


Introduction
Forests host an enormous diversity of defoliating insects, most of which persist at low population levels, nibbling at foliage but rarely causing conspicuous damage. A comparatively small percentage of species are prone to occasional outbreaks, attaining population levels that can strip single trees or entire forests of foliage. Although taxonomically rare, outbreak defoliators have for more than 60 years served as the model system of choice for developing theories of animal population dynamics (Cappuccino Reference Cappuccino1995). This bias may reflect several factors. Outbreak defoliators tend to be conspicuous and easy to sample, attack economically and sociopolitically important plants, and undergo immense, but not unbounded, fluctuations in population density. Moreover, there are numerous long-term time series of population levels for most major outbreak species, many of which offer tantalising hints of cyclic dynamics at local, regional, or continental scales. Much of what we know about animal population dynamics can be traced to pioneering studies involving outbreak defoliators.
Our understanding of defoliator population dynamics has progressed significantly during the past century, aided by technological and analytical advances and guided by vibrant debate (reviewed by Turchin Reference Turchin1995; Liebhold and Kamata Reference Liebhold and Kamata2000). Debates have centred largely on fundamental questions concerning the underlying nature of outbreak population behaviour. For instance, why are some defoliators prone to outbreak while the majority are not? Do outbreaks occur with a regular periodicity and are they spatially synchronised? What are the main ecological processes driving outbreak cycles? Or, as Royama (Reference Royama1992) put it simply, “Why do animal populations fluctuate as they do?” Our review summarises the efforts of researchers during the past century to address these questions for tree-defoliating insects in Canada. We begin with a basic synopsis of the outbreak defoliators reported in Canadian forests during the past century. Next, we examine studies that have addressed the above questions, focussing on broad historical trends, the frequency of various research approaches used, and some general patterns in the conclusions drawn from these studies. Finally, we feature selected case studies to highlight the history and scope of population research in Canada and to illustrate some of the key insights provided from the different approaches.
Scope and data sources
Our summary of outbreak defoliators was derived primarily from annual Forest Insect and Disease Survey (FIDS) reports (1936–1995). Broadly defined, a “defoliator” might include any insect species that consumes the leafy, photosynthetic tissue of its host plant. However, for the purposes of our review, we focus on defoliators that feed externally on leaves and exclude those that feed from within leaves (e.g., leaf miners and gall formers). This is in part because few miners or gallers are considered to be major outbreak pests in Canada, but is also because the ecological processes governing population fluctuations of these groups tend to differ from those of external feeders (Price Reference Price1997). It should be noted also that our reference to so-called “outbreak species” encompasses a significant breadth of population behaviours ranging from brief, local bouts of intense defoliation to multi-year, landscape-level epidemics. Not all species included would fall under other more rigorous definitions of “outbreak species” (e.g., Hunter Reference Hunter1995); however, for completeness, we have opted to include all species reported to have caused “severe” defoliation at any scale in the FIDS reports.
Details on population studies were gathered from available reference sources, including Google Scholar (dates unknown), Biological Abstracts (1989–2014), and various books and review articles that have explored the population dynamics of outbreak insects (e.g., Cappuccino and Price Reference Cappuccino and Price1995; Barbosa et al. Reference Barbosa, Letourneau and Agrawal2012). As key words, we used “defoliator” combined with each of “time series”, “chronology”, “periodicity”, “spatial synchrony”, “life table”, “key factor”, or “density dependent”, and selected articles (and references therein) that involved research in Canadian forests, regardless of the nationalities of author or journal. Although this search was not exhaustive, it should provide a fair representation of Canadian population ecology research.
Outbreak defoliators in Canada
At least 106 species of defoliators have been reported to outbreak in Canadian forests during the past century, all from the orders Lepidoptera, Hymenoptera (sawflies), and Coleoptera (beetles) (from most to fewest species; Fig. 1A, Table 1). However, if we were to restrict this list to include only those that have significant long-term economic or ecological impacts on Canadian forests, less than half a dozen species would remain, with most being Lepidoptera (e.g., tent caterpillar (Malacosoma disstria Hübner; Lasiocampidae), spruce budworm (Choristoneura fumiferana Clemens; Tortricidae), western spruce budworm (Choristoneura freemani Rozowski; Tortricidae), hemlock looper (Lambdina fiscellaria Guenée; Geometridae)) or Hymenoptera (larch sawfly (Pristiphora erichisonii Hartig; Tenthredinidae)). Most outbreaking species in Canada are native (86%), although it is notable that nearly 25% of all outbreaking sawfly species are exotics from Europe (Fig. 1A). Whereas most lepidopteran (74%) and all beetle outbreaks occur on deciduous hosts, sawfly outbreaks occur mainly on conifers (70%, including both evergreen and deciduous conifers) (Fig. 1B). Most outbreak defoliator species are univoltine (i.e., have one generation per year), except for beetles, which generally have short generation times and are split equally between univoltine and bivoltine species (Fig. 1C).
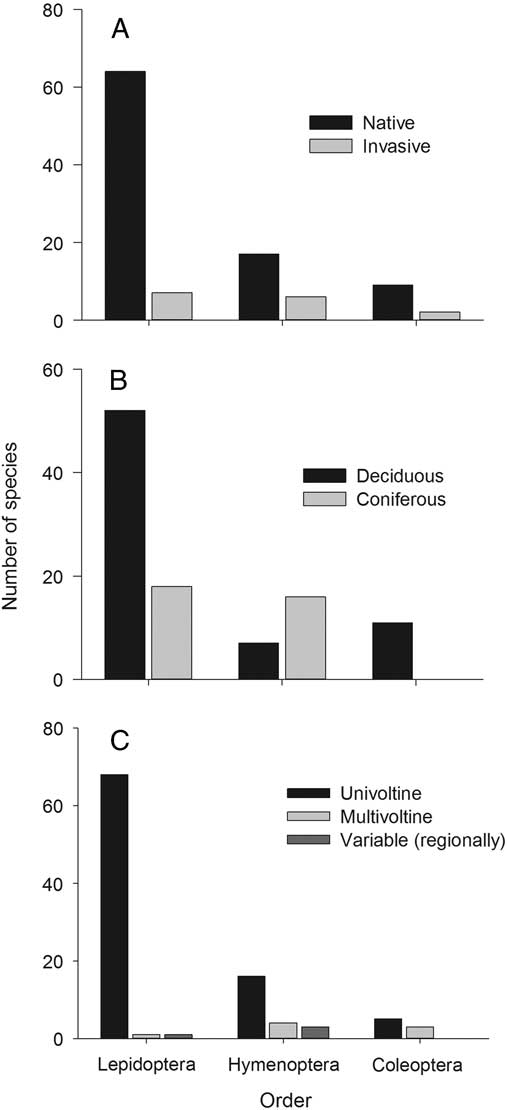
Fig. 1 The number of defoliator species in the orders Lepidoptera, Hymenoptera, and Coleoptera reported to have caused severe defoliation in Canada on at least one occasion between 1936 and 2014. Defoliators are categorised with respect to (A) native/invasive status, (B) host-type preference, and (C) voltinism.
Population studies
Why are some species prone to outbreak?
Comparative studies
It remains a great mystery why most insect defoliators persist at relatively low, stable population levels, but a small percentage, perhaps as little as 1–3%, are prone to outbreaks (Faeth Reference Faeth1987; Hunter Reference Hunter1991, Reference Hunter1995). Comparative studies aim to shed light on this mystery by identifying ecological traits associated with the differing outbreak tendency between closely related species occupying similar habitats (Hunter Reference Hunter1991; Koricheva et al. Reference Koricheva, Klapwijk and Bjökman2012). One of the major challenges of this approach is that it requires substantial knowledge of basic life-history traits for both outbreak and non-outbreak species (e.g., size, fecundity, egg, and larval distribution, etc.). Although ample information is often available for outbreak species, rare or cryptic species are largely unstudied outside of taxonomy, and even basic traits remain unknown. A few researchers, however, have been able to cobble together enough information for certain groups to provide interesting insights on patterns of traits seemingly common among outbreaks species (reviewed in Koricheva et al. Reference Koricheva, Klapwijk and Bjökman2012).
In one such Canadian study of North American Lepidoptera on hardwoods, Hunter (Reference Hunter1991) found numerous traits common (though not universal) among outbreak species, including spring feeding, polyphagy, overwintering as eggs, gregarious larvae, clustered eggs, high fecundity, large size, and strong defenses against natural enemies. In general, outbreak defoliators could be further delineated into one of two groups: (1) spring feeders that remain as eggs for most of the year and (2) summer feeders that feed gregariously (Hunter Reference Hunter1991). Subsequent analyses including European species detected similar trends and ruled out possible confounding effects of phylogeny (Hunter Reference Hunter1995). Although not Canadian in origin, similar studies for sawflies have noted that many outbreak-prone species have some of the same ecological traits as outbreaking Lepidoptera, in particular clustered egg lay and larval feeding (Haack and Mattson Reference Haack and Mattson1993; Larsson et al. Reference Larsson, Bjökman and Kidd1993; Koricheva et al. Reference Koricheva, Klapwijk and Bjökman2012), as well as an unusually high outbreak tendency among species that prefer coniferous over deciduous hosts (Price et al. Reference Price, Roininen and Ohgushi2005). Although comparative analyses are not definitive in explaining why certain species have outbreak population dynamics, they do underscore potentially significant evolutionary and ecological trends that warrant further investigation (e.g., the adaptive significance of clustered egg lay and gregarious larval feeding for outbreak species, reviewed in Cappuccino et al. Reference Cappuccino, Damman and Dubuc1995).
Are outbreaks periodic and spatially synchronised?
Time-series studies
One of the major goals in the study of insect population dynamics is to understand the spatiotemporal nature of population fluctuations. In particular, population behaviours such as periodicity (i.e., statistically significant regularity in population cycles) and spatial synchrony (i.e., similar density fluctuations among spatially disjunct populations) have caught the interest of many population ecologists due at least in part to their seeming ubiquity among most major outbreak defoliator species (Liebhold and Kamata Reference Liebhold and Kamata2000). However, elucidating these spatial and temporal trends with any statistical rigor can be challenging, as it requires relatively long-term data sets on insect density, ideally covering multiple outbreaks and the full outbreak range. Most of the major outbreak defoliators outbreak at 10–35-year intervals and thus require decades or even centuries of time-series data for there to be any conclusive analysis of outbreak trends.
In Canada, there have been at least 50 time-series studies for 12 defoliator species examining (at least peripherally) the periodicity and/or spatial synchrony of outbreaks (Table 2). The frequency of time-series studies has increased during the past 60 years (Fig. 2), with almost half of all studies focussing on spruce budworm. These time series have been built from various proxies for population density (Fig. 3). Anecdotal reports from historical documents formed the basis of some of the earliest published time series in Canada, which were for fall webworm and tent caterpillar (Baird Reference Baird1917; Sippell Reference Sippell1962). However, time series constructed from the tree rings of ancient timbers are by far the most common source of long-term time series, taking advantage of the tendency of trees to suffer diminished radial growth during periods of intense herbivory (Fig. 3). A smaller number of studies have used annual estimates of defoliation from aerial or ground surveys (Fig. 3). Although defoliation time series are typically shorter than those from tree rings (Fig. 3), they tend to encompass relatively large, contiguous areas, making them highly amenable to spatial statistical analyses (Table 2). Direct measures of defoliator populations (e.g., eggs or tents) clearly provide the most precise estimates of population density but are laborious to collect, short in duration, and perhaps because of this the least common source of time-series data (Fig. 3). At least one study has used macrofossils (ancient frass) to detect spruce budworm outbreaks nearly 7000 years in the past, although trends were too crude to provide much information on population behaviour (Simard et al. Reference Simard, Morin and Lavoie2006).
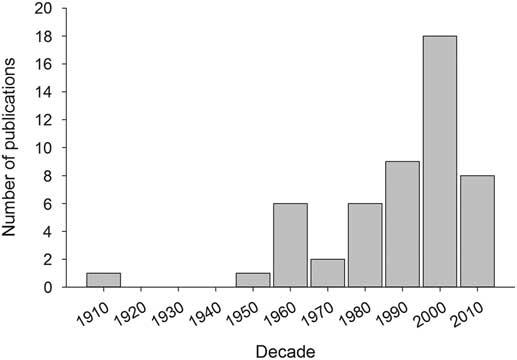
Fig. 2 The number of peer-reviewed articles per decade using time series of population density (or proxies) to assess periodicity and/or spatial synchrony in populations of outbreaking tree defoliators.
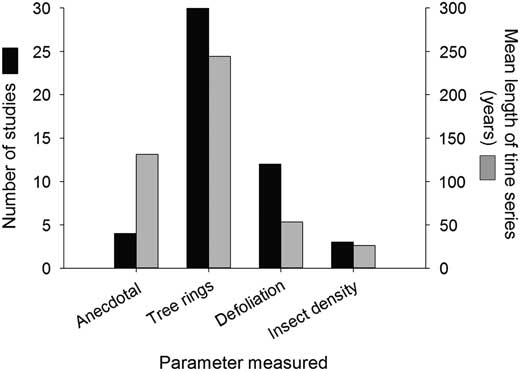
Fig. 3 The number and average length of time-series data sets assembled between 1936 and 2014 from different proxies of population density, including anecdotal reports, tree rings (i.e., dendrochronology), defoliation, and insect density.
Table 1 Summary of outbreak tree defoliators in Canada derived from the Forest Insects and Disease Surveys (1936–1995), including common and Latin names, voltinism, primary host preference(s) during outbreak, and regional distribution of reported outbreaks within Canada.
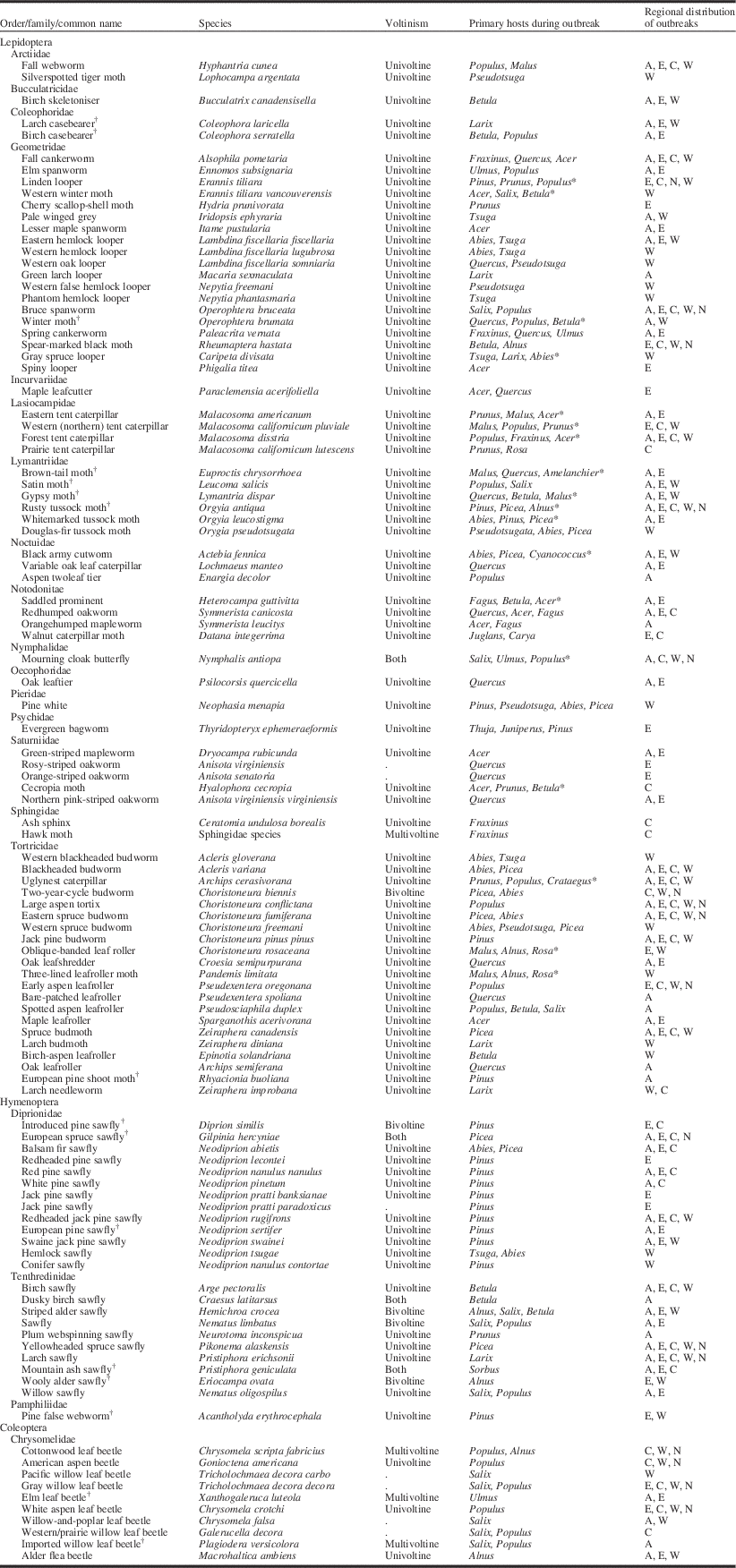
* Species reported to feed on > 3 host genera. The genera listed are the most common attacked during outbreaks.
† Invasive species.
A, Atlantic (Newfoundland and Labrador, Prince Edward Island, Nova Scotia, New Brunswick); E, eastern (Québec, Ontario), C, central (Manitoba, Saskatchewan), W, western (Alberta, British Columbia), N, northern territories (Nunavut, Northwest Territories, Yukon).
Table 2 Summary of time-series studies for outbreaking defoliators in Canadian forests during the past century assessing the periodicity and/or spatial synchrony of population cycles.
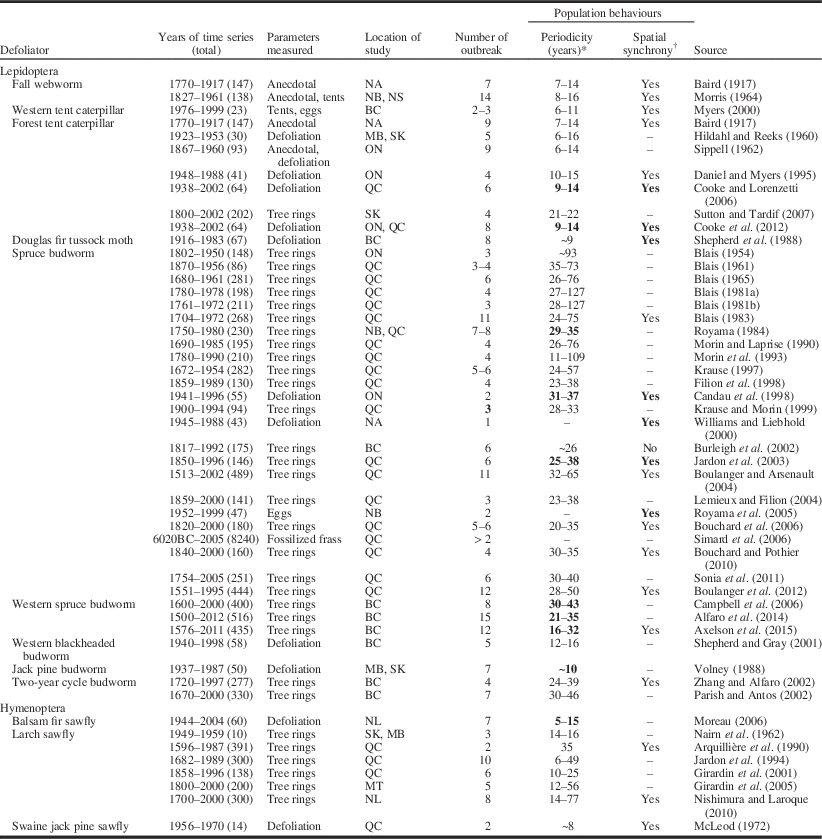
* Numbers represent the range of periods observed. Bold emphasis indicates that statistical analyses were carried out to support observed trends.
† Bold emphasis indicates that statistical analyses were carried out to support observed trends.
ON, Ontario; QC, Québec; MB, Manitoba; BC, British Columbia; SK, Saskatechewan; AB, Alberta; NS, Nova Scotia; NB, New Brunswick; NL, Newfoundland and Labrador; NA, North America; USA, United States of America.
Most time-series studies in Canada provide indications of regular, albeit imperfect periodicity for most major outbreak species. Only larch sawfly has been consistently cited as lacking any clear regularity in its cycles (Table 2). In studies with time series from multiple sites, spatial synchrony in population trends is often evident, at least in the sense that outbreak cycles in one site or region appear to fluctuate in a similar manner to those in seemingly disjoined locations (Table 2). However, the scale at which spatial synchrony occurs varies significantly among studies, sometimes occurring among fairly local sites (e.g., Moreau Reference Moreau2006) and in other instances occurring at regional (e.g., Cooke et al. Reference Cooke, MacQuarrie and Lorenzetti2012) or transcontinental scales (e.g., Williams and Liebhold Reference Williams and Liebhold2000). It is important to note that relatively few Canadian studies have carried out statistical analyses to detect either periodicity or spatial synchrony, or to examine some of the more subtle mathematical properties of the outbreak cycles (e.g., harmonic cycles, quasi-periodicity, density dependence, limit cycles, chaos, etc.). This lack of analysis, coupled with the tendency of time-series studies to focus on species that characteristically undergo periodic, spatially synchronous outbreaks, may exaggerate the ubiquity of these traits among outbreak defoliator species (Liebhold and Kamata Reference Liebhold and Kamata2000).
What are the main ecological processes that drive outbreak cycles?
Life-table studies
Life tables were originally developed as part of actuarial and demographic studies of human populations, but were later adapted to study the processes driving insect outbreaks (Morris and Miller Reference Morris and Miller1954; Harcourt Reference Harcourt1960). Whereas time series tend to focus on broad-scale or long-term population trends based on annual estimates of population density, life tables provide a more meticulous and systematic accounting across multiple life stages to identify those factors most responsible for changes in population density (Morris and Miller Reference Morris and Miller1954; Harcourt Reference Harcourt1960). Researchers in Canada have contributed significantly to the arduous task of collecting life-table data for many major outbreaking insects, as well as to the development of analytical techniques for detecting possible regulatory agents (i.e., those acting in a density-dependent manner) (Morris Reference Morris1963a; Royama Reference Royama1992).
For Canadian forest defoliators, there have been at least 12 life-table studies for seven defoliator species (Table 3). The majority of these studies have been carried out during the past 55 years (Fig. 4), and nearly half of them are for conifer-feeding budworms (Table 3). Whereas the duration of these life tables ranges between three to 35 years, all of these studies tend to focus on dynamics in only a few sites and cover only a portion of an outbreak cycle, and in most cases focus on populations in decline (Table 3). Only three life-table studies have assessed populations at low density (Table 3) and no study to date has captured the transition of a population from low to high density. In general, many factors contribute to generational mortality as populations decline; however, in most studies the primary driver of population collapse and subsequent maintenance of populations at low density was natural enemies (usually parasitoids) (Table 3). A few studies also provide evidence of host-plant feedbacks that may in some cases cause or contribute to population decline (Table 3). It should be noted that half of the studies cited have drawn their conclusions from key factor (also known as k-factor) analyses (Morris Reference Morris1959). This analytical approach, however, has been abandoned owing to various statistical and logical issues (Royama Reference Royama1996) in favor of alternative approaches (Royama Reference Royama1992). At least for spruce budworm, this shift in analytical techniques led to a new interpretation of the main factors driving outbreak cycles, from what was essentially the same data (Table 3: Morris Reference Morris1963a versus Royama Reference Royama1984).
Table 3 Summary of life-table studies for outbreaking defoliators in Canadian forests during the past century examining the potential factors driving changes in population density during outbreaks.
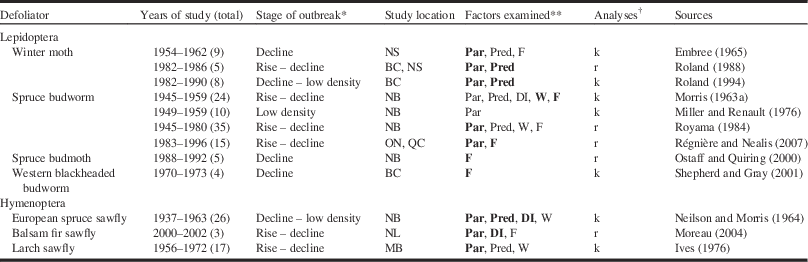
*General stages in the population cycle: rise, decline, low density.
** Bold text represents the main ecological factor(s) influencing changes in population density (based on analyses).
† Factor(s) contributing to population behaviour were detected through either key-factor or k-factor analyses (k) (Morris Reference Morris1959), or through analyses of rates of change in log population density between different life stages (r) (Royama Reference Royama1992).
ON, Ontario; QC, Québec; MB, Manitoba; BC, British Columbia; SK, Saskatechewan; NS, Nova Scotia; NB, New Brunswick; NL, Newfoundland and Labrador; Par, parasitism; Pred, predation; DI, disease; W, weather; F, food availability or quality.
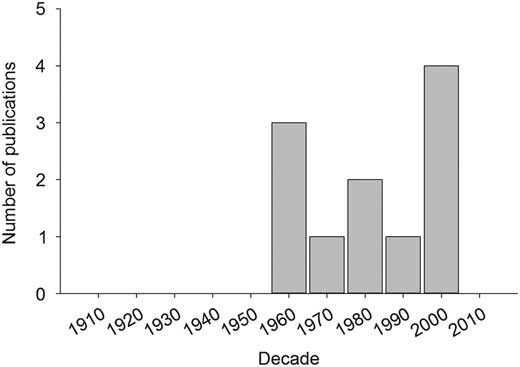
Fig. 4 The number of peer-reviewed articles per decade using life-table data to identify possible regulatory agents (i.e., those acting in a density dependent manner) for outbreak tree defoliators.
Experiments
Experimental approaches are also commonly used to study the processes influencing insect outbreak cycles (Myers and Rothman Reference Myers and Rothman1995). The scope and complexity of such experiments can vary significantly, ranging from fairly targeted studies assessing relationships among herbivores, host plants, and natural enemies, to more complex field experiments where the researchers have sought to manipulate outbreak cycles through preventing outbreaks, creating outbreaks, or manipulating the processes hypothesised to drive changes in population density (reviewed in Myers and Rothman Reference Myers and Rothman1995). In some cases, the theoretical advancements derived from experiments have been profound. For example, Holling’s (Reference Holling1959) experimental work on European pine sawfly–predator interactions was the cornerstone of his work on functional responses, a fundamental concept in animal population dynamics, and may be one of the most cited papers in The Canadian Entomologist’s history. Although we have not summarised the myriad experiments that have been carried out using outbreak forest defoliators, we have provided many noteworthy examples in the following section.
Case studies
In the following section, we feature case studies blending together the population research for some of the most-studied lepidopteran and sawfly defoliators in Canada, focussing in particular on those that have been examined through combinations of time-series, life-table, and experimental approaches. These case studies allow us to discuss details on impact, provide some historical context, and illustrate how knowledge gained through this work has helped to inform the development of management tactics and strategies to limit defoliator impact. Notably, we will not discuss here the extensive literature for conifer-feeding budworms (i.e., Choristoneura Lederer (Lepidoptera: Tortricidae) and Acleris Hübner (Lepidoptera: Tortricidae) species), which are covered in Nealis (Reference Nealis2015).
Lepidoptera
Fall webworm, Hyphantria cunea Drury (Lepidoptera: Arctiidae)
Fall webworm is a tent-forming defoliator, native to North America that is fairly innocuous compared with many of Canada’s most destructive defoliating pests. Outbreaks tend to have a limited impact and centre on open-grown or roadside speckled alder (Alnus incana (Linnaeus) Moench; Betulaceae) (Morris Reference Morris1971). Despite not being considered a major defoliator, the fall webworm was the focus of the earliest insect population studies ever published in Canada (Baird Reference Baird1917; Tothill Reference Tothill1922). To early entomologists, fall webworm was an intriguing model system, in part because it was easy to identify and sample, but also because it underwent relatively modest population fluctuations that seemed indicative of strong regulatory mechanisms (Tothill Reference Tothill1922; Morris and Fulton Reference Morris and Fulton1970a). It was expected that building an understanding of the processes regulating fall webworm populations might provide insight into the processes driving outbreaks of some of the more injurious defoliators of the time, such as spruce budworm (Morris Reference Morris1963a) and European spruce sawfly (Neilson and Morris Reference Neilson and Morris1964).
The first population ecology research for fall webworm, and the first of its kind for a defoliator in Canada, was a crude time series that reconstructed outbreak history using anecdotal records extending back to the late 1700s (Baird Reference Baird1917). Tothill (Reference Tothill1922) complemented this work with a more detailed eight-year time series collected by sighting tents from a slow-moving vehicle in the area surrounding Fredericton, New Brunswick, Canada. Morris (Reference Morris1964) revisited these old data and expanded upon them using FIDS data from Nova Scotia to Saskatchewan. Collectively, these approaches suggested an outbreak periodicity of 8–16 years and some regional spatial synchrony, although these data were not analysed statistically (Table 2). Tothill (Reference Tothill1922) also published one of the first examples of life-table data in Canada using fall webworm, although the information it provided was quite basic by today’s standards. Research on fall webworm increased significantly between 1957 and 1973 thanks to a series of field and laboratory studies in New Brunswick and Nova Scotia carried out by R.F. Morris and collaborators. Although starvation sometimes reduced colony survival at peak density, it was primarily parasitism and predation that were thought to regulate populations (Morris Reference Morris1972a, Reference Morris1972b, Reference Morris1976a). Interactions with natural enemies, however, were greatly complicated by factors such as larval size (Morris Reference Morris1963b), annual variability in parasitoid egg encapsulation by caterpillars (Morris Reference Morris1976b), colony size (Morris Reference Morris1972a, Reference Morris1976c), and hyperparasitism (Morris Reference Morris1976d). Survival also varied annually and regionally in response to variations in weather and the associated capacity of larvae to accumulate sufficient degree-days to enter diapause (Morris and Fulton Reference Morris and Fulton1970a, Reference Morris and Fulton1970b).
In the end, the intended simulation model was never built from the pieces provided by these experiments, and fall webworm continues to be a fairly inconsequential pest of roadside alder. However, Morris’ work has provided an invaluable basis for comparison for researchers in other countries where fall webworm has become an important invasive pest of fruit crops (Hidaka Reference Hidaka1977).
Winter moth, Operophtera brumata Linnaeus (Lepidoptera: Geometridae)
Winter moth is an invasive defoliator of a variety of deciduous trees that was introduced from Europe into Nova Scotia in the mid 1930s (Hawboldt and Cuming Reference Hawboldt and Cuming1950) and again onto Vancouver Island, British Columbia around 1972 (Gillespie et al. Reference Gillespie, Finlayson, Tonks and Ross1978). Much of the economic losses associated with defoliation by winter moth occurred on shade and ornamental trees in cities and towns, and occasionally in fruit orchards (Cuming Reference Cuming1961). In natural forests, red oak, Quercus rubra var. borealis (Michxaux) Farwell (Fagaceae), appeared to sustain particularly high defoliation, with potential indirect effects on the susceptibility of valuable white pine understorey to white pine weevil, Pissodes strobe (Peck) (Coleoptera: Curculionidae) (Embree Reference Embree1965). In Nova Scotia and later in Vancouver, two European parasitoids of winter moth, Cyzenis albicans (Fallén) (Diptera: Tachinidae) and Agrypon flaveolatum (Gravenhorst) (Hymenoptera: Ichneumonidae), were released as part of a classical biological control programme (Embree Reference Embree1965). Within just a few years of release, winter moth populations collapsed, and there have been no large-scale outbreaks since (Embree Reference Embree1965; Roland Reference Roland1988).
Population trends in Nova Scotia have been examined in time series covering the peak to decline (Embree Reference Embree1965) and the subsequent maintenance of winter moth populations at low densities following the introduction of parasitoids (Embree Reference Embree1991). Life-table studies have also been carried out for winter moth in Nova Scotia (Embree Reference Embree1965), British Columbia (Roland Reference Roland1988, Reference Roland1994), and its native range in Europe (Varley and Gradwell Reference Varley and Gradwell1968). There has been some debate whether the introduced parasitoids were solely or only indirectly the cause of winter moth decline. Whereas Embree (Reference Embree1965, Reference Embree1991) argued for the importance of the introduced parasitoids to population collapse and subsequent regulation at low density, Roland (Reference Roland1988) attributed the population decline to pupal predation associated with larval ground beetles (Coleoptera: Carabidae), with parasitism adding significant mortality but playing a more indirect, non-regulatory role. Interestingly, the European parasitoids that were introduced have very little impact on winter moth populations in its native European range, where pupal mortality associated with predation is the main regulatory factor (Varley and Gradwell Reference Varley and Gradwell1968). Roland (Reference Roland1994) continued his life-table studies on Vancouver Island for six years after the decline of winter moth to determine what processes were keeping populations low. Although parasitism still caused significant mortality, pupal predators were the main factor regulating populations at low density through a direct density-dependent response to pupal density (Roland Reference Roland1994). Winter moth populations have since remained relatively low in Canadian oak forests, although small outbreaks do still sometimes occur in fruit orchards (Embree Reference Embree1991).
This strategy of classical biological control for winter moth, which was largely validated through the aforementioned population studies, continues to be the primary approach for managing new winter moth introductions (e.g., Donahue Reference Donahue2015).
Forest tent caterpillar, Malacosoma disstria Hübner (Lepidoptera: Lasiocampidae)
Forest tent caterpillar outbreaks occur across Canada and can cause major defoliation to a variety of broadleaf hosts. Outbreaks have been noted at least anecdotally in North America as far back as 1790 (Baird Reference Baird1917). The colonial distribution of eggs and larvae make this group highly convenient for population studies, and few defoliators in Canada have been studied more intensively.
Whether forest tent caterpillar outbreaks are periodic or irregular has been the subject of extensive analyses and debate. The earliest work by Baird (Reference Baird1917) examining historical trends suggested a periodicity of 7–14 years (Table 2). More recent time-series analyses have taken advantage of long-term defoliation surveys and suggest an outbreak periodicity of 6–16 years, with the most commonly reported average being 10 years (Table 2). Time-series analyses by Cooke and Lorenzetti (Reference Cooke and Lorenzetti2006) indicate that population oscillations are “characterized by strong second-order negative feedback, suggesting regulation by lagged density-dependent processes”. Outbreaks appear to occur with some degree of spatial synchrony, although the degree of synchronisation may vary regionally (Hildahl and Reeks Reference Hildahl and Reeks1960; Sippell Reference Sippell1962; Cooke and Lorenzetti Reference Cooke and Lorenzetti2006; Cooke et al. Reference Cooke, MacQuarrie and Lorenzetti2012).
Early studies of forest tent caterpillar dynamics attributed increases or declines in population density primarily to early-spring and overwintering temperatures (Wellington Reference Wellington1952; Blais et al. Reference Blais, Prentice, Sippell and Wallace1955; Witter and Kulman Reference Witter and Kulman1972; Ives Reference Ives1973). However, when this hypothesis was tested explicitly by Daniel and Myers (Reference Daniel and Myers1995), using records from Ontario, the authors were unable to detect any relationship between interannual variation in defoliation (a proxy for population density) and either winter or spring temperatures. They did, however, report that defoliation was less severe in areas with the coldest winter temperatures, and suggested that extreme weather conditions may play a role in synchronising populations (e.g., “Moran effects”, Myers Reference Myers1998). More recent studies suggest that winter temperatures may be important determinants of outbreak duration in northern climates (Cooke and Roland Reference Cooke and Roland2003), albeit secondary to the effects of forest fragmentation (Roland et al. Reference Roland, Mackey and Cooke1998; Cooke and Roland Reference Cooke and Roland2000). Aside from weather influences, outbreak collapse may be caused by a nucleopolyhedrovirus (Clark Reference Clark1958; Stairs Reference Stairs1966; Bird Reference Bird1969; Ebling and Kaupp Reference Ebling and Kaupp1995) and to a lesser extent a parasitic fly, Sarcophaga aldrichi Parker (Sippell Reference Sippell1962).
As mentioned above, there is also strong evidence that forest tent caterpillar dynamics are influenced by forestry and land-development practices, with the degree of forest fragmentation being a major determinant of outbreak duration (Roland Reference Roland1993; Roland et al. Reference Roland, Mackey and Cooke1998; Cooke and Roland Reference Cooke and Roland2000; but see Wood et al. Reference Wood, Parry, Yanaia and Pitel2010). Roland (Reference Roland2005) also found that populations exhibited more rapid cycles in contiguous than in fragmented forests and concluded that forest structure had a significant impact on the lagged density-dependent processes that drive outbreak cycles, in particular through its effects on natural enemies (Rothman and Roland Reference Rothman and Roland1998; Roland Reference Roland2005). For example, forest tent caterpillar larvae at the edge of fragmented forests may have reduced virus transmission (Roland and Kaupp Reference Roland and Kaupp1995) and lower mortality associated with at least some parasitoids (Roland and Taylor Reference Roland and Taylor1997; Roth et al. Reference Roth, Roland and Roslin2006).
Since the earliest work by Baird (Reference Baird1917), forest tent caterpillar has provided an ideal model system for developing and testing theories of population dynamics for outbreaking defoliators. The practical implications of this long-term study include the revelations of how forestry practices, such as those that lead to increasing forest fragmentation, may exacerbate the intensity of damage caused by some outbreak-prone pests.
Hymenoptera (sawflies)
European spruce sawfly, Diprion hercyniae Hartig (Hymenoptera: Diprionidae)
European spruce sawfly is an invasive defoliator of mature foliage in spruce (Picea Dietrich; Pinaceae) that was first discovered on the Gaspé Peninsula of eastern Québec in 1930 (Neilson and Morris Reference Neilson and Morris1964). Incidentally, it was this discovery that has been largely credited with hastening the establishment of the long-standing Forest Insect and Disease Survey programme in Canada (Wallace and Cunningham Reference Wallace and Cunningham1995). By 1938, an outbreak was raging throughout much of the Maritimes and New England and prompted the introduction of a comprehensive classical biological control programme from 1933 through 1945 (Reeks Reference Reeks1953). By 1936, however, a highly infectious virus (a nucleopolyhedrovirus) was also observed in reared larvae and, by 1939, had spread through the population. It was speculated that the virus was vectored into the population by some of the introduced parasitoids (Balch and Bird Reference Balch and Bird1944; Bird and Elgee Reference Bird and Elgee1957). By 1945, the sawfly outbreak had collapsed and has not since reached the widespread, epidemic population levels seen during that outbreak.
There are limited long-term population studies of European spruce sawfly in Canada and only a handful of experiments, most of which focus on the mechanisms of virus transmission (Neilson and Elgee Reference Neilson and Elgee1968). However, there is at least one detailed 25-year population study that was conducted in New Brunswick from 1938 through 1961, which spanned the peak to collapse of the outbreak, as well as the following 18 years when population densities remained low (Neilson and Morris Reference Neilson and Morris1964). This study provided a rare opportunity to study not only the factors driving population collapse, but also the processes that maintained the population at low levels after collapse. Virus and weather appeared to be the “key factors” driving population collapse, but a combination of parasitism and cocoon predation was largely responsible for regulating populations at low levels after the collapse (Neilson and Morris Reference Neilson and Morris1964). Complementary experiments where the virus was introduced into populations with no previous infection further confirmed the efficacy of the virus (Bird and Elgee Reference Bird and Elgee1957). An incidental treatment of dichlorodiphenyltrichloroethane (DDT) to the study sites from 1960 through 1962, which was actually meant to suppress a local spruce budworm outbreak, essentially eradicated the local population.
The biological control work for European spruce sawfly led to the discovery and ultimate establishment of the virus into Canada, and the population studies that followed confirmed the efficacy of this agent for suppressing outbreaks. Although there have been a few local spikes in populations reported in the past few decades, there have been no more significant regional outbreaks since the virus was introduced (Forest Insects and Disease Survey 1936–1995).
Swaine jack pine sawfly, Neodiprion swainei (Middleton) (Hymenoptera: Diprionidae)
The Swaine jack pine sawfly is a major but infrequent native pest of mature jack pine, Pinus banksiana Lambert (Pinaceae), which has been studied primarily in Ontario and Québec. As of the 1970s, there were approximately two million ha of jack pine stands that were considered susceptible to sawfly outbreaks (McLeod Reference McLeod1970). At the time, jack pine was a cheap and easily harvested resource for the pulp and paper industry, and the extensive tree mortality that often followed outbreaks was cause for significant concern (McLeod Reference McLeod1970). Between 1940 and 1970, outbreaks of jack pine sawfly had a periodicity of around eight years and may have been synchronised regionally (McLeod Reference McLeod1972), although no sufficiently long time series have been published to confirm this periodicity. Although outbreaks are often intense, leading to complete tree-wide defoliation, they are also typically short, often lasting only two to five years (McLeod Reference McLeod1970, Reference McLeod1972). Surveys indicated, however, that when outbreaks occurred, populations rose quickly. During an outbreak in Roberval, Québec during the 1960s, for example, populations rose at a rate of 2.6-fold per year (McLeod Reference McLeod1970). At such high growth rates, populations reached tree-killing densities within as few as four years, and peak larval densities rose as high as 9.8 million per ha (McLeod Reference McLeod1970).
Although there are no published time series or life tables for jack pine sawfly, there has been some experimental work attempting to disrupt cycles through the use of insecticides. An experiment carried out in Québec from 1962 to 1967 found that a single, direct insecticide (phosphamidon) application diminished population densities and disrupted the cycling of populations relative to other populations in the area for at least five years following spray; however, populations in surrounding areas that suffered only a light insecticide application quickly regained normal cycling (McLeod Reference McLeod1972). The rebound of the lightly treated populations was associated primarily with recovery of larval survival rates rather than with immigration of egg-laying females from nearby populations (McLeod Reference McLeod1972). This study suggested that the processes synchronising jack pine sawfly population cycles over broad spatial areas are relatively robust, but that they can be overwhelmed by strong perturbations.
There remain many gaps in the study of jack pine sawfly population dynamics, in particular a conspicuous lack of long-term time-series and life-table studies. However, McLeod’s (Reference McLeod1972) experimental work is noteworthy as it provides a rare example of a study that has taken advantage of insecticide applications to better understand the population behaviour of an outbreak herbivore.
Larch sawfly, Pristiphora erichsonii Hartig (Hymenoptera: Tenthredinidae)
Larch sawfly is a Holarctic defoliator with a distribution that essentially follows that of larch, Larix Miller (Pinaceae), throughout the northern hemisphere. Major outbreaks in Canada have been credited with killing large swaths of larch since the late 19th century (Fyles Reference Fyles1892; Ives and Muldrew Reference Ives and Muldrew1984). A striking example is provided by Fyles (Reference Fyles1892), who reported a township in Québec that lost virtually all of its merchantable tamarack (Larix laricina (Du Roi) Koch) in a 260-ha stand to a larch sawfly outbreak. Similarly intense outbreaks of larch sawfly have been reported repeatedly across Canada over the past century (Ives and Muldrew Reference Ives and Muldrew1984).
Time-series studies of larch sawfly have focussed largely on establishing that larch sawfly is a native defoliator to Canada (Ives and Muldrew Reference Ives and Muldrew1984; Nishimura and Laroque Reference Nishimura and Laroque2010). The first recorded outbreak in North America occurred along the eastern seaboard in 1880 and then appeared to spread north and west – this outbreak was speculated to represent the unintentional introduction of larch sawfly to North America from Europe (Fyles Reference Fyles1892; Hewitt Reference Hewitt1912; Ives and Muldrew Reference Ives and Muldrew1984). However, evidence from tree-ring growth patterns in tamarack from Québec and Labrador indicates that larch sawfly outbreaks have a long history in Canada that far predates the 1880s (Jardon et al. Reference Jardon, Filion and Cloutier1994; Nishimura and Laroque Reference Nishimura and Laroque2010). Although the periodicity of outbreaks is highly variable (i.e., between 14 and 77 years), there is some observational evidence of regional synchronicity in outbreaks (Table 2).
Early studies indicated that there were many factors causing sawfly mortality in high-density populations, including starvation (Heron Reference Heron1955; Ives Reference Ives1963), storms and flooding (Lejeune Reference Lejeune1955; Graham Reference Graham1956; Ives Reference Ives1976), larval and cocoon predation (Buckner Reference Buckner1958, Reference Buckner1959; Buckner and Turnock Reference Buckner and Turnock1965), and some limited parasitism (Turnock and Muldrew Reference Turnock and Muldrew1971, Reference Turnock and Muldrew1972; Ives Reference Ives1976). A life-table study in southeastern Manitoba from 1956 to 1972 concluded that mammal predation of cocoons was the key factor driving population cycles, although periodic flooding events also appeared to contribute to population collapse (Ives Reference Ives1976). To mitigate an apparent paucity of mortality agents, an extensive programme of classical biological control was implemented and eventually resulted in the successful establishment of two European parasitoids, Olesicampe benefactor Hinz and Mesoleius tenthredinis Morley (Hymenoptera: Ichneumonidae) (Hewitt Reference Hewitt1917; Turnock and Muldrew Reference Turnock and Muldrew1971). Parasitism by O. benefactor in particular has been an important factor in keeping populations low, although its effectiveness may be offset to some extent by a hyperparasitoid, Mesochorus dimidiatus Holmgren (Hymenoptera: Ichneumonidae) (Ives Reference Ives1976).
Population studies were instrumental in confirming larch sawfly was a native pest to Canada with a long history of outbreak. No large-scale outbreaks of larch sawfly have occurred in recent decades, which may be related to the extensive biological control work that has been carried out; however, although studies were able to show the successful establishment of O. benefactor, it has yet to be confirmed that this parasitoid is responsible for keeping populations low over the long term.
Balsam fir sawfly, Neodiprion abietis (Harris) (Hymenoptera: Diprionidae)
Balsam fir sawfly is a major native defoliating pest of mainly balsam fir, Abies balsamea (Linnaeus) Miller (Pinaceae). Although this sawfly has a transcontinental distribution in North America (Ross Reference Ross1955; Wallace and Cunningham Reference Wallace and Cunningham1995), outbreaks are most common in Atlantic Canada (Martineau Reference Martineau1985). In the late 1990s, an outbreak in central Newfoundland became a source of major concern for the local pulp and paper industries due to its unusually high severity, geographical extent, and longevity (Moreau Reference Moreau2006), prompting nearly a decade of intensive research to explain the cause of the outbreak and to develop control strategies to reduce damage.
Outbreaks are typically localised but can be quite severe, sometimes resulting in almost complete tree defoliation (current-year foliage is rarely eaten); however, such outbreaks are also usually short in duration, rarely lasting more than three to six years (Cunningham Reference Cunningham1984; Moreau Reference Moreau2006). In western Newfoundland, historical time-series records of defoliation spanning over 60 years indicated an outbreak periodicity of about six years, with evidence of oscillatory population behaviour (Moreau Reference Moreau2006). Moreau (Reference Moreau2006) noted that increases in outbreak severity and duration corresponded to increased use of precommercial thinning, a forestry practice that involves removing stems to reduce competition between neighbouring trees. Contemporaneous studies in the same region supported this hypothesis, showing that at peak outbreak, egg density and defoliation due to larval feeding were higher in thinned than in unthinned stands (Ostaff et al. Reference Ostaff, Piene, Quiring, Moreau, Farrell and Scarr2006).
To determine why precommercial thinning increased the intensity of outbreaks, a life-table study was conducted in western Newfoundland from 1999 to 2002 (Moreau Reference Moreau2004). Although the number of generations sampled within individual sites was relatively low (i.e., three to four generations), a range of sites was selected to represent late-rising, peaking, and collapsing populations, thereby providing a fair snapshot of the broader outbreak cycle in thinned and unthinned stands. In general, rising populations were characterised by relatively high fecundity, female-biased sex ratios, high late-instar survival, and immigration, all of which translated into enhanced intergenerational recruitment rates (Moreau Reference Moreau2004). In contrast, the collapse of populations was driven primarily by increased infection rates by a virus (nucleopolyhedrovirus, NeabNPV), which caused extreme larval mortality and moderate prepupal and pupal mortality (Moreau Reference Moreau2004; Moreau et al. Reference Moreau, Eveleigh, Lucarotti and Quiring2006a). Mortality associated with a generalist parasitoid, Mesopolobus verditer (Norton) (Hymenoptera: Pteromalidae), also contributed to population declines in some sites. Although the duration of the outbreak peak to collapse was similar between thinned and unthinned stands (i.e., two to four years), the amplitude of population fluctuations and associated defoliation was higher in thinned stands, owing at least in part to higher larval survival (Moreau Reference Moreau2004; Moreau et al. Reference Moreau, Eveleigh, Lucarotti and Quiring2006a; Ostaff et al. Reference Ostaff, Piene, Quiring, Moreau, Farrell and Scarr2006). Complementary experiments showed that precommercial thinning altered the relative and overall impact of bottom-up (host plant quality) versus top-down (natural enemies) forces on larval survival (Moreau et al. Reference Moreau, Eveleigh, Lucarotti and Quiring2006b) and reduced the negative influence of previous defoliation on foliage nutritional quality and associated larval performance (Moreau and Quiring Reference Moreau and Quiring2011).
Overall, these studies drew attention to the potentially negative influence of commercial forestry practices on defoliator outbreaks, but also provided the basis for developing a useful biological control agent (the virus) that has since become part of the integrated pest management programme for balsam fir sawfly (Moreau et al. Reference Moreau, Lucarotti, Kettela, Thurston, Holmes and Weaver2005).
Conclusions
Our review underscores the tremendous efforts of researchers to address the fundamental questions driving population ecology research on outbreak defoliators in Canada. Comparative studies, although few in number, have provided interesting insights on the key traits possessed by outbreak species and lacking in typically non-outbreak species. There have been a fair number of time series and life-table data sets collected for the major outbreak species in Canada, particularly given the significant cost and commitment required to collect these data. Qualitative examination of time-series data provides strong indications of both regular periodicity and spatial synchrony in outbreaks of major defoliators (Table 2). Life-table studies highlight the particular importance of top-down trophic interactions for driving the decline and maintenance of populations at low density (Table 3). Putting aside for now the important theoretical advancements that we have highlighted in this review, we will conclude by addressing questions of a more practical nature: after more than 100 years of research and countless millions spent, what has the study of defoliator population dynamics achieved? And, what work still lies ahead?
What have we achieved? Broadly speaking, we would highlight several practical advancements arising from Canadian population research for outbreak defoliators. Much of the research to date draws attention to the predominance of natural enemies in regulating populations, although recent work has reintroduced the potential importance of host quality as an indirect factor influencing population decline (e.g., Régnière and Nealis Reference Régnière and Nealis2007; Moreau and Quiring Reference Moreau and Quiring2011). This insight has contributed to several striking examples of successful biological control for invasive species (e.g., winter moth and European spruce sawfly) and to some extent native pests (e.g., larch sawfly and balsam fir sawfly). Moreover, insights into the temporal and spatial nature of defoliator outbreaks have come to form the basis of pest management strategies, which is perhaps best illustrated in the long-standing debate over whether to manage spruce budworm outbreaks through a “foliage protection” versus “population control” strategy (Royama Reference Royama1984; Régnière et al. Reference Régnière, Deslisle, Pureswaran and Trudel2012). Another important contribution of this work has been confirmation of the importance of forestry practices in affecting the behaviour and structure of defoliator populations (e.g., forest fragmentation, Roland Reference Roland1993, Reference Roland2005; precommercial thinning, Moreau et al. Reference Moreau, Eveleigh, Lucarotti and Quiring2006b; forest composition, Eveleigh et al. Reference Eveleigh, McCann, McCarthy, Pollock, Lucarotti and Morin2007). Understanding the role forest composition and structure play in shaping outbreaks provides a basis for selecting silvicultural practices aimed at reducing forest susceptibility to defoliator damage.
What lies ahead? There is no shortage of work to be done on outbreak defoliators, whether that be through revisiting older data sets, collecting more data, or harnessing new, innovative technologies. As we discussed previously, qualitative analyses of time-series data could be buttressed in the future with deeper quantitative analyses (e.g., Cooke et al. Reference Cooke, MacQuarrie and Lorenzetti2012). In a similar vein, there may be value in revisiting some past life-table data sets to see what further insights could be drawn from more advanced analyses (e.g., Morris Reference Morris1963a versus Royama Reference Royama1984). Technological advances during the past decades, in particular with respect to genomics (Cusson Reference Cusson2008), are becoming an invaluable component of population, community, and landscape ecology involving outbreak defoliators, with many remarkable and sometimes unexpected revelations (e.g., Smith et al. Reference Smith, Eveleigh, McCann, Merilo, McCarthy and Van Rooyen2011; James et al. Reference James, Cooke, Brunet, Lumley, Sperling and Fortin2014). As our review highlights, there is also a paucity of studies of defoliator populations during the low density or rising phases of outbreak. This represents a significant knowledge gap and a potentially rich avenue for future investigation (e.g., Régnière et al. Reference Régnière, Deslisle, Pureswaran and Trudel2012). Finally, it is becoming increasingly evident that studies of outbreak defoliator population dynamics are going to need to be considered in the context of a changing climate (Pureswaran et al. Reference Pureswaran, De Grandpré, Paré, Taylor, Barrette and Morin2015). Not only is climate change likely to alter the intensity, distribution, and extent of common outbreak species (Gray Reference Gray2013; Pureswaran et al. Reference Pureswaran, De Grandpré, Paré, Taylor, Barrette and Morin2015), it also has the potential to promote outbreaks among species that have rarely or never been known to outbreak in the past.
As a final point, our review makes clear the benefits of long-term, persistent research in terms of our understanding of insect outbreaks for some of the major defoliator species in Canada (e.g., budworms and tent caterpillars). However, the trends and processes driving these outbreaks are not necessarily representative of all species that have been reported to outbreak at one time or another. Indeed, the apparently cyclic and synchronous nature of most of the half-dozen major outbreaks species in Canada are at odds with the majority of species, most of which undergo relatively sporadic and localised outbreaks. One particularly striking omission is the paucity of research on outbreaking coleopteran defoliators, although this is probably due to the relative infrequency, brevity, and limited distribution of outbreaks, as well as the low economic value of the host trees attacked (i.e., mainly Populus Linnaeus; Salicaceae and Salix Linnaeus; Salicaceae). Studies of these and other defoliators that undergo more local or sporadic outbreaks (or that never outbreak at all) could provide a deeper perspective on the overarching ecological question of why animal populations fluctuate as they do.
Acknowledgements
The authors thank H. Blaquiere and Z. Fitch for helping to gather materials for the large defoliator table, and G. Pohl and H. Goulet for providing estimates of the total number of defoliators in Canada. We thank R. Alfaro, S. Heard, D. Langor, C. MacQuarrie, E. Moise, Z. Sylvain, D. Quiring, and several anonymous reviewers for comments and discussions on earlier versions of the manuscript.







