


Introduction
Gurney's Pitta Pitta gurneyi is the only bird species endemic to peninsular Thailand and Myanmar where it inhabits Sundaic lowland forests. Sundaic forest is perhaps the world's most threatened ecosystem (Lambert and Collar Reference Lambert and Collar2002). In Thailand and Myanmar, the greatest threat is clearance for oil palm, which supports few forest species and low biodiversity (Aratrakorn et al. Reference Aratrakorn, Thunhikorn and Donald2006).
First described from Burma (now Myanmar) in 1875, Gurney's Pitta has a chequered ornithological history. Early records suggest that it was common in semi-evergreen forests between around 7°N and 12°N (Collar et al. Reference Collar, Round and Wells1986), within which range it was probably resident (Round Reference Round1995). However, numbers must have declined rapidly prior to 1952, since it was not recorded in the wild between that year and its rediscovery in a small area in southern Thailand in 1986 (Round and Treesucon Reference Round and Treesucon1986). At this site, around Khao Nor Chuchi in Krabi Province, the population by 1989 was estimated at just 20–30 pairs and appeared to be heading to extinction due to rapid forest loss (Gretton et al. Reference Gretton, Kohler, Lansdown, Pankhurst, Parr and Robson1993). The failure of several conservation efforts to do more than slow the rate of decline led in 2002 to the formulation and agreement of a Gurney's Pitta Species Recovery Plan (GPSRP), adopted by representatives of the relevant Thai authorities, national and international NGOs, and a range of local stakeholders. The GPSRP included a range of measures, including research, which needed to be put in place to save the population from extinction. The results presented in this paper make a contribution to the technical actions agreed in the GPSRP.
In 2003, the species was rediscovered in Tanintharyi Division in southern Myanmar in what was assumed, because of the extent of apparently suitable forest, to be considerable numbers (Eames et al. Reference Eames, Htin, Leimgruber, Kelly, Sein Myo, Moses and U Saw Nyunt2005). Based largely on observations in southern Thailand, it has been suggested that Gurney's Pitta is a species of level, lowland forest generally below 100 m a.s.l., possibly with a preference for secondary or degraded forest (Collar et al. Reference Collar, Round and Wells1986, Round Reference Round1995, BirdLife International 2001). Because of this, preliminary surveys in Myanmar were restricted to flat, lowland forest below 160 m a.s.l. and the resulting population estimate of around 5,000 to 8,500 pairs based upon the assumption that birds were restricted to this habitat (Eames et al. Reference Eames, Htin, Leimgruber, Kelly, Sein Myo, Moses and U Saw Nyunt2005). On the basis of this estimate, the species's IUCN Red List threat category was down-listed from ‘Critically Endangered’ to ‘Endangered’ in 2007 (BirdLife International 2008).
This paper presents the results of research on the distribution, habitat use and breeding ecology of the species undertaken in Thailand from 2004 to 2008 and in Myanmar in 2007 and 2008.
Methods
Myanmar
Fieldwork teams visited the area from March to May 2007 and again from March to April 2008. These periods coincided with the species's main period of territorial activity (Gretton et al. Reference Gretton, Kohler, Lansdown, Pankhurst, Parr and Robson1993, Round Reference Round1995, Eames et al. Reference Eames, Htin, Leimgruber, Kelly, Sein Myo, Moses and U Saw Nyunt2005). Access into the forest was opportunistic and greatly restricted by the small number of roads and tracks, the weather and the security situation, so a systematic survey of the whole of the lowland forest area of southern Tanintharyi was not possible (Figure 1). Point transects were used to estimate the distribution and habitat use of Gurney's Pitta, with observation points separated by at least 100 m. Observation periods began without playback to collect unbiased data suitable for distance sampling estimation of density. After 10 minutes of recording, playback was used to determine the species’ presence followed by a further 10 minutes of recording, a period within which another pitta species, Pitta nympha, in Taiwan usually responded when present (Lin et al. Reference Lin, Lee, Ding and Lin2007a). Observations after playback were not considered suitable for distance sampling because playback might draw birds closer to the observer before they responded, leading to inflated density estimates. As too few contacts were made during the pre-playback period to allow density estimation, the population was assessed by modelling the distribution and applying the range of densities recorded in Thailand to this distribution. This follows the approach of Eames et al. (Reference Eames, Htin, Leimgruber, Kelly, Sein Myo, Moses and U Saw Nyunt2005), but the extent of distribution differed from that used by those authors because the collection of more field data allowed a better estimate of the latitudinal, altitudinal and environmental limits of the species.

Figure 1. Location of focal area (insert) and extent of remaining lowland evergreen forest (in green) in southern Myanmar, based on Stibig et al. Reference Stibig, Belward, Roy, Rosalina-Wasrin, Agrawal, Joshi, Hildanus, Beuchle, Fritz, Mubareka and Giri2007.
At each point visited in 2007, a number of habitat variables were collected (Table 1). In 2008, vegetation data were collected only at points over 150 m (as these were under-represented in 2007) and so the sample sizes for analyses of habitat structure was smaller than the total number of points visited.
Table 1. Vegetation structure recorded in Myanmar (at sites occupied and not occupied by Gurney's Pitta) and Thailand (from inside the core range of the species). Mean values are given. Mean values of each variable did not differ significantly (at P < 0.05) between sites sharing the same superscripted letter (ANOVA, post hoc Tukey tests). All variables differed significantly between the site in Thailand and one or both of the classes of site in Myanmar. dbh = diameter at breast height.

Collection of habitat variables:
1 Estimated by eye within three 1 m2 quadrats within 10 m of the point and averaged;
2 Average counted within three 1 m2 quadrats within 10 m of the point;
3 Number of clumps within 25 m of the point;
4 Number of trees within 25 m of the point.
Thailand
Complete surveys were undertaken in 2003, 2005 and 2007, during which all forest below 200 m a.s.l. within the known range of the species in southern Thailand was visited. Forest patches were identified from aerial photographs and satellite imagery and visited at least twice. Playback was used to assess or confirm the presence of birds. In 2004 and 2006, surveys were restricted to a core c. 130-km2 study area of Gretton et al. (Reference Gretton, Kohler, Lansdown, Pankhurst, Parr and Robson1993). Nests were searched for from March to October in each year. When active nests were found, a hide was erected around 20 m away and nests guarded round the clock in 6-hour shifts. At night, the nest and surroundings areas were briefly checked by torchlight at regular intervals and any predators intercepted. Detailed observations were taken on chick diet and provisioning rates.
Soil moisture measurements were taken using an electronic soil moisture meter along 12 gullies, half of them used regularly by Gurney's Pittas and the other half not used. Measurements were taken across 10 cross-sections of each gully. On each cross-section, a single measurement was taken at the top of each side, three measurements down each slope and a single measurement at the bottom of the gully, giving 90 measurements for each gully.
In April 2008, the same observer who collected habitat measurements in Myanmar (TWH) visited southern Thailand and collected habitat measurements using the same methods at a number of sites where Gurney's Pitta were recorded during recent fieldwork. The use of the same observer ensured comparability between countries in the habitat data collected.
Data analysis
The potentially intercorrelated habitat structure variables (Table 1) were reduced to a smaller number of uncorrelated axes using unconstrained ordination (Principal Components Analysis, PCA; McGarigal et al. Reference McGarigal, Cushman and Stafford2000). Standard parametric tests were used to compare PCA scores between occupied and unoccupied sites in Myanmar and occupied sites in Thailand to assess structural differences. Maximum entropy models were used to predict the distribution of Gurney's Pitta in Myanmar as a function of land cover, slope and altitude using Maxent 2.3 (Phillips et al. Reference Phillips, Anderson and Schapire2006). Land cover characteristics were derived from information collected by the SPOT – Vegetation sensor. This covers the globe daily, and collects data at a 1-km resolution. The maximal NDVI (Normalised Difference Vegetation Index) values over 10-day (dekad) periods for 2005 were extracted from http://free.vgt.vito.be. The 36 images that cover the entire area in 2005 were reduced by using PCA (in Erdas Imagine 8.7), with the top four components being used in further analysis. Topography data were derived from 90 m Shuttle Radar Topography Mission SRTM (USGS 2004), 30 arc seconds (Global Land Cover Facility, University of Maryland, College Park, Maryland, February 2000; www.landcover.org). These indicate spot heights every 90 m, and were used to calculate average altitude and slope (using the slope procedure in ArcMap) across 1-km squares throughout southern Myanmar south of 13° N. The coordinates of all confirmed presences were used (recent surveys and data from Eames et al. Reference Eames, Htin, Leimgruber, Kelly, Sein Myo, Moses and U Saw Nyunt2005). This presented problems, as some 1-km squares (the unit of the analysis) contained multiple point counts, and the likelihood of a bird being present in a 1-km square increases with the number of points surveyed (χ2 = 9.43, P < 0.005, n = 104). However it was not possible to correct for this in the software, and 1-km squares were recorded as occupied irrespective of amount of observer effort. A map of potentially suitable habitat was produced using geo-referenced occurrences and environmental data comparing the spatial environmental variables at locations where birds were recorded against data from a random selection of other locations across the focal area, which was the area of lowland forest in southern Myanmar shown in Figure 1. The 47 1-km squares in which birds were recorded were compared to 10,000 random putative absences. All non-terrestrial habitats were excluded from the analysis. All data layers were included in the analysis, but 15% of points were used as test data, not included in the model building, to assess the efficacy of the model.
Because the maximum altitude visited during the field surveys was 226 m (see results), up to which altitude there was no evidence of a reduction in the probability of recording the species, three Maxent models were produced. The first was fitted with altitude (producing models that predict the species's distribution based upon the observed altitudes), the second with a binary variable denoting above or below 500 m a.s.l. (to predict the distribution if the altitudinal limit were 500 m) and the third fitted without altitude (producing models that predict the species’ distribution based solely on land cover). Within each of these three modelled distributions, we applied territory densities of 2.8 km-2 and 6 km-2, being the extremes recorded within a 2.5 km2 forest study plot in southern Thailand shortly after the rediscovery of the species there (Gretton et al. Reference Gretton, Kohler, Lansdown, Pankhurst, Parr and Robson1993).
Results
Myanmar
In total 391 points were visited, 207 along 21 transects in 2007 and 184 along 29 transects in 2008 (Figure 1). Of these, habitat data were collected at 220 points. Gurney's Pitta was recorded at 101 (25.8%) points, either before playback (11% of first contacts) or after (89%). Too few contacts were made before playback to allow density estimation using distance sampling.
There were significant differences in forest structure between points where birds were recorded (before or after playback) and points where they were not recorded (Table 1). Logistic regression with backward deletion retained four variables (number of bamboo stands, density of seedlings, number of rattan trees and number of trees 1–10 cm dbh) which were uncorrelated and contributed independent significant (all P < 0.02) explanatory power. All were positively related to the likelihood of occurrence of Gurney's Pitta and the final model correctly assigned 70.3% of points.
Altitude of visited points ranged from 5 m a.s.l. to 226 m a.s.l., with an average of 60.8 m. There was no difference in altitude between occupied and unoccupied sites (F 1,389 = 2.04, P > 0.1), though the highest altitude at which birds were recorded was 184 m (Figure 2). Similarly, there was no association between the likelihood of occurrence and slope (χ23 = 4.37, P > 0.2).

Figure 2. Histogram of altitude (m a.s.l.) of points surveyed in Myanmar. The shaded portion indicates points at which Gurney's Pitta was recorded.
Gurney's Pitta was recorded between 10°42’ N and 11°47’ N, the last some 20’ (approximately 36 km) north of the previous most northerly record in Myanmar at Lenya (Eames et al. Reference Eames, Htin, Leimgruber, Kelly, Sein Myo, Moses and U Saw Nyunt2005), and at almost the same latitude as the most northerly historical location in Thailand at 11°50’ N (BirdLife International 2001).
Maximum entropy models based upon land cover and topography (altitude and slope) predicted a range of 3,320 km2 (Figure 3). Darker red colours indicate greater chance of occurrence. The ROC value for the model was reasonably high (0.924), suggesting that the model was appropriate for predicting distributions. For the 15% of data held back as test data, the ROC was 0.851, which we consider satisfactory. The optimal cut-off for separating occupied and apparently unoccupied areas was 0.30, which meant that 13% of the focal area was predicted to be suitable, equating to approximately 3,320 km2. Most of the predicted range fell within the GLC2000 land cover (Stibig et al. Reference Stibig, Belward, Roy, Rosalina-Wasrin, Agrawal, Joshi, Hildanus, Beuchle, Fritz, Mubareka and Giri2007) class ‘Lowland evergreen forest’, though some also fell into squares classified as ‘Swamp forest/woodland’.

Figure 3. Modelled distribution (using maximum entropy) of potentially suitable Gurney's Pitta habitat in southern Myanmar, based on altitude, slope and SPOT – Vegetation data. Darker red colours indicate greater modelled suitability. Green points show locations where birds were recorded by current study and purple points are those where birds were recorded by Eames et al. (Reference Eames, Htin, Leimgruber, Kelly, Sein Myo, Moses and U Saw Nyunt2005).
When altitude was fitted as a binary variable relating to above or below 500 m a.s.l., the predicted range increased to 5,573 km2, and when altitude was removed from the models and the range predicted solely from land cover, the predicted range was 5,908 km2. Much of this lies within the known altitudinal range of Gurney's Pitta (Figure 4), so even if the species's altitudinal limit is significantly higher than previously indicated, this might not result in a much larger distribution.

Figure 4. Relationship between the area of predicted suitable habitat in southern Myanmar (from a model of distribution that includes land cover but excludes altitude) and the altitudinal limit of Gurney's Pitta, which is currently uncertain.
When combined with the extreme range of territory densities recorded in Thailand, the estimated population was predicted to lie between 9,296 territories (assuming a range of 3,320 km2 and a density of 2.8 pairs km−2) and 3,5076 (assuming a range of 5,846 km2 and a population density of 6.0 pairs km−2). If the true range size and population density fall towards the mid point of the range of estimates, the population can be estimated at 20,165 territories.
Thailand – nesting
In Thailand, 24 nests were found, of which eight were found when active (at four of which 24-hour watches were established). Nests were built almost exclusively in spiny Calamus (rattan) or Salacca palms (20 of 24 nests), with smaller numbers in climbers (n = 2) and the canopy of other trees (n = 2). Nests were located 0.7–6.2 m above ground level (mean = 3.2 m, SE = 0.37, n = 24) and tended to be built close to streams, pools or flooded gullies (mean distance to water = 14.5 m, SE = 3.3, n = 24) and to trails (mean distance = 20 m). Clutch sizes of two eggs (n = 1), three eggs (n = 2) and four eggs (n = 2) were recorded. The incubation period was around 17 days. Of the 18 nests for which an outcome could be determined with any degree of confidence, only four successfully produced fledged chicks, and three of these were nests that were guarded around the clock. This might underestimate nest success, as damaged nests could have been damaged following successful breeding. However, intensive nest guarding confirmed the high predation rate, as during a total of 53 days and nights of observation at four nests, 11 snake attacks were observed and prevented. All attacks took place at night between 20h00 and 05h00. If it is assumed that each attack would have led to complete nest failure, this equates to a daily Mayfield nest survival estimate (Mayfield Reference Mayfield1975) during the chick stage of 0.792, giving an estimate of overall nest survival rate over the 16 days the chicks are in the nest of just 2.4%. Because of removal of snakes from the vicinity of the nest, three out of four protected nests survived to produce at least one fledgling (the fourth failed when a snake was not detected in time). All snakes were of the genus Boiga (cat snakes), a genus of generalist and adaptable predators. The species identified in attacks on guarded nests included Mangrove Snake B. dendrophila, Dog-toothed Cat Snake B. cynodon, Jasper Cat Snake B. jaspidea and Grey Cat Snake B. ocellata.
After hatching, chicks remained in the nest for 13–15 days before leaving. Intensive observations at four nests identified a range of food items but the diet of chicks was clearly dominated by earthworms (Table 2).
Table 2. Identity of items brought to nests by adult Gurney's Pittas feeding chicks in southern Thailand. Tabulated values are percentages of all items.

Thailand – population and distribution
Accurate determination of the number of territories of Gurney's Pitta in Thailand is extremely difficult because of the frequent movements of calling birds. However, the plotting of near-simultaneous contacts suggested a population of 15–20 territories throughout the study period of 2003 to 2007. Minimum numbers of territories were estimated at 16 in 2003, 20 in 2005 and 15 in 2007. In each year, all birds were recorded in the Khao Nor Chuchi lowland forest, including 2–3 territories at Ao Tong in neighbouring Trang province. In total, birds were recorded at 27 sites over the study period, more than half of them in the Bang Khram Reserved Forest, with the remainder in the Khao Pra-Bang Khram Wildlife Sanctuary (Table 3). All records were below 100 m a.s.l. with the exception of a single male recorded in 2003 moving through forest at 180 m a.s.l. (this bird was followed for over 500 m and was presumed to be moving between sites) and a male at 153 m a.s.l. in 2005. The Thai population lies approximately 300 km due south of the population in Myanmar.
Table 3. Number of confirmed Gurney's Pitta territories recorded in the Khao Pra-Bang Kram Wildlife Sanctuary and the Khao Nor Chuchi National Reserved Forest, southern Thailand.

* Surveys only undertaken in the core area of Gretton et al. (Reference Gretton, Kohler, Lansdown, Pankhurst, Parr and Robson1993), which included parts of both the Wildlife Sanctuary and the National Reserved Forest.
Soil moisture in gullies used by Gurney's Pittas was significantly lower along the tops (F 1,233 = 16.8, P < 0.0001) and sides (F 1,702 = 21.4, P < 0.0001) than gullies not used by birds, but significantly higher at the bottom of gullies (F 1,108 = 14.9, P < 0.0001), where birds preferentially fed.
There was evidence of a decline in the rate of loss of forest within the core area of the species in southern Thailand after 2000 (Figure 5), suggesting that recent conservation efforts may have had some success.
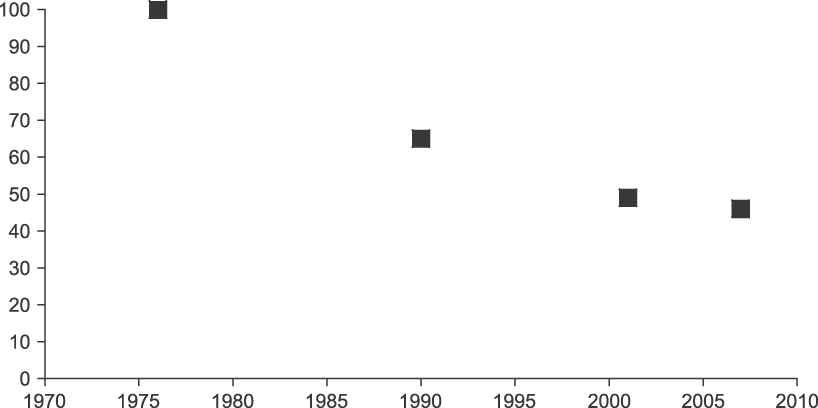
Figure 5. Changes in forest area in the core Gurney's Pitta area in southern Thailand, estimated from Landsat and ASTER remote imagery. Values show the percentage cover by forest (primary, secondary and regenerating) within the 133 km2 area shown in Figure 5 of Gretton et al. Reference Gretton, Kohler, Lansdown, Pankhurst, Parr and Robson1993 in four years. This area held most of the Gurney's Pittas recorded in southern Thailand in each year. There is evidence of a recent decline in the rate of forest loss, though the estimate from 2007 is subject to some doubt because of cloud cover during the satellite overpass.
Comparing habitats in Thailand and Myanmar
All measures of the structure of occupied forest in Thailand differed significantly from those in both occupied and unoccupied sites in Myanmar (Table 1), the Thai sites having in particular a higher density of saplings and small trees, spiny palms and rattan and a lower density of bamboo and large trees. There was consequently very little overlap between forest in Thailand and Myanmar in ordination space (Figure 6). Occupied sites in Myanmar were more similar in forest structure to unoccupied forests in Myanmar than they were to occupied sites in Thailand (Figure 6).

Figure 6. Scatterplot of scores of the first two principal components of an ordination of point locations by vegetation variables. Black circles: unoccupied sites in Myanmar, red squares: occupied sites in Myanmar; green diamonds: occupied sites in Thailand.
Discussion
All results must be interpreted in the light of the extreme difficulty of surveying this species. Without tape luring, encounters with the species are infrequent even in core areas during the main calling season. Even using tape luring it is difficult to estimate encounter probability. The recorded absence of birds from any area must therefore not be taken to indicate certain absence and the species is likely to be under-recorded where it occurs. However, the fact that the vegetation measurements in Myanmar varied non-randomly with respect to the recorded presence or absence of the species suggests that, within the lowland forest biome, birds do exhibit some measure of habitat selection. The Maxent models also suggest that not all lowland evergreen forest in southern Myanmar is suitable for the species, and indeed there is a considerable difference between forested areas in southern Myanmar (Figure 1) and the predicted distribution of the species (Figure 3). Although the forest structure of sites where the species was recorded in Myanmar differed greatly from that at sites in Thailand (Figure 6), it was interesting to note that occupied sites in Myanmar held a significantly higher density of seedlings and saplings and more rattans than unoccupied sites, reflecting perhaps a preference for disturbed or regenerating forest. In Thailand, sites occupied by Gurney's Pitta are largely secondary forest of less than 50 years old, regenerating from previous pastures. The use of secondary forest in Thailand has previously been assumed to reflect the almost complete loss of primary forest on low, flat land, forcing the remaining population into sub-optimal alternatives, but the results from Myanmar provide some support for a preference for disturbed or secondary habitats. The greatly differing habitat structures recorded in occupied sites in Thailand and Myanmar suggest that Gurney's Pitta can occupy a range of forest types and might be able to tolerate a significant degree of forest disturbance. It also appears that the species occupies a greater altitudinal range than previously supposed, as there was no evidence of a decline in encounter probability up to the highest visited altitude of 226 m in Myanmar. It has been suggested that this wider altitudinal range might reflect a lack of competition with Banded Pitta Pitta guajana, a species which is apparently absent from southern Myanmar but which might compete with Gurney's Pitta in southern Thailand (P. D. Round in litt). The modelled distribution of the species in Myanmar suggests an extent of occurrence of at least 3,320 km2, rising to nearly 6,000 km2 if the altitudinal limit is higher than currently thought. The model suggests that populations might occur as far north as 12°20’ N, some 60 km north of the most northerly sightings made during the present study, and as far south as 10°35’ N. Further, it indicates that some areas within western Thailand may still hold suitable habitat, despite being distant from the known occupied areas within this country. In addition to validating these distribution maps, further work needs to be undertaken in Myanmar to clarify the altitudinal and latitudinal limits of the species, as this will allow much more precise estimation of the species’ distribution and population. Forests in Thailand that are currently considered too high for the species might actually hold birds.
The range of population estimates from Myanmar must be treated with caution since they are based upon territory densities from Thailand, where the results show the forest structure to differ significantly. It may be therefore that the average territory density in Myanmar falls outside the extremes recorded in Thailand. However, the range of territory densities of 10 other species of pitta given in Lambert & Woodcock (Reference Lambert and Woodcock1996) fell within or above the range of estimates used to estimate the population in Myanmar, suggesting that the estimates from Myanmar are unlikely to be overestimates.
A heavy reliance on earthworms in the diet of nestlings, and possibly adults, appears to be a common feature of the genus, as they were found to be the most important item in the diet of nestling Fairy Pittas Pitta nympha (Lin et al. Reference Lin, Yao and Lee2007b) and of adult and nestling Rainbow Pittas Pitta iris (Zimmerman & Noske Reference Zimmerman and Noske2003). The selection by Gurney's Pittas in southern Thailand of wetter than average gullies and the siting of nest sites close to water sources might reflect a reliance of damp habitats rich in earthworms. This suggests that reforestation efforts currently underway through the GPSRP should be focused on naturally damper areas.
Extremely heavy snake predation appears to lead to the loss of a very high proportion of nests in Thailand. The prevention of no fewer than 11 separate snake attacks at four guarded nests indicates the severity of this problem, and of only four nests thought to have produced chicks out of the 24 nests found during the study, three were heavily guarded by 24-hour wardening. Snakes are perhaps the most important predators of bird nests in the tropics (Weatherhead and Blouin-Demers Reference Weatherhead and Blouin-Demers2004) and their access to nests is facilitated by the interconnectivity of tree canopies (Koenig et al. Reference Koenig, Wunderle and Enkerlen-Hoeflich2007). The secondary nature of the forest used by Gurney's Pitta in southern Thailand means that canopy connectivity is very high, and the strong selection by nesting Gurney's Pittas in Thailand for palms of the genera Calamus and Salacca, which are protected by sharp spines, might reflect an attempt to reduce predation. The snake species recorded attacking nests in southern Thailand all belonged to the genus Boiga, which is known to be highly invasive and to cause high nest predation elsewhere (Wiles et al. Reference Wiles, Bart, Beck and Aguon2003). Although the species involved in attacks of Gurney's Pitta nests were all native, their densities might have been inflated by the highly fragmented nature of the forest patches occupied by the bird (e.g. Chalfoun et al. Reference Chalfoun, Ratnaswamy and Thompson2002). Tests using electrical barriers have proved successful in reducing snake predation of the nests of rare birds (Aguon et al. Reference Aguon, Campbell and Morton2002) and might be useful in the conservation of Gurney's Pitta in southern Thailand.
The apparent stabilisation of the population and its lowland forest habitat in Thailand suggest that recent conservation efforts have had some success in containing previous declines in both. An apparent reduction in the rate of loss of lowland forest coincides with efforts to reduce illegal forest clearance through improved forest protection patrols, awareness-raising in the local community and other actions set out in the GPSRP. A tree nursery has been established, local staff trained in forest restoration and reforestation plots established. Due to its secondary nature, it has been estimated that habitat suitable for Gurney's Pitta could be recreated from confiscated oil palm plantations within as little as 10–15 years (S. Elliott, pers comm.). However, the complex political and socioeconomic problems besetting efforts to save the Thai population of Gurney's Pitta, described by Gretton et al. (Reference Gretton, Kohler, Lansdown, Pankhurst, Parr and Robson1993) and BirdLife International (2001), have been reduced rather than removed, and Gurney's Pitta in Thailand can be regarded as conservation dependent. As well as being threatened by forest loss, Gurney's Pittas appear to be very susceptible to disturbance during the breeding season, leading the local authorities to close the trail system in the core area during the breeding period. Unfortunately, this ban is ignored by both local people and visiting birdwatchers. In Myanmar, the population may be considerably higher than the 5,000 to 8,500 territories estimated by Eames et al. (Reference Eames, Htin, Leimgruber, Kelly, Sein Myo, Moses and U Saw Nyunt2005) and forest loss in southern Tanintharyi was relatively low between 1990 and 2000 (Leimgruber et al. Reference Leimgruber, Kelly, Steininger, Brunner, Müller and Songer2005). As a result, the IUCN threat category for Gurney's Pitta might merit downlisting to ‘Vulnerable’ (under category B1) or even ‘Near Threatened’. However, forest loss in Myanmar is likely to accelerate through state policies to support oil palm plantations and level lowland forest cannot be regarded as safe anywhere in southeast Asia. Although the species appears able to tolerate a degree of forest disturbance, and might survive or even benefit from some forest disturbance, the main threat remains clear-felling for oil palm plantations. The future of this species in both Myanmar and Thailand will depend on adequate and sustainable forest protection and management.
Acknowledgements
This work was largely funded by the Darwin Initiative of the UK Government's Department of the Environment, Farming and Rural Affairs (DEFRA). Additional funding was provided by the British Bird Fair. For constructive comments on earlier drafts we thank Stuart Marsden, Anthony Sebastian, Phil Round and an anonymous referee. For help in many ways during this programme of research we thank Isorn Sawaddiparp, Pitak Kaeoploy, Phil Round, Kritsana Kaewplang, U Uga, Petch Manopawitr, Brian and Margaret Sykes, Budsabong Kanchanasaka, Gawin Chutima, Ian Barber, Yotin Meekaeo, Uthai Treesucon, Stephen Elliott, Steve Parr, John Parr, Debbie Pain, Dieter Hoffman, Tim Melling, Jeremy Lindsell, Gary Woodburn, Malcolm and Sheena Davies, Andrew Owen, Somchai Prempanitnukul, Wasun Klomjinda, Prasan Prempree and Somprat Phonchu.









