-smoothed maps in Figure 1 for each season during the 3 years. The southeastern and northeastern parts of Jutland and the western part of Bornholm showed a high prevalence throughout different seasons. In almost all areas, summer was the season with the highest prevalence, while few areas were colonized during winter and spring.
Spatial analysis
An initial analysis showed that the farm locations (independently of colonization status) significantly deviated from a homogeneous spatial distribution. The modified K function analyses did not detect significant global clustering (D function within 95% simulation envelops) for any of the years.
Spatio-temporal scan statistics identified significant spatio-temporal clusters of Campylobacter colonization during the summer months in Denmark from 2007 to 2009 (Fig. 3, Table 1). Inclusion of a maximum 10% of the population at risk and 20% of the time periods resulted in significant clusters. In 2007, three significant clusters were identified. The primary cluster was located in the northern part of Jutland and the secondary clusters in the southern part of Jutland. In 2008, the primary cluster was located in the same area as in 2007. Secondary clusters were identified in the southern and northern parts of Jutland. In 2009 a primary cluster was identified in the northern part of Jutland, north to Limfjord, and secondary clusters were identified in the same places in other years. All clusters were identified either in July or August (summer in Denmark). Identified clusters substantially coincided with the underlying prevalence of the area (Fig. 3).

Fig. 3. Location of significant primary and secondary spatio-temporal clusters of Campylobacter colonization in Danish broilers. (a) 2007, (b) 2008, (c) 2009. Thick circle indicates primary clusters. Prevalence during summer is shown using kernel density estimation as background, where darker shading indicates higher prevalence.
Table 1. Statistically significant spatio-temporal clusters (P⩽0·05) for Campylobacter test-positive broiler farms

RR, Relative risk.
An exponential semivariogram was fitted to the empirical semivariogram of the Campylobacter prevalence during the summer period each year (Fig. 4). The range of influence was estimated as an average measure of the distance between farms at which Campylobacter colonization is not correlated. In 2007, the range of influence was estimated at the distance of 9·6 km (s.e.=4·2 km) and in 2008 and 2009, 12·9 (s.e.=5·7 km) and 13·5 km (s.e.=5·4 km), respectively. The semivariogram analyses for the separate months and seasons (data not shown) showed that the estimated range of influence was only significant during the summer months and the summer season.

Fig. 4. Empirical (•) and fitted semivariogram (—) of Campylobacter colonization in Danish broiler farms in summer. (a) 2007, (b) 2008, (c) 2009. The range of influence was estimated at distances 9·6 km (s.e.=4·2 km), 12·9 km (s.e.=5·7 km) and 13·5 km (s.e.=5·4 km) in 2007, 2008 and 2009, respectively.
DISCUSSION
While temporal variation in Campylobacter colonization in Danish broilers is well documented [Reference Wedderkopp22, Reference Bang23], the spatial variation of the colonization is still not well understood. In our study we identified significant spatial variation in Campylobacter occurrence during the summer period (presence of significant spatio-temporal clusters; Fig. 3, Table 1).
Several hypotheses can be drawn to explain this spatial variation in the risk of colonization. The presence of spatial risk factors could act as a driver for the spread of the bacteria in an area. A possible significant spatial factor may be the presence of colonized broiler farms within a certain radius of a susceptible farm. Spread of infection from a colonized farm or from the environment contaminated by colonized farms in the neighbourhood (such as spread of contaminated faeces on surrounding fields) to a susceptible farm can be facilitated by humans, feed trucks and other kind of vehicles, birds and flies, of these flies are regarded as the most important source [Reference Hald24]. In Denmark visitors are not allowed onto farms. Therefore, there is a reduced risk of spreading bacteria by this means. However, farmers who own more than one farm could spread the bacteria between farms or workers could introduce the bacteria to the farm from a colonized environment, given poor on-farm hygiene and biosecurity practice. Flies and birds could spread bacteria between farms more easily than people, although this hypothesis needs to be studied more thoroughly. Flies have a flying range within which they may influence the spread of bacteria around a colonized farm and to farms located nearby. Some studies have shown that the presence of other farm animals within a 1 km radius of a broiler farm is a significant risk factor for Campylobacter colonization [Reference Bouwknegt9, Reference Lyngstad10]. Bacteria could also spread through shared equipment between farms and disposal or spreading of manure.
Spatial variation in climatic factors (temperature, humidity, etc.) might also influence the distribution of an infectious disease. Variation in temperature, humidity or soil texture influences the survivability of an organism and its biological vectors. Significant clusters of farms with Campylobacter-positive broilers were repeatedly detected in the same areas of Jutland during summer (Fig. 3). To explain the occurrence of the clusters in the same area over the years, an extended study with inclusion of suspected spatial and temporal risk factors is needed.
The range of influence indicates that a colonized broiler farm is spatially correlated with other colonized farms up to 9·6–13·5 km, depending on the year (Fig. 4). The dispersal ability of a housefly is around 5–7 km [Reference Nazni25] which is within the 95% confidence interval for the estimated range of influence in our study. Birds fly much greater distances and could also be one of the factors influencing this range. The practice of sharing equipment between farms and the strategy of disposal and spreading of manure onto fields could also spread bacteria over shorter distances. Processing plants use their own trucks and cages to carry birds from farm to processing plant. Improper cleaning and drying of cages and trucks between the collections of birds from different farms could spread bacteria over longer distances. The range of influence was only statistically significant in summer, which may be due to an increase of statistical power as most farms become colonized during summer. Ersbøll & Nielsen [Reference Ersbøll and Nielsen26] have previously reported that when prevalence was below 5% and/or the number of test-positive flocks was low, there is a large standard error for the spatial parameter estimates in the semivariogram model. Moreover, higher abundance of flies and birds, and other related seasonal factors could also trigger clustered occurrences of colonization in space which might influence the result in summer. The estimated range of influence for each year was different although not significantly as the standard error intervals overlap.
We detected a marked seasonal pattern of Campylobacter colonization in broilers (Fig. 1), corroborating what has been documented in many studies in humans and other animals [Reference Kapperud8, Reference Meldrum12–Reference Jore14, Reference Stanley and Jones27–Reference Wallace29]. In our study, the semivariogram model showed significant spatial correlation only in summer, which further supports the influence of season for this colonization. Moreover, significant spatio-temporal clusters can only be identified during summer. Several hypotheses have been proposed to explain this seasonality. Jore et al. [Reference Jore14] showed that temperature is highly correlated with incidence of Campylobacter in broilers. Wallace et al. [Reference Wallace29] also showed that there is a positive correlation between infection in broilers and minimum and maximum temperatures. A higher temperature implies a higher survival of Campylobacter in the environment but also a greater abundance of flies and other insects that act as either mechanical or biological vectors [Reference Hald24, Reference Hald, Skovgård and Pedersen30]. The influx of insects in broiler houses depends on the amount of ventilation needed and therefore also on the outdoor temperature [Reference Hald24]. Moreover, shedding of thermophilic Campylobacter in fresh faeces of dairy cattle also increases during summer which makes the organism available for flies in the environment during this specific period [Reference Stanley31].
In some areas, like northern Jutland, north of Limfjord, where the density of farms is high (Fig. 2), the prevalence was also high in all seasons, not just summer. There were also some cases where the opposite happened, the density of farms was high but the prevalence was low in winter and spring although high in summer, e.g. south of Limfjord. Although both areas had a high farm density, the influence of season or other factors on prevalence is higher south of Limfjord than to the north. Moreover, it is probable that spatio-temporal clusters in 2007 and 2008 were detected in an area with low farm density. Farm density, therefore does not seem to be a determinant for high prevalence. Nevertheless, four of the eight farms that never delivered any positive flocks during the study period were situated in the two different high farm-density areas where prevalence was low during different seasons. Other factors like poor hygiene on farms, abundance of vectors in the area, density of colonized farms may be involved in the maintenance of high prevalence during the year in areas where the density of farms is high.
We used prevalence of colonization in different seasons to visualize the spatio-temporal pattern in maps (Fig. 1). The inhomogeneous pattern of farm distribution (Fig. 2) in space might influence the visual assessment of the distribution of prevalence. Moreover, prevalence and spatio-temporal visualization will be influenced by the number of flocks that the farm rears during a season. A farm delivering only two flocks for slaughter in a season where both turn out to be positive would obtain a prevalence of 100%. In cluster analysis, spatio-temporal clusters of colonization were identified within the same regions indicated as high-prevalence areas by the kernel-smoothed maps of prevalence during different seasons across years and summer in different years (Figs 1, 3). The spatial pattern shown by the kernel-smoothed maps of prevalence might be validated by cluster analysis and vice versa in the present study. Some studies suggested using more than one technique to analyse spatial disease distribution in order to increase the power of the cluster analysis [Reference Ward and Carpenter32, Reference Ward33]. However, Song & Kulldorff [Reference Song and Kulldorff34] described spatial scan statistics as a powerful analytical tool to detect localized clusters compared to other techniques. It should be noted, however, that in our spatio-temporal cluster analysis a farm was regarded as positive if at least one flock was found positive in a month for that farm. Farms delivering more flocks to processing plants per month had therefore a higher probability of becoming positive for at least one flock, which might have incorporated some bias into the study. An analysis for further understanding was conducted to identify if the number of chicken houses (a proxy of size of the farm) is different between farms within and outside space–time clusters, turned out significant. However, number of chicken houses on the farm is a significant risk factor for Campylobacter colonization in broilers [Reference Lyngstad10, Reference Refregier-Petton35]. It is difficult to conclude from this analysis about what percentage of bias and the effect of number of houses has influenced the identified clusters.
No significant global clustering was present in our study. Even though we used data from 226 broiler farms (which is close to the total number of broiler farms in Denmark), it is possible that the small number of data points caused a reduction in statistical power of K function analysis. The same situation has been observed when calculating the K function for plasmacytosis with a similar number of observations [Reference Themudo, Østergaar and Ersbøll36].
The risk of Campylobacter colonization is highest during the summer. Taking extra precautions during this season by cleaning and drying of floor between flocks, use of fly repellent or fly nets, more restriction on the movement of staff into the chicken houses may contribute to a reduction in prevalence. These precautions and others could be particularly relevant to implement as part of an intervention against Campylobacter colonization on farms located in the cluster areas. Moreover, farmers could be advised to consider implementation of biosecurity measures with regard to the presence of colonized farms within the estimated range of influence. A future goal is to provide better tailored recommendations for groups of farms (common quality of housing, common attitude towards biosecurity and common external risk factors) about biosecurity measures such as those mentioned earlier. A greater success in implementation of biosecurity measures would then hopefully follow.
An extensive study aimed at identification of other farm-level risk factors that could lead to recommendations as on farm control measures is planned. Areas with high risk were identified and this knowledge can be used to improve the design of surveillance programmes (e.g. risk-based surveillance). Moreover, our estimates of the range of spatial dependence in the data added new knowledge on the transmission pattern of bacteria during summer. Estimated range of influence has strengthened the hypothesis about the role of flies and birds in transmission.
ACKNOWLEDGEMENTS
This work was financed by the Danish Agency for Science, Technology and Innovation, project number 09-065034/FTP.
DECLARATION OF INTEREST
None.





