



Introduction
Kawasaki disease is a paediatric systemic vasculitis that can lead to severe complications, such as coronary artery abnormalities and myocarditis. Reference Aggarwal, Pilania and Sharma1 It is the most common cause of acquired heart disease in children, predominantly affecting boys and nearly 85% of those under 5 years old. Reference Aggarwal, Pilania and Sharma1,Reference Burns2 In 2021–2022, China established expert guidelines for using intravenous immunoglobulin, acetylsalicylic acid, and glucocorticosteroids in paediatric Kawasaki disease, providing crucial direction for its diagnosis and treatment. 3 However, treatment is often delayed due to challenges in identifying symptoms, which lacks sufficient principal clinical signs. Reference Chen, Chu and Chang4 This highlights the urgent need for an early diagnostic test to enable the prompt initiation of immunoglobulin therapy, as coronary artery damage begins early in the disease course. Reference Burns2,Reference Banerjee, Pal, Chakravarti, Basu and Ahmed5 Abnormal urinary findings, such as sterile pyuria, proteinuria, and microscopic haematuria, are often seen in the acute phase of Kawasaki disease. Reference Kawamura, Takeshita and Kanai6 Recent research has highlighted the potential of urinary biomarkers for diagnosing Kawasaki disease, with studies revealing novel candidate molecular markers that exhibit excellent diagnostic performance. Reference Kentsis, Shulman and Ahmed7
Among these biomarkers, exosomes, nano-sized extracellular vesicles (30–200 nm) enclosed by a lipid bilayer membrane, are released by all cell types and transport biologically active substances such as nucleic acids, proteins, lipids, and metabolites. Reference Ferro, Saccu and Mattivi8,Reference Tsai, Kuo and Hung9 Urine has recently been identified as a source of exosomes that selectively transport biomolecules, including proteins, lipids, and microRNAs, Reference Cocci, Bondi and Santangelo10 which are small non-coding RNAs playing critical regulatory roles in physiological processes and potentially serving as non-invasive biomarkers for Kawasaki disease diagnosis. Reference Weng, Cheng, Chien, Ger, Huang and Tsai11,Reference Yun, Lee, Yun, Lim and Choi12 Building on these findings, analysing microRNAs in urinary exosomes presents a promising method for early and non-invasive Kawasaki disease diagnosis, potentially improving timely treatment and management of the disease.
The microRNA-200 family consists of two gene clusters: microRNA-200a/microRNA-200b/microRNA-429 on chromosome 1p36.33 and microRNA-200c/microRNA-141 on chromosome 12p13.31. Reference Hsieh, Huang and Yu13 This family is involved in various paediatric conditions. Reference Smith, Zuckerman, Kandanearatchi, Thompson and Davenport14–Reference Liu, Cai, Wang, Zou and Zhao16 Moreover, the microRNA-200 family has been implicated in various inflammatory and cardiovascular diseases. Reference Zhang and Wang17,Reference Reddy, Jin and Villeneuve18 This family is particularly relevant to Kawasaki disease due to its known roles in regulating inflammation and endothelial dysfunction, which are critical factors in the pathogenesis of vascular diseases.
Given this background, our study aimed to investigate the diagnostic potential of urinary exosomal microRNA-200 family members in Kawasaki disease patients and their association with clinical parameters. By employing high-throughput sequencing and quantitative real-time polymerase chain reaction, we analysed the expression profiles of these microRNAs in Kawasaki disease patients and healthy controls. We further assessed their diagnostic accuracy using receiver operating characteristic analysis and explored correlations with inflammatory markers and platelet counts. Our findings suggest that specific microRNA-200 family members are significantly upregulated in Kawasaki disease patients and closely associated with key clinical parameters, underscoring their potential as non-invasive biomarkers for early Kawasaki disease diagnosis and management.
Methods and materials
Subjects
A total of 15 paediatric Kawasaki disease patients selected from January to December 2022, whose clinical presentations met the diagnostic criteria for Kawasaki disease as outlined in The expert consensus on diagnosis and acute-phase treatment of Kawasaki disease in 2022 19 were included if disease duration was less than 10 days and without immunoglobulin therapy. A control group of 15 healthy children, who underwent routine physical examinations during the same period, was also recruited. The control group consisted of healthy children without symptoms and/or signs of infection or any underlying disease/disorders. Exclusion criteria for both groups included the presence of underlying kidney disease or a family history of hereditary kidney disorders, concomitant acute or chronic diseases, ongoing medication treatments, or non-cooperation from the child’s family, which could impede sample collection or cause contamination during the process. The study was approved by the Ethics Committee of our hospital, and informed consent was obtained from the parents or guardians of all participants.
Sample collection
Midstream morning urine (20 mL) was collected from all study participants. Each sample was divided into two 10 mL aliquots. One aliquot was collected for high-throughput sequencing. The second aliquot was used to validate the findings by quantifying the urinary exosomal expression of microRNA-200 family members using quantitative real-time polymerase chain reaction. Samples were kept at 4°C and processed within 4–6 h after the collection time.
Isolation of urinary exosomes
Total exosome isolation (Catalog Number: 4484452, Thermo Fisher Scientific Inc., China) was used for the isolation process. Urine samples were removed from storage and placed on ice. The samples were centrifuged at 2000 × g for 30 minutes at 4°C to remove cells and debris. The supernatant containing the clarified urine was carefully transferred to a new tube without disturbing the pellet. An equal volume of total exosome isolation reagent (from urine) was added to the clarified urine, and the mixture was homogenised by inverting the tube. The sample was incubated at room temperature for 1 hour. After incubation, the sample was centrifuged at 10,000 × g for 1 hour at 4°C. The supernatant was carefully aspirated and discarded without disturbing the exosome-containing pellet at the bottom of the tube. Any residual supernatant was removed by careful aspiration. The pellet was resuspended in 1X PBS by vortexing to ensure the exosomes were fully resuspended.
Isolation of RNA from exosomes
RNA from exosomes was isolated using the total exosome RNA and Protein Isolation Kit (Catalog Number 4478545, Thermo Fisher Scientific Inc., China). Exosome samples were adjusted to 200 µL with 1X PBS, then mixed with an equal volume of 2X denaturing solution and incubated on ice for 5 minutes. Acid-phenol was added, vortexed, and centrifuged at ≥ 10,000 × g for 5 minutes. The aqueous phase was mixed with 100% ethanol and passed through a filter cartridge. After washing with miRNA Wash Solution, RNA was eluted with 50 µL of preheated nuclease-free water and stored at ≤ −20°C. Purity and concentration of RNA were assessed using the Agilent 2100 Bioanalyzer with the RNA 6000 p Kit to ensure that samples met the quality requirements for further analysis.
MicroRNA library preparation and illumina SBS high-throughput sequencing
Small RNA libraries were prepared using the TruSeq Small RNA Sample Preparation kit (Illumina, San Diego, CA) according to the manufacturer’s protocols. After ligating RNA with adapters, RT-PCR was performed to create cDNA libraries. The amplified cDNA fragments were size selected and pooled for sequencing on the Illumina HiSeq2500 platform. Quality control was performed using FastQC to ensure high-quality sequencing reads (score > 30), followed by trimming of adapter sequences and filtering for appropriate read sizes (16–35 nt). Low-abundance reads (<10 reads) were discarded, and reads were aligned to the human genome (hg18) and miRBase v16 for miRNA identification. Reference Hackenberg, Sturm, Langenberger, Falcon-Perez and Aransay20
Quantitative real-time polymerase chain reaction
RNA from exosomes was isolated using the total exosome RNA and protein isolation kit (Catalog Number 4478545, Thermo Fisher Scientific Inc., China). Complementary DNA was synthesised from 5 ng of total RNA using the miRCURY Locked Nucleic Acid Reverse Transcription kit (QIAGEN, Hilden, Germany, Cat. No. 339340) in 20 μL reaction volumes, with the inclusion of a 10 × miRCURY Reverse Transcription Enzyme Mix and UniSp6 RNA spike-in (QIAGEN). Then, quantitative real-time polymerase chain reaction analysis was performed on a CFX384 C1000 Thermal Cycler (Bio-Rad, Hercules, CA, USA) using a 2× miRCURY SYBR Green Master Mix (QIAGEN). The primer sequences used in this study are listed in Table 1. Cycle threshold values were obtained, and the relative RNA expression levels were calculated using the comparative 2-ΔΔCt method.
Table 1. Sequences of the microRNA primers used in this study

Statistical analyses
The statistical analyses were conducted using GraphPad Prism (GraphPad Software Inc., San Diego, CA, USA), MedCalc software (MedCalc Software bv, Ostend, Belgium), or SPSS (IBM Corp., Armonk, NY, USA). The Shapiro–Wilk test was employed to assess normal distribution, with normally distributed data compared using an unpaired t-test (mean ± standard deviation) and non-normally distributed data using the Mann–Whitney test (median, interquartile range). Spearman correlation analysis evaluated correlations between urinary exosomal microRNA-200 family members and clinical parameters in Kawasaki disease patients. The diagnostic potential of these microRNAs for Kawasaki disease was assessed using receiver operating characteristic analysis in MedCalc, with cut-off values determined by the maximum Youden index. Logistic regression models in SPSS examined the relationship between Kawasaki disease occurrence and microRNA-200a-3p/5p, microRNA-200b-3p/5p, and microRNA-429, while the combined predictive capacity was evaluated by analysing the area under the receiver operating characteristic curve followed by DeLong test for comparisons. Reference Demler, Pencina and D’Agostino21 Statistical significance was set at p < 0.05 for all analyses.
Results
Baseline characteristics of enrolled participants
Fifteen Kawasaki disease patients and 15 healthy control subjects were included in the study (Table 2). Both groups had an equal sex distribution (8 males and 7 females), with no significant difference in mean age (3.68 ± 1.65 years for controls vs. 3.31 ± 1.18 years for Kawasaki disease, p = 0.482) or body mass index (15.83 ± 1.35 kg/m2 for controls vs. 16.06 ± 1.06 kg/m2 for Kawasaki disease, p = 0.612). Significant differences were observed in blood parameters, where Kawasaki disease patients had lower haemoglobin (p = 0.002), higher white blood cell counts (p < 0.001), higher platelet counts (p = 0.023), higher erythrocyte sedimentation rate (p < 0.001), higher C-reactive protein levels (p < 0.001), higher aspartate aminotransferase (p < 0.001), and higher alanine aminotransferase (p < 0.001) compared to controls.
Table 2. Comparisons of baseline characteristics between 15 patients with Kawasaki disease (KD) and 15 healthy control subjects

BMI = body mass index; Hb = haemoglobin; WBC = white blood cell count; ESR = erythrocyte sedimentation rate; CRP = C-reactive protein; AST = aspartate aminotransferase; ALT = alanine aminotransferase.
Urinary exosomal expression of microRNA-200 family members in Kawasaki disease patients
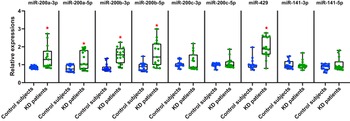
Heatmap analysis of 74 urinary exosomal microRNAs distinguished between control samples and Kawasaki disease urine samples (p < 0.10, Figure 1A). We observed a total of 43 differentially expressed urinary exosomal microRNAs in Kawasaki disease patients, all of which were upregulated compared to controls [log2 fold change > 0, p < 0.05]. Among these, 20 microRNAs (microRNA-215-5p, microRNA-151b, microRNA-455-5p, microRNA-99a-5p, microRNA-107, microRNA-193b-3p, microRNA-532-3p, microRNA-200b-5p, let-7f-2-3p, microRNA-140-3p, microRNA-148b-3p, microRNA-125b-5p, microRNA-501-5p, microRNA-345-5p, microRNA-1246, microRNA-190a-3p, microRNA-135a-5p, microRNA-365b-3p, and microRNA-200a-5p, let-7a-3p) were found to be significantly upregulated (log2 fold change > 2, p < 0.05). Notably, the microRNA-200 family, including hsa-microRNA-429 (p = 0.004), hsa-microRNA-200b-3p (p = 0.007), hsa-microRNA-200b-5p (p = 0.021), hsa-microRNA-141-3p (p = 0.044), hsa-microRNA-200a-5p (p = 0.044), hsa-microRNA-200a-3p (p = 0.044), and hsa-microRNA-200c-3p (p = 0.046), were all upregulated. These results are clearly illustrated in the volcano plot, showing the distinct separation and significant upregulation of the microRNA-200 family members (Figure 1B). Quantitative real-time polymerase chain reaction analysis confirmed the upregulation of microRNA-200a-3p (1.64-fold, p = 0.001), microRNA-200a-5p (1.58-fold, p = 0.006), microRNA-200b-3p (1.81-fold, p < 0.001), microRNA-200b-5p (1.82-fold, p = 0.001), and microRNA-429 (1.88-fold, p < 0.001) in Kawasaki disease patients compared to controls (Figure 2). However, microRNA-200c-3p/5p and microRNA-141-3p/5p did not show significant differences between the two groups (all p > 0.05).

Figure 1. Dysregulation of urinary exosomal microRNAs in Kawasaki disease patients. A: Heatmap analysis of 74 urinary exosomal microRNAs (miRNAs) distinguishing control samples from Kawasaki disease (KD) urine samples (p < 0.10). B: 43 differentially expressed urinary exosomal miRNAs in KD patients, all of which were upregulated compared to controls (log2 fold change [log2FC] > 0, p < 0.05).

Figure 2. Urinary exosomal expression of microRNA (miR)-200 family members in Kawasaki disease (KD) patients. Comparison of the urinary exosomal expression levels of miR-200 family members between control samples (n = 15) and KD urine samples (n = 15) using quantitative reverse transcription polymerase chain reaction (qRT-PCR). *Indicates a statistically significant difference compared to control subjects (p < 0.05).
Diagnostic potential of urinary exosomal microRNA-200 family members for Kawasaki Disease
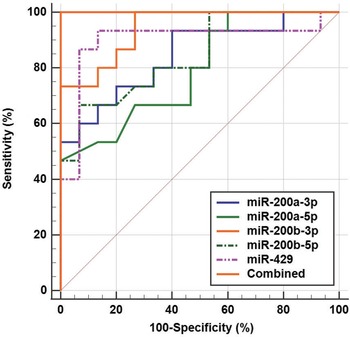
The diagnostic potential of urinary exosomal microRNA-200 family members for Kawasaki disease was evaluated using receiver operating characteristic analysis (Table 3). MicroRNA-200c-3p and microRNA-200c-5p, with area under the curves of 0.516 and 0.507, and microRNA-141-3p and microRNA-141-5p, with area under the curves of 0.520 and 0.533, respectively, all showed limited and statistically insignificant diagnostic potential (all p > 0.05). Conversely, microRNA-200b-3p exhibited the highest diagnostic accuracy with an area under the curve of 0.942 (sensitivity: 100.00%, specificity: 73.33%, p < 0.001). MicroRNA-429 had an area under the curve of 0.898 (sensitivity: 93.33%, specificity: 86.67%, p < 0.001), followed by microRNA-200a-3p with an area under the curve of 0.844 (sensitivity: 93.33%, specificity: 60.00%, p < 0.001), and microRNA-200b-5p with an area under the curve of 0.842 (sensitivity: 66.67%, specificity: 93.33%, p < 0.001). microRNA-200a-5p showed moderate diagnostic value with an area under the curve of 0.791 (sensitivity: 46.67%, specificity: 100.00%, p < 0.001). The combined analysis of microRNA-200a-3p/5p, microRNA-200b-3p/5p, and microRNA-429 yielded a perfect area under the curve of 1.000, indicating a highly accurate diagnostic test with both 100% sensitivity and specificity (Figure 3). Pairwise comparisons showed significant improvements in diagnostic accuracy when these microRNAs were analysed together (all p < 0.05).

Figure 3. Diagnostic potential of urinary exosomal microRNA (miR)-200a-3p/5p, miR-200b-3p/5p, and miR-429 for Kawasaki disease (KD) using receiver operating characteristic (ROC) analysis. The combined analysis of these microRNAs yielded a perfect area under the curve (AUC) of 1.000, indicating a highly accurate diagnostic test with both 100% sensitivity and specificity.
Table 3. Diagnostic value of urinary exosomal miR-200 family members for Kawasaki disease (KD) using Receiver Operating Characteristic (ROC) analysis

Correlation of urinary exosomal microRNA-200 family members with inflammatory markers in Kawasaki disease
As demonstrated in Table 4, Spearman correlation analysis demonstrated strong positive correlations between microRNA-200a-3p and white blood cells (r = 0.629, p = 0.014), platelet counts (r = 0.590, p = 0.023), and C-reactive protein levels (r = 0.609, p = 0.018). MicroRNA-200a-5p also correlated with C-reactive protein (r = 0.735, p = 0.003), platelet counts (r = 0.558, p = 0.033), and white blood cells (r = 0.586, p = 0.024). Both microRNA-200b-3p and microRNA-429 were positively correlated with white blood cells, platelet counts, erythrocyte sedimentation rate, and C-reactive protein (all p < 0.05). Additionally, microRNA-200b-5p displayed moderate positive correlations with platelet counts (r = 0.595, p = 0.021) and erythrocyte sedimentation rate (r = 0.521, p = 0.049). Conversely, microRNA-200c-3p, microRNA-200c-5p, microRNA-141-3p, and microRNA-141-5p showed no significant correlations with any of the clinical parameters (all p > 0.05). Additionally, none of the urinary exosomal microRNA-200 family members displayed correlations with aspartate aminotransferase or alanine aminotransferase levels (all p > 0.05).
Table 4. Correlation analysis of urinary exosomal miR-200 family members with inflammatory markers in Kawasaki disease (KD) patients

BMI = body mass index; Hb = haemoglobin; WBC = white blood cell count; ESR = erythrocyte sedimentation rate; CRP = C-reactive protein; AST = aspartate aminotransferase; ALT = alanine aminotransferase.
Discussion
The present study investigated the diagnostic potential of urinary exosomal microRNA-200 family members in Kawasaki disease patients. High-throughput sequencing and quantitative real-time polymerase chain reaction analyses revealed a significant upregulation of several microRNA-200 family members, particularly microRNA-200a-3p, microRNA-200a-5p, microRNA-200b-3p, microRNA-200b-5p, and microRNA-429, suggesting their involvement in the inflammatory processes characteristics of Kawasaki disease.
Kawasaki disease is a vasculitis of unknown aetiology, and if left untreated, there is a high risk of developing coronary artery aneurysms. Reference Mossberg, Mohammad, Kahn, Segelmark and Kahn22 The inflammatory process in Kawasaki disease is complex, with cytokines such as interleukin-6, interleukin-8, tumour necrosis factor-alpha, interferon-gamma, and C-reactive protein closely associated with disease progression and aneurysm formation. Reference Sarejloo, Shahri and Azami23,Reference Kim, Shimizu and He24 High neutrophil levels, in particular, promote vascular infiltration, leading to necrosis and aneurysm formation. Reference Sarejloo, Shahri and Azami23 In this context, our Spearman correlation analysis revealed strong associations between microRNA-200a-3p, microRNA-200a-5p, microRNA-200b-3p, microRNA-200b-5p, microRNA-429, and key clinical parameters such as white blood cells, platelet count, erythrocyte sedimentation rate, and C-reactive protein. This indicates that these microRNAs may actively modulate the inflammatory response in Kawasaki disease. Similar findings have been reported in other inflammatory diseases; for example, microRNA-200b has been positively correlated with platelet count in patients with systemic lupus erythematosus. Reference Wang, Tam and Li25 These associations support the idea that microRNA-200 family members are more closely linked to inflammatory and haematological parameters rather than liver function, as evidenced by the absence of significant correlations with aspartate aminotransferase or alanine aminotransferase levels.
Endothelial injury and dysfunction are key triggers for the inflammatory reactions that drive the development of atherosclerosis. Reference Seki and Minami26 Similarly, Kawasaki disease patients may experience endothelial dysfunction as a vascular sequela, suggesting a potential early development of atherosclerosis in these individuals. Reference Fukazawa and Ogawa27 Members of the microRNA-200 family, particularly microRNA-200c, are known regulators of apoptosis, inflammation, and endothelial dysfunction in atherosclerosis. Reference Letonja and Petrovic28 Specifically, microRNA-200c is induced by reactive oxygen species, promotes inflammation, and reduces antioxidant capacity by decreasing the transcription of manganese superoxide dismutase. Reference Magenta, D’Agostino and Sileno29,Reference Magenta, Sileno and D’Agostino30 In atherosclerosis, microRNA-200c is upregulated in unstable plaques and correlates with instability markers such as interleukin-6 and cyclooxygenase-2. Reference Magenta, Sileno and D’Agostino30 This role in endothelial dysfunction and inflammation may also contribute to vascular injury in Kawasaki disease, potentially linking microRNA-200c to adverse outcomes like aneurysm formation.
Zhang F et al. found that microRNA-200b-3p was upregulated in epicardial adipose tissue during atherosclerosis, promoting endothelial cell apoptosis by targeting histone deacetylase 4 under oxidative stress conditions, which contributed to vascular inflammation and damage. Reference Zhang, Cheng, Du, Zhang and Zhang31 Furthermore, microRNA-429 has been shown to promote apoptosis of endothelial cells in atherosclerosis by downregulating the antiapoptotic protein B-cell lymphoma 2. Reference Zhang, Tian, Wang, Jing, Zhou and Chen32 Similarly, microRNA-141 has been implicated in cardiovascular conditions. Reference Zhang, Kong and Ma33 Li Y et al. found that microRNA-141-5p was downregulated in atherosclerosis and inhibited vascular smooth muscle cell inflammation and proliferation by targeting the high mobility group box 1/nuclear factor-kappa B pathway. Reference Li, Li, Chen, Yang and Hao34 These findings suggest that the upregulated microRNA-200 family members in Kawasaki disease may contribute to the disease’s pathogenesis through mechanisms similar to those observed in other vascular inflammatory diseases.
However, in our study, microRNA-200c-3p/5p and microRNA-141-3p/5p did not show significant differences between the Kawasaki disease and control groups and had limited diagnostic potential. This is in contrast to previous findings. Yun et al. reported significantly upregulated serum microRNA-200c levels in children with Kawasaki disease, highlighting its role in the inflammatory response. Reference Yun, Lee, Yun, Lim and Choi12 Similarly, Zhang et al. found that serum microRNA-200c levels were elevated in Kawasaki disease patients who did not respond to intravenous immunoglobulin therapy, suggesting its potential as a diagnostic biomarker and therapeutic target. Reference Zhang, Wang, Zeng, Hu and Zou35 Conversely, Huang BQ et al. found that microRNA-200c-3p was downregulated in plasma exosomes from patients with Takayasu’s arteritis, suggesting its involvement in vascular physiology and autoimmunity. Reference Huang, Li, Tian and Zeng36 These discrepancies may be explained by variations in sample types (serum vs. exosomes), disease stages, or distinct underlying mechanisms unique to each condition. For microRNA-141, prior studies have demonstrated its involvement in cardiovascular diseases, which share common inflammatory and vascular characteristics with Kawasaki disease. Zhao et al. reported that proprotein convertase subtilisin/kexin type 9 inhibitors enhance plaque stability by targeting microRNA-141-5p, influencing cardiovascular smooth muscle cell proliferation and migration. Reference Zhao, Liu, Zhang and Zhao37 Additionally, microRNA-141-3p has been linked to antioxidant pathways, as hydroxyurea treatment was predicted to activate its expression by inducing antioxidant genes and reducing oxidative stress via the Nrf2 signalling pathway in endothelial cells. Reference Santana, Pitanga and de Santana38 Although we did not find significant differences in urinary exosomal microRNA-141-3p and microRNA-141-5p levels in Kawasaki disease patients, microRNA-141’s established role in vascular inflammation and smooth muscle regulation in cardiovascular diseases suggests that it may play a complex, albeit subtle, role in Kawasaki disease pathogenesis.
Despite these promising findings, this study has several limitations. First, the relatively small sample size may limit the generalisability of the results. With only 15 Kawasaki disease patients and 15 controls, the study may lack sufficient power to detect smaller or subtler effects, potentially increasing the risk of type II errors. Larger cohort studies are needed to validate these findings and confirm the diagnostic potential of urinary exosomal microRNA-200 family members in broader populations. Second, the lack of a febrile control group means that we cannot fully ascertain whether the observed miRNA changes are specific to Kawasaki disease or reflective of general febrile conditions. Febrile illnesses often involve systemic inflammation, which could potentially influence miRNA expression. Including patients with other febrile diseases in future studies would help clarify whether the identified microRNAs are uniquely associated with Kawasaki disease or represent common inflammatory responses. This would strengthen the specificity of these biomarkers for Kawasaki disease diagnosis. Finally, while we demonstrate correlations between microRNA-200 family members and inflammatory markers, the underlying mechanisms linking these microRNAs to Kawasaki disease pathogenesis remain unclear. Longitudinal studies are required to assess the role of these microRNAs in disease progression and response to therapy, providing insights into their potential as prognostic markers.
Conclusion
In conclusion, urinary exosomal microRNA-200 family members, particularly microRNA-200a-3p, microRNA-200a-5p, microRNA-200b-3p, microRNA-200b-5p, and microRNA-429, exhibit strong potential as non-invasive biomarkers for Kawasaki disease diagnosis. Their significant association with inflammatory markers and platelet counts suggests a role in the disease’s pathogenesis, offering insights into potential targets for therapeutic intervention. The integration of these microRNAs into clinical practice could enhance the early diagnosis and management of Kawasaki disease, ultimately improving patient outcomes.
Availability of data and materials
The datasets generated and/or analysed during the current study are available from the corresponding author upon reasonable request.
Authors’ contributions
Yu-E Wang, Da-Qing Sun, and Hui-Zhong Hu contributed to the conception and design of the study. Yu-E Wang performed the experiments and data analysis. Da-Qing Sun contributed to the collection and interpretation of data. Hui-Zhong Hu supervised the project and wrote the manuscript. All authors reviewed and approved the final manuscript.
Financial support
None.
Competing interests
The authors declare that they have no competing interests.
Ethical standard
This study was conducted in accordance with the Declaration of Helsinki and was approved by the Ethics Committee of Xi’ an Third Hospital, Xi’ an, Shanxi, China. Written informed consent was obtained from the parents or guardians of all paediatric participants involved in the study.
Consent for publication
Not applicable.






 Open access
Open access