No CrossRef data available.
Published online by Cambridge University Press: 19 September 2024
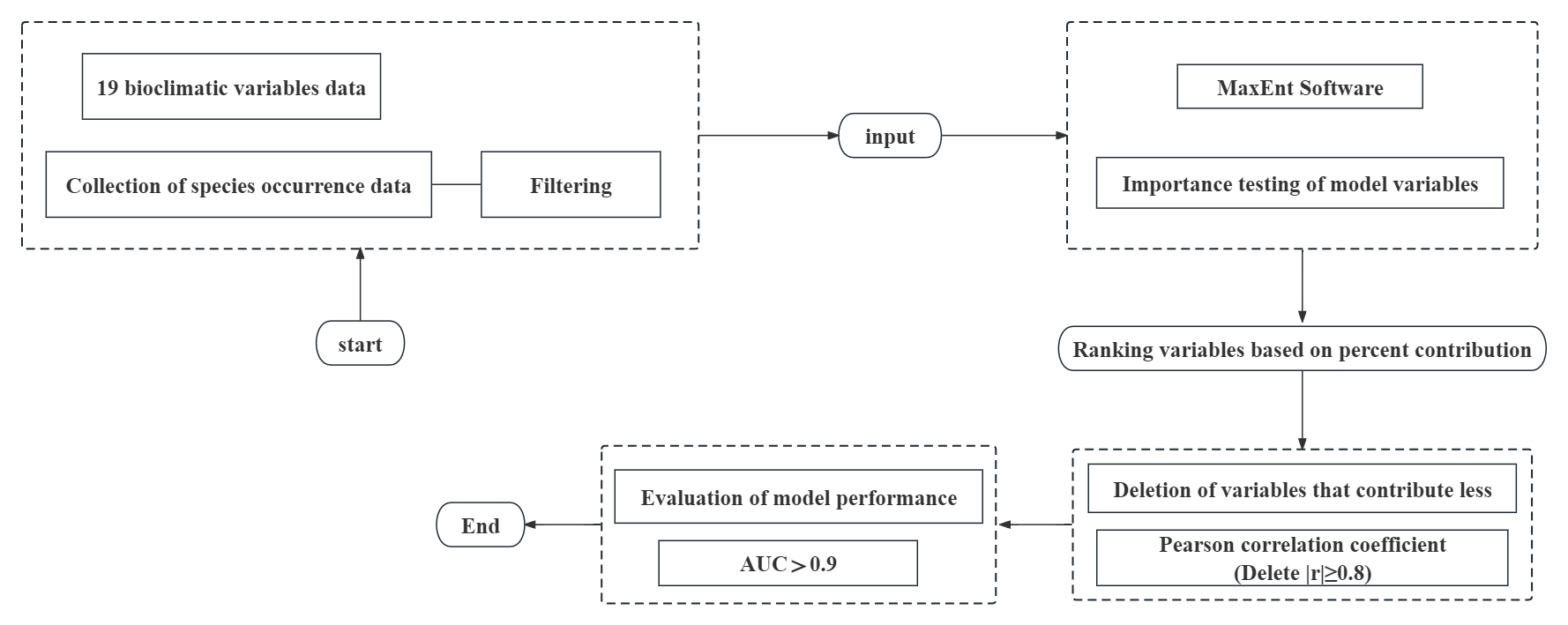
Changes in the distribution of species due to global climate change have a critically significant impact on the increase in the spread of invasive species. An in-depth study of the distribution patterns of invasive species and the factors influencing them can help to better predict and combat invasive alien species. Rhynchophorus ferrugineus Olivier is an invasive species that primarily harms plants of Trachycarpus H. Wendl. The pest invades trees in three main ways: by laying eggs and incubating them in the crown of the plant, on roots at the surface and at the base of the trunk or petiole. Most of the plants in the genus Trachycarpus are taller, and the damage is concentrated in the middle and upper parts of the plant, making control more difficult. In this paper, we combine 19 bioclimatic variables based on the MaxEnt model to project the current and future distributions of R. ferrugineus under three typical emission scenarios (2.6 W m−2 (SSP1-2.6), 4.5 W m−2 (SSP2-4.5) and 8.5 W m−2 (SSP5-8.5)) in the 2050s and 2090s. Among the 19 bioclimatic variables, five variables were screened out by contribution rates, namely annual mean temperature (BIO 1), precipitation of driest quarter (BIO 17), minimum temperature of coldest month (BIO 6), mean diurnal range (BIO 2) and precipitation of wettest quarter (BIO 16). These five variables are key environmental variables that influence habitat suitability for R. ferrugineus and are representative in reflecting its potential habitat. The results showed that R. ferrugineus is now widely distributed in the southeastern coastal area of China (high suitability zone), concentrating in the provinces of Hainan, Guangdong, Fujian, Guangxi and Taiwan. In the future, the area of high and low suitability zones will increase and the area of medium suitability zones will decrease. The area of low suitability zone will still be in the largest proportion. This study aims to provide a theoretical reference for the future control of R. ferrugineus from the perspective of geographic distribution.
These authors contributed equally to this work and should be regarded as co-first authors.