


Introduction
Nearly 300 species of landbirds, whose combined populations represent billions of birds, migrate between the Neotropics and North America (Martin and Finch Reference Martin and Finch1995, DeGraaf and Rappole Reference DeGraaf and Rappole1996, Berlanga et al. Reference Berlanga, Kennedy, Rich, Arizmendi, Beardmore and Blancher2010). As many as 50 of these species make an annual round trip of > 10,000 km but the number of individuals is declining year after year, especially among those species that migrate farthest (Sauer et al. Reference Sauer, Hines, Pardieck, Ziolkowski and Link2014). The same is apparent in Afro-Palearctic migrants (Sanderson et al. Reference Sanderson, Donald, Pain, Burfield and van Bommel2006). Global declines in migratory species have led to concerns that migration and the capacity of our ecosystems to maintain this process are in danger of disappearing (Wilcove and Wikelski Reference Wilcove and Wikelski2008, Bauer and Hoye Reference Bauer and Hoye2014). For many species, the majority of their long-distance migrations take place south of the Tropic of Cancer, yet our understanding of migration is strongly biased by studies in the temperate zone. Consequently the Neotropics continue to be described as the “black box” in our knowledge of migratory landbirds (Faaborg et al. Reference Faaborg, Holmes, Anders, Bildstein, Dugger, Gauthreaux, Heglund, Hobson, Jahn, Johnson, Latta, Levey and Marra2010a).
As early as the 1980s, results from the North American Breeding Bird Survey highlighted widespread declines in Nearctic-Neotropical migratory landbirds (Terborgh Reference Terborgh, Keast and Morton1980, Robbins et al. Reference Robbins, Sauer, Greenberg and Droege1989), and considerable efforts have been made since to both understand and address the year-round conservation needs of this group (Keast and Morton Reference Keast and Morton1980, Hagan and Johnston Reference Hagan and Johnston1992, Holmes Reference Holmes2007, Faaborg et al. Reference Faaborg, Holmes, Anders, Bildstein, Dugger, Gauthreaux, Heglund, Hobson, Jahn, Johnson, Latta, Levey and Marra2010a). Partners in Flight (PIF), for example, was created in 1990 to develop strategies for the conservation of migratory landbirds throughout their annual cycle, and today is the umbrella group for a diverse set of actors from across the Americas (Finch and Martin Reference Finch and Martin1995, Rich et al. Reference Rich, Beardmore, Berlanga, Blancher, Bradstreet and Butcher2004, Berlanga et al. Reference Berlanga, Kennedy, Rich, Arizmendi, Beardmore and Blancher2010, Rosenberg et al. Reference Rosenberg, Kennedy, Dettmers, Ford, Reynolds and Alexander2016). More than 30 years of research into Neotropical migratory birds has substantially advanced our understanding of how populations may be limited by events on the breeding grounds (Holmes et al. Reference Holmes, Marra and Sherry1996) or the wintering grounds (Latta and Faaborg Reference Latta and Faaborg2002, Studds and Marra Reference Studds and Marra2005, Johnson et al. Reference Johnson, Sherry, Holmes and Marra2006), and how effects may carry over from one season to the other (Nott et al. Reference Nott, Desante, Siegel and Pyle2002, Norris et al. Reference Norris, Marra, Kyser, Sherry and Ratcliffe2004, Norris and Marra Reference Norris and Marra2007, Harrison et al. Reference Harrison, Blount, Inger, Norris and Bearhop2011, González-Prieto and Hobson Reference González-Prieto and Hobson2013). Indeed, the formulation of the winter limitation hypothesis (Sherry and Holmes Reference Sherry, Holmes, Martin and Finch1995) and the subsequent collection of empirical evidence for the limiting role of winter habitats have firmly placed Neotropical wintering grounds on the research and conservation agenda (Marra Reference Marra2000, Latta and Faaborg Reference Latta and Faaborg2002, Studds and Marra Reference Studds and Marra2005). The same cannot be said, however, of the ecological needs and constraints of these same birds while migrating through the Neotropics (Faaborg et al. Reference Faaborg, Holmes, Anders, Bildstein, Dugger, Gauthreaux, Heglund, Hobson, Jahn, Johnson, Latta, Levey and Marra2010a), despite the considerable distances that many long-distance migrants must traverse there.
For some bird species, migration is by far the greatest source of mortality during their annual cycle (Sillett and Holmes Reference Sillett and Holmes2002, Newton Reference Newton2006, Rockwell et al. Reference Rockwell, Wunderle, Sillett, Bocetti, Ewert and Currie2016). For example, successive delayed arrivals or habitat degradation at even a single major stopover site can lead to significant declines, threatening the viability of populations across the Western Hemisphere (Baker et al. Reference Baker, González, Piersma, Niles, do Nascimento and Atkinson2004). Although some en-route flexibility exists (Tottrup et al. Reference Tottrup, Thorup, Rainio, Yosef, Lehikoinen and Rahbek2008, Stanley et al. Reference Stanley, MacPherson, Fraser, McKinnon and Stutchbury2012), migration strategies are under strong genetic control (Gwinner Reference Gwinner1996, Delmore and Irwin Reference Delmore and Irwin2014), and migrants are expected to be limited in their ability to change routes if conditions change or stopover habitats are no longer available in the short term but may be able to adapt in the longer term (Sutherland Reference Sutherland1998). Recognition of the critical importance of migration stopovers and bottlenecks has led to repeated calls for more studies and collaborations into the ecology and needs of birds during the migration periods (Hutto Reference Hutto1998, Heglund and Skagen Reference Heglund and Skagen2005, Mehlman et al. Reference Mehlman, Mabey, Ewert and Duncan2005, Faaborg et al. Reference Faaborg, Holmes, Anders, Bildstein, Dugger, Gauthreaux, Heglund, Hobson, Jahn, Johnson, Latta, Levey and Marra2010a, Reference Faaborg, Holmes, Anders, Bildstein, Dugger, Gauthreaux, Heglund, Hobson, Jahn, Johnson, Latta, Levey and Marra2010b).
These appeals have led to more work within North America, especially along the USA Gulf Coast (Buler et al. Reference Buler, Moore and Woltmann2007, Moore and Buler Reference Moore and Buler2011, Laughlin et al. Reference Laughlin, Taylor, Bradley, LeClair, Clark and Dawson2013, Cohen et al. Reference Cohen, Barrow, Buler, Deppe, Farnsworth, Marra, McWilliams, Mehlman, Wilson, Woodrey and Moore2017), the Pacific flyway (Carlisle et al. Reference Carlisle, Skagen, Kus, van Riper, Paxton and Kelly2009), riparian forests in the south-west (Yong and Finch Reference Yong and Finch1997, Yong et al. Reference Yong, Finch, Moore and Kelly1998) and around the Great Lakes (Dunn Reference Dunn2001, Reference Dunn2002, Ewert et al. Reference Ewert, Hamas, Smith, Dallman and Jorgensen2011). For many migratory species, however, the majority of their migratory route lies south of these well-studied areas. For example, millions of migrating landbirds converge each year on the northern Neotropics (La Sorte et al. Reference La Sorte, Fink, Hochachka and Kelling2016), and species whose breeding range may stretch over 4,000 km from east to west concentrate into areas spanning less than 100 km as a result of the funnel-shaped geography of Central America (Bildstein Reference Bildstein2004). Yet, despite this obvious importance, migration studies of Nearctic-Neotropical landbirds south of the United States of America (USA) and Canada remain rare, limiting our ability to design effective conservation strategies that address their needs throughout the annual cycle.
Nonetheless, the last few decades have seen studies from Mexico’s Gulf Coast and the Yucatan Peninsula (Winker Reference Winker1995, Deppe and Rotenberry Reference Deppe and Rotenberry2005, Johnson and Winker Reference Johnson and Winker2008, Bayly and Gómez Reference Bayly and Gómez2011, Shaw and Winker Reference Shaw and Winker2011), and from northern Colombia (Colorado Reference Colorado2010, Bayly et al. Reference Bayly, Gómez, Hobson, González and Rosenberg2012a, Reference Bayly, Gómez and Hobson2013, Gómez et al. Reference Gómez, Bayly and Rosenberg2013, Reference Gómez, Gómez-Bahamón, Cárdenas-Ortiz and Bayly2015), in some cases demonstrating the critical importance of stopover sites within the Neotropics (Bayly et al. Reference Bayly, Gómez, Hobson, González and Rosenberg2012a, Reference Bayly, Gómez and Hobson2013). Enormous gaps remain, however, and in particular our knowledge of how nocturnally migrating birds use the region south of the Yucatan Peninsula through Central America to northern South America; large Caribbean Islands; and especially regions within South America, is poor. In addition, for over 30 species that winter primarily in South America, much confusion persists around which regions are used during migration because of imprecisely mapped winter distributions and the difficulty of distinguishing migrating from wintering individuals (Remsen Reference Remsen2001). Some of these gaps are being partially filled as a result of innovative lightweight tracking technologies and their application to a growing number of Neotropical migratory landbirds (McKinnon et al. Reference McKinnon, Fraser and Stutchbury2013a, Taylor et al. Reference Taylor, Crewe, Mackenzie, Lepage, Aubry, Crysler, Finney, Francis, Guglielmo, Hamilton, Holberton, Loring, Mitchell, Norris, Paquet, Ronconi, Smetzer, Smith, Welch and Woodworth2017). The migration of raptors is better understood, with multiple studies detailing the main migration routes (Martell et al. Reference Martell, Henny, Nye and Solensky2001, Bildstein Reference Bildstein2004, Porras-Peñaranda et al. Reference Porras-Peñaranda, Robichaud and Branch2004, Colorado et al. Reference Colorado, Bechard, Márquez and Castaño2006, Ruelas Inzunza et al. Reference Ruelas Inzunza, Goodrich and Hoffman2010a, Kochert et al. Reference Kochert, Fuller, Schueck, Bond, Bechard and Woodbridge2011, Bayly et al. Reference Bayly, Cárdenas-Ortiz, Rubio and Gómez2014).
While general patterns of migration speed and routes have been shown to differ between spring and autumn migrations for many species (Gómez et al. Reference Gómez, Bayly and Rosenberg2013, La Sorte et al. Reference La Sorte, Fink, Hochachka, DeLong and Kelling2014a, Reference La Sorte, Fink, Hochachka, Farnsworth, Rodewald and Rosenberg2014b, Hobson and Kardynal Reference Hobson and Kardynal2015), detailed seasonal comparisons for most species and regions are lacking. In addition, the strategies and needs of diurnal and nocturnal migrating species are expected to differ markedly, making generalisations across species groups difficult. Given the critical importance of migration to annual survival and fitness (Sillett and Holmes Reference Sillett and Holmes2002, Finch et al. Reference Finch, Pearce-Higgins, Leech and Evans2014) and its ecological significance for long-term population viability, our lack of knowledge of stopover regions and migratory bottlenecks within the Neotropics poses a serious constraint on our ability to address declines in Nearctic-Neotropical migrants (Faaborg et al. Reference Faaborg, Holmes, Anders, Bildstein, Dugger, Gauthreaux, Heglund, Hobson, Jahn, Johnson, Latta, Levey and Marra2010a, Sheehy et al. Reference Sheehy, Taylor, McCann and Norris2010).
In this review we synthesise current knowledge of stopover regions and migratory bottlenecks within the Neotropics, focusing on diurnal and nocturnally migrating landbirds. Our primary objective is to stimulate research in regions and for species that lack even baseline data. We outline potential threats at known and suspected stopover regions, and present an emerging picture of the migration strategies of long-distance migratory birds in the Neotropics. We also propose consistent terminology that, for example, can make the distinction between “true” stopover—where birds engage in multi-day stops with significant refuelling—and rest-roost stops lasting one day or less with no or limited refuelling. Then, based on a conceptual framework for understanding the needs of migratory birds, we propose a targeted and collaborative research agenda at an expanded network of sites across the Neotropics within the context of new regional conservation planning strategies (e.g. Rosenberg et al. Reference Rosenberg, Kennedy, Dettmers, Ford, Reynolds and Alexander2016).
The needs of migrating landbirds in the Neotropics
To successfully migrate between their breeding and over-wintering grounds, where over-wintering is defined as the stationary period/s of the non-breeding season during the boreal winter (Table 1), Nearctic-Neotropical migrants depend on a series of sites along the length of their migratory route. These sites provide critical resources such as the fuel for migratory flights, safe roosting sites, and refuges during unfavourable climatic conditions (Mehlman et al. Reference Mehlman, Mabey, Ewert and Duncan2005, Newton Reference Newton2008). A successful migration also depends on birds avoiding in-flight hazards such as collisions with human infrastructure (Longcore et al. Reference Longcore, Rich, Mineau, MacDonald, Bert and Sullivan2013) and persecution by humans (Newton Reference Newton2006). Migrating landbirds stop at multiple sites across regions, yet not all sites are of equal importance for determining the speed of migration or survival (Weber et al. Reference Weber, Ens and Houston1998). Indeed, diurnal migrants typically stop on a nightly basis between daytime flights, while nocturnal migrants generally stop on a daily basis when migrating overland (Newton Reference Newton2008). As a consequence, migrants can be encountered at numerous sites along the length of their migratory routes, making it difficult to differentiate between sites used to rest between flights and those used to accumulate energy reserves (Warnock Reference Warnock2010).
Table 1. Stopover and migration terminology adopted throughout this review and its equivalence to the categories proposed by Mehlman et al. (Reference Mehlman, Mabey, Ewert and Duncan2005), which was used as the foundation.

The lack of distinction between these two very different types of “stopover sites” has given rise to a general perception that most migratory landbirds use numerous stopover sites during any one migration (McKinnon et al. Reference McKinnon, Fraser and Stutchbury2013a) and that population level movement across space is continuous. Because of this perception, research, conservation, and management activities for birds on migration have often focused on sites where large numbers of birds are present (Moore et al. Reference Moore, Kerlinger and Simmons1990, Simons et al. Reference Simons, Moore and Gauthreaux2000, Dunn Reference Dunn2001), without considering the ecological function of those sites. Recent studies on a range of species have challenged this paradigm, showing that in reality, individual landbirds often make long stopovers at just three or four key areas along the migratory route (Figure 1) (Heckscher et al. Reference Heckscher, Taylor, Fox and Afanasyev2011, Kochert et al. Reference Kochert, Fuller, Schueck, Bond, Bechard and Woodbridge2011, Delmore et al. Reference Delmore, Fox and Irwin2012, Fraser et al. Reference Fraser, Stutchbury, Silverio, Kramer, Barrow and Newstead2012, Stanley et al. Reference Stanley, MacPherson, Fraser, McKinnon and Stutchbury2012, Callo et al. Reference Callo, Morton and Stutchbury2013, McKinnon et al. Reference McKinnon, Fraser and Stutchbury2013a, Renfrew et al. Reference Renfrew, Kim, Perlut, Smith, Fox and Marra2013), at which they accumulate large energy reserves (Figure 2) (Bayly et al. Reference Bayly, Gómez, Hobson, González and Rosenberg2012a, Reference Bayly, Gómez and Hobson2013). These findings highlight an urgent need to identify major stopover regions (defined here as areas varying in size from 100 km2 to 50,000 km2 with relatively uniform environmental conditions and offering a certain set of resources) and assess the needs of birds within them. This need is magnified in the Neotropics where the funnel-shaped geography of Mexico and Central America acts as a bottleneck, concentrating millions of migratory landbirds (Porras-Peñaranda et al. Reference Porras-Peñaranda, Robichaud and Branch2004, Batista et al. Reference Batista, Miro, Angehr and Bildstein2005, Ruelas Inzunza et al. Reference Ruelas Inzunza, Goodrich and Hoffman2010a, La Sorte et al. Reference La Sorte, Fink, Hochachka and Kelling2016). This also highlights a need to revise the current stopover terminology to better define the contribution of individual sites to migration strategies, and to this end we present an updated terminology in Table 1 that we utilize throughout the review.
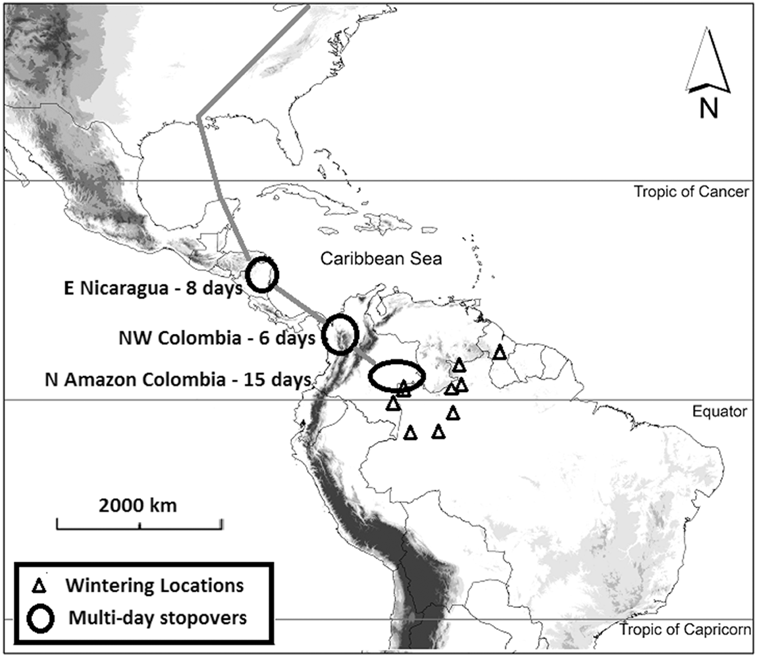
Figure 1. Spring migration strategy (northward) and stopover site use of Red-eyed Vireos from a breeding site in the north-eastern United States as revealed by geolocators recovered from nine individuals (adapted from Callo et al. Reference Callo, Morton and Stutchbury2013). Eight of the nine individuals appeared to make a rest-roost stop close to the tip of the Yucatan peninsula before crossing the Gulf of Mexico.
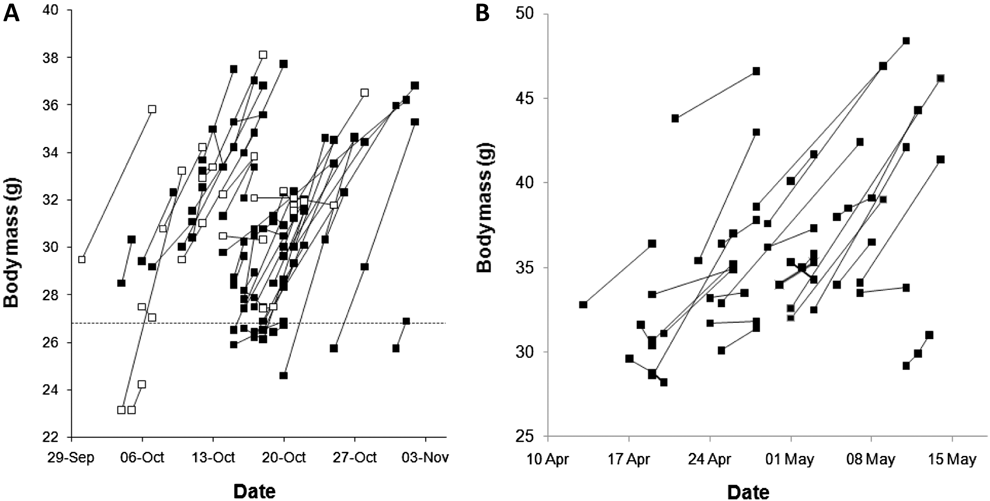
Figure 2. Individual A. Veery and B. Grey-cheeked Thrush recaptured on more than one occasion during an autumn and spring stopover respectively, in the Sierra Nevada de Santa Marta in northern Colombia. Birds increased significantly in body mass, providing evidence for extensive fuelling in this region (taken from Bayly et al. Reference Bayly, Gómez, Hobson, González and Rosenberg2012a , 2013).
One of the major factors driving the organisation, speed and, ultimately, the success of migration is the quality of resources at stopover sites and their impact on the rate of fuel deposition (Hedenström Reference Hedenström2008). Indeed, with time-minimiation emerging as the main currency shaping the evolution of migration strategies (Alerstam Reference Alerstam2011), migratory birds are expected to seek out sites that maximize their fuel deposition rate and, accordingly, their speed of migration (Buler et al. Reference Buler, Moore and Woltmann2007, La Sorte et al. Reference La Sorte, Fink, Hochachka, DeLong and Kelling2014a). It follows that one of the fundamental needs of migratory birds is therefore the availability of high quality habitats (those supporting high fuel deposition rates), in specific regions along their migratory route (Weber et al. Reference Weber, Houston and Ens1999, Bayly et al. Reference Bayly, Gómez, Hobson and Rosenberg2016). These habitats must not only sustain high rates of fuel deposition but also offer low to moderate predation risk, as migratory birds may avoid sites with elevated densities of predators or trade-off their foraging rate against predator surveillance (Lindström Reference Lindström1990, Schmaljohann and Dierschke Reference Schmaljohann and Dierschke2004, Pomeroy Reference Pomeroy2006, Warnock Reference Warnock2010). Recent work on Catharus thrushes in northern Colombia has shown that the energy reserves acquired at key stopovers may enable birds to cover 30% or more of their total migratory distance (Bayly et al. Reference Bayly, Gómez, Hobson, González and Rosenberg2012a, Bayly et al. Reference Bayly, Gómez and Hobson2013, Gómez et al. Reference Gómez, Bayly, Norris, Mackenzie, Rosenberg, Taylor, Hobson and Cadena2017), again highlighting the enormous influence that individual stopover sites can have on a migratory journey (Figure 2).
Additional sites used by migrating birds between major stopover regions (which we will refer to as rest-roost stops hereafter) are believed to have far less impact on the speed and organistion of migration (Alerstam Reference Alerstam2011); nevertheless they have the potential to affect survival (Newton Reference Newton2006). Energetic requirements at rest-roost stops are generally low, as individuals may only be resting between successive nocturnal or diurnal flights, although the ability to top up reserves may constitute a key element of the migratory strategy in certain species (Robson et al. Reference Robson, Barriocanal, Garcia and Villena2001, Delingat et al. Reference Delingat, Dierschke, Schmaljohann, Mendel, Bairlein and Delingat2006). The primary function of rest-roost stops is therefore to provide safe roosting conditions and access to resources such as water. Sites used as rest-roost stops may assume a greater importance when situated close to ecological barriers, such as the Gulf of Mexico or the Caribbean Sea (Moore et al. Reference Moore, Kerlinger and Simmons1990). Under optimal conditions, birds typically overfly coastal habitats either side of these barriers (Lowery Reference Lowery1945, Moore et al. Reference Moore, Kerlinger and Simmons1990) but when they encounter unfavourable conditions such as rain or strong headwinds, these habitats may temporarily support high concentrations of birds (Simons et al. Reference Simons, Moore and Gauthreaux2000). Habitats used in such emergencies, often referred to as “fire escapes”, can also be important away from ecological barriers (Mehlman et al. Reference Mehlman, Mabey, Ewert and Duncan2005). For example, passing tropical storms in Central America occasionally ground migrating raptors for days at sites where normally they would not stop (Bildstein Reference Bildstein2004) and possibly delay subsequent returns to the breeding grounds.
Just as stopover sites used for migratory fuelling affect the outcome of migration, the quality of sites occupied prior to the onset of migration can also have a fundamental impact on migration strategies (Bearhop et al. Reference Bearhop, Hilton, Votier and Waldron2004, Norris and Marra Reference Norris and Marra2007). For example, many species undergo pre-migratory fuelling on or near their Neotropical overwintering grounds, and the energy reserves gained there may enable birds to cover a significant proportion of their northbound migration (Stutchbury et al. Reference Stutchbury, Tarof, Done, Gow, Kramer and Tautin2009, Heckscher et al. Reference Heckscher, Taylor, Fox and Afanasyev2011, DeLuca et al. Reference DeLuca, Woodworth, Rimmer, Marra, Taylor and McFarland2015). Further, the quality of over-wintering habitats can influence the timing of migration, both in terms of initiation and arrival on the breeding grounds, with carry-over effects on territory acquisition and breeding performance (Norris et al. Reference Norris, Marra, Kyser, Sherry and Ratcliffe2004, Studds and Marra Reference Studds and Marra2005, Norris and Marra Reference Norris and Marra2007, Guillemain et al. Reference Guillemain, Elmberg, Arzel, Johnson and Simon2008). Overwintering habitats therefore likely constitute an essential element in many birds’ migration strategies but, unlike stopover habitats, they tend not to support high concentrations of birds. As a consequence, the loss of stopover habitat along migration routes may have more severe population consequences than the loss of overwinter habitat used for pre-migratory fuelling (Baker et al. Reference Baker, González, Piersma, Niles, do Nascimento and Atkinson2004, Newton Reference Newton2004).
While moult rarely overlaps with migration, this is not true in all species and stopover or pre-migratory fuelling sites may also be used to meet the energetic requirements of feather replacement. Moult-migration, whereby birds undergo a complete moult along their migration route, apparently “pausing” their migration to do so, occurs in the monsoon region of south-western USA and north-western Mexico in several species that breed in western North America (Leu and Thompson Reference Leu and Thompson2002, Rohwer et al. Reference Rohwer, Butler, Froehlich, Greenberg and Marra2005). Energetic demands at moult-migration sites are presumably higher than at typical stopover sites, yet the extent to which this phenomenon occurs elsewhere in the Neotropics is unknown. Similarly, pre-alternate moults that give rise to breeding plumages also likely occur at sites used for pre-migratory fuelling, but specific strategies or needs of birds during this critical phase of the life cycle have rarely been studied. In at least one long-distance migrant, the Bobolink Dolichonyx oryzivorus, areas used in South America for pre-alternate moult are part of a sequence of stationary non-breeding sites used by this species, blurring the concepts of over-wintering, stopover, or staging sites (Renfrew et al. Reference Renfrew, Kim, Perlut, Smith, Fox and Marra2013).
Current knowledge of stopover regions and migratory bottlenecks for landbirds in the Neotropics
In this section we discuss our current knowledge of diurnal migrants and nocturnal migrants separately, given the differing needs of these two groups; e.g. diurnal migrants both migrate and feed by day and sleep at night, whereas nocturnal migrants migrate at night and feed and/or rest by day. This may not hold true, however, when birds make long over-water flights that take more than 12 hours to complete. Secondly, we discuss findings from geolocators and other tracking technologies independently of on-the-ground field studies, in light of the qualitative differences in the data they provide. Finally, although we discuss migrants from both eastern and western North America, we concentrate on the preponderance of eastern breeding species whose migration routes take them through Central and northern South America and across barriers such as the Gulf of Mexico and Caribbean Sea. In contrast, many species from western North America overwinter in the northern extent of the Neotropics (Pacific slope of Mexico and northern Central America; Kelly and Hutto Reference Kelly and Hutto2005) and key stopover regions linked with survival and productivity occur outside of our region of interest (LaManna et al. Reference LaManna, George, Saracco, Nott and Desante2012, Drake et al. Reference Drake, Rock, Quinlan, Martin and Green2014). Nonetheless, a handful of western species migrate to South America and likely also rely on Neotropical stopovers.
The studies presented in this section were compiled following an ongoing and exhaustive literature search, employing the Google Scholar and Web of Science search engines. We entered the following key words singly or in combination: fuelling, fuel load, fuel deposition rate, geolocator, migration, migratory birds, migration strategy, stopover; in combination with geographical locations such as Neotropics, Central America, Costa Rica, Colombia, etc. In addition, the literature cited in relevant publications was surveyed for appropriate references. In compiling and summarising information, we specifically looked for papers in which evidence for stopovers such as stopover duration or changes in fuel loads were presented. Finally, we used the general Google search engine to discover technical reports published on the web.
Raptors and other diurnal migrants
Among the diurnally migrating species that use terrestrial habitats, perhaps the best-known group is the raptors (in which we include falcons and New World Vultures). In addition to long-term monitoring programmes at a variety of sites between Veracruz, Mexico (Ruelas Inzunza et al. Reference Ruelas Inzunza, Goodrich and Hoffman2010a), and the Canal Zone in Panama (Porras-Peñaranda et al. Reference Porras-Peñaranda, Robichaud and Branch2004, Batista et al. Reference Batista, Miro, Angehr and Bildstein2005), the use of satellite transmitters has revealed considerable detail with respect to routes and stopovers (Fuller et al. Reference Fuller, Seegar and Schueck1998, Martell et al. Reference Martell, Henny, Nye and Solensky2001, Kochert et al. Reference Kochert, Fuller, Schueck, Bond, Bechard and Woodbridge2011).
More than 30 raptor species have been recorded migrating through Mesoamerica but the majority of the 4 to 6 million raptors that migrate to or through the Neotropics on an annual basis belong to just four species: Turkey Vulture Cathartes aura (2 million), Broad-winged Hawk Buteo platypterus (2.1 million), Swainson’s Hawk Buteo swainsonii (1.2 million), and Mississippi Kite Ictinia mississipiensis (0.2 million) (Bildstein Reference Bildstein2004, Ruelas Inzunza et al. Reference Ruelas Inzunza, Goodrich and Hoffman2010b). Other less abundant but visible species along the flyway include Osprey Pandion haliaetus, Peregrine Falcon Falco peregrinus, and Swallow-tailed Kite Elanoides forficatus. Acting like a natural bottleneck, the narrowing geography of Central America concentrates migrating raptors at several points. The most well studied bottlenecks include the narrow gap between the Sierra Madre Oriental and the Gulf of Mexico in Veracruz, Mexico (Ruelas Inzunza et al. Reference Ruelas Inzunza, Goodrich, Hoffman, Martínez, Smith and Peresbarbosa2009), the corridor between the Cordillera de Talamanca and the Caribbean Sea in Costa Rica and western Panama (Porras-Peñaranda et al. Reference Porras-Peñaranda, Robichaud and Branch2004) and the Isthmus of Panama (Batista et al. Reference Batista, Miro, Angehr and Bildstein2005). Nevertheless, large numbers of migrating raptors can be seen at several other points along the route including the Darién of Panama and Colombia (Bayly et al. Reference Bayly, Cárdenas-Ortiz, Rubio and Gómez2014), and further study will likely reveal a concentrated passage along the Pacific coasts of Guatemala, El Salvador and Nicaragua (Bildstein Reference Bildstein2004).
The routes taken by raptors between Mexico and South America are well-defined, with the main flyway dividing in two in southern Mexico, after birds have passed through coastal Veracruz (Bildstein Reference Bildstein2004). The majority of birds cross to the Pacific coast at this point, following it down through Guatemala, El Salvador and Nicaragua until crossing to the Caribbean lowlands in southern Nicaragua and northern Costa Rica (Porras-Peñaranda et al. Reference Porras-Peñaranda, Robichaud and Branch2004, Kochert et al. Reference Kochert, Fuller, Schueck, Bond, Bechard and Woodbridge2011); however, Broad-winged Hawk, for example, may follow the Caribbean coast throughout. On entering Panama, many individuals cross again to the Pacific slope, although many others continue down the Caribbean coast, entering South America via the Darién. In South America routes are not well studied (Colorado et al. Reference Colorado, Bechard, Márquez and Castaño2006), but satellite tracks suggest that while Swainson’s Hawks continue on an almost direct southerly route (Kochert et al. Reference Kochert, Fuller, Schueck, Bond, Bechard and Woodbridge2011), Turkey Vultures head east-south-east towards the Llanos of Venezuela and Colombia. Many Osprey, Peregrine Falcon and Merlin Falco columbarius do not join the main flight through Mesoamerica until its lower portion, having left North America via the Florida peninsula and migrating over-sea via Cuba (Fuller et al. Reference Fuller, Seegar and Schueck1998, Martell et al. Reference Martell, Henny, Nye and Solensky2001, Rodríguez-Santana et al. Reference Rodríguez-Santana, Vega, Padilla, Plasencia León, Adán and Losada2014). Western Cuba has also been shown to be a major migratory route for USA-breeding Swallow-tailed Kites (Zimmerman and Meyer Reference Zimmerman and Meyer2004).
Migrating raptors generally pass along the routes described above rapidly, rarely pausing to top up fuel reserves (Smith et al. Reference Smith, Goldstein and Bartholomew1986, Fuller et al. Reference Fuller, Seegar and Schueck1998, Martell et al. Reference Martell, Henny, Nye and Solensky2001, Kochert et al. Reference Kochert, Fuller, Schueck, Bond, Bechard and Woodbridge2011); however, a number of stopover regions have been identified. For example, while autumn migrating Swainson’s Hawks appear to accumulate much of their energy requirements at stopovers in the southern USA, birds do make occasional stops in south-eastern Mexico, western Guatemala and Nicaragua, north-western Colombia and in the Colombian Llanos (Kochert et al. Reference Kochert, Fuller, Schueck, Bond, Bechard and Woodbridge2011). In contrast, the spring migration of Swainson’s Hawks does not appear to involve Neotropical stopovers (Smith et al. Reference Smith, Goldstein and Bartholomew1986, Kochert et al. Reference Kochert, Fuller, Schueck, Bond, Bechard and Woodbridge2011). Swallow-tailed Kites make a prolonged stopover on the Yucatan peninsula during autumn migration (Zimmerman and Meyer Reference Zimmerman and Meyer2004), seemingly to recover after an over-sea crossing from Florida or Cuba. The lack of information on other species and the general notion that most raptors can complete their migration through the Mesoamerican corridor without stopping to fuel (Smith et al. Reference Smith, Goldstein and Bartholomew1986), imply that their principal need may be safe roosting sites. However, anecdotal evidence aside (Hicks et al. Reference Hicks, Rogers and Child1966, Bildstein and Saborio 2000, Ruelas Inzunza et al. Reference Ruelas Inzunza, Goodrich, Hoffman, Martínez, Smith and Peresbarbosa2009), there is very little information regarding the use of roosting sites by migrating raptors. Nonetheless, the highly concentrated flyway used by most raptors suggests that they likely have as yet unidentified needs, which in part may be dictated by their diet which impedes the en masse use of stopover sites.
Besides raptors, very little has been published on other diurnally migrating landbirds, although these species are routinely encountered during raptor migration monitoring. Hirundines are a highly abundant group of diurnal migrants and seasonal counts of Cliff Petrochelidon pyrrhonota, Barn Hirundo rustica and Bank Riparia riparia Swallows at bottlenecks in Veracruz and the Colombian Darién involve hundreds of thousands of birds (Winkler Reference Winkler2006, Bayly et al. Reference Bayly, Cárdenas-Ortiz, Rubio and Gómez2014). Little is known, however, about stopover behaviour in these species. Given the declining status of many aerial insectivores (Table 2) (Nebel et al. Reference Nebel, Mills, McCracken and Taylor2010, Rosenberg et al. Reference Rosenberg, Kennedy, Dettmers, Ford, Reynolds and Alexander2016), determining stopover and major roost sites for this group is a priority, especially if they adopt staging behaviours similar to those seen in Purple Martin Progne subis and Tree Swallow Tachycineta bicolor (Fraser et al. Reference Fraser, Stutchbury, Kramer, Silverio, Barrow and Newstead2013, Laughlin et al. Reference Laughlin, Taylor, Bradley, LeClair, Clark and Dawson2013), where birds remain at sites longer than would appear necessary for fuelling purposes. Many aerial insectivores migrate as far as the southern cone of South America and likely have complex migration strategies and differential strategies between populations of a single species (Table 2). Other abundant yet rarely reported diurnal migrants are even less well known, and aside from counts at concentration points in the Darién (Bayly et al. Reference Bayly, Cárdenas-Ortiz, Rubio and Gómez2014), little is known of migration or even overwintering grounds of species such as Chimney Swift Chaetura pelagica, Black Swift Cypseloides niger, Common Nighthawk Chordeiles minor, Eastern Kingbird Tyrannus tyrannus and Dickcissel Spiza americana.
Table 2. Over-wintering destination and population status of diurnally migrating landbirds that winter in South America. Season counts are based on the highest single autumn counts from a single site in the Darién of Colombia (see Figure 4, region 3) reported in Bayly et al. (Reference Bayly, Cárdenas-Ortiz, Rubio and Gómez2014) or on counts from the same study site in 2014 and 2015. Three species (marked N/A) are rarely recorded at this watchsite presumably because their migration route takes them over the Caribbean Sea or the Pacific Ocean.
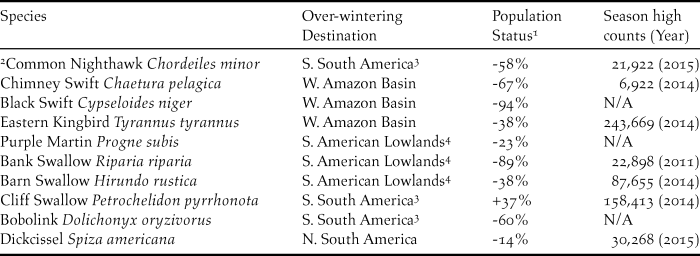
1 Population loss from 1970-2014 based on Rosenberg et al. Reference Rosenberg, Kennedy, Dettmers, Ford, Reynolds and Alexander2016.
2 Nighthawks regularly initiate and terminate flights during daylight hours but may migrate primarily at night.
3 Includes regions primarily south and east of the Amazon Basin.
4 Includes various lowland areas including Amazon Basin and regions to south and east.
Nocturnal migrants
The vast majority of Nearctic-Neotropical migratory landbirds (e.g. warblers, thrushes, tanagers, cuckoos) migrate solely or primarily at night (DeGraaf and Rappole Reference DeGraaf and Rappole1996, Alerstam Reference Alerstam2009), and many traverse vast stretches of the Caribbean basin or Central and northern South America to reach wintering grounds in the Andes, the Amazon Basin, or farther south still. Despite the importance of this extensive geography to annual survival, many Neotropical regions are still completely unexplored in terms of where, when and why migratory landbirds stop (Faaborg et al. Reference Faaborg, Holmes, Anders, Bildstein, Dugger, Gauthreaux, Heglund, Hobson, Jahn, Johnson, Latta, Levey and Marra2010b). In this section, we describe current knowledge of Neotropical stopover sites for nocturnal migrants within six broad geographic regions (Fig. 3): (1) south-eastern Mexico and the Yucatan peninsula; (2) northern Central America including Honduras, Nicaragua, and Guatemala; (3) Costa Rica and Panama; (4) northern Colombia; (5) the Caribbean islands; and (6) western Mexico. Other regions in the Neotropics are no doubt used for stopover but they remain largely unstudied.

Figure 3. The Neotropical region lying between the Tropic of Cancer and the Tropic of Capricorn. Major regions discussed in the section on nocturnal migrants and referred to throughout the text are outlined in shades of grey and named following the text.
South-eastern Mexico and the Yucatan Peninsula — Perhaps the best studied region, with several studies from Veracruz (Winker Reference Winker1995, Martínez Leyva et al. Reference Martínez Leyva, Ruelas Inzunza, Cruz Carretero, Barr, Peresbarbosa Rojas and Chávez Domínguez2005, Shaw and Winker Reference Shaw and Winker2011), the northern tip of the Yucatan (Deppe and Rotenberry Reference Deppe and Rotenberry2005), north-eastern Belize (Bayly and Gómez Reference Bayly and Gómez2011, Gómez and Bayly Reference Gómez and Bayly2011), and southern Belize (Johnson and Winker Reference Johnson and Winker2008). During autumn migration, species richness (≈70 species) and abundance of migratory landbirds is high and it is evident that the coastal geography of the region concentrates individuals of many species that cross the Gulf of Mexico or migrate around its western edge (Winker Reference Winker1995, Bayly and Gómez Reference Bayly and Gómez2011). However, mist-netting has revealed that individuals of most species carry moderate fuel reserves and therefore have little need to refuel at stopover sites (Winker Reference Winker1995, Johnson and Winker Reference Johnson and Winker2008, Bayly and Gómez Reference Bayly and Gómez2011). Low recapture probabilities and short between-capture durations also imply that most birds were making rest-roost stops between nocturnal flights (Bayly and Gómez Reference Bayly and Gómez2011).
Winker (Reference Winker1995) and Johnson and Winker (Reference Johnson and Winker2008) provide evidence that birds may be able to top up their reserves during these daytime stops but provide no evidence that birds stopover for longer. Together these findings suggest that most migrants (19 out of 20 transient species in north-east Belize, for example) passing through the region may accumulate sufficient energy reserves north of the Gulf of Mexico to cross both the Gulf and the Yucatan Peninsula (Bayly and Gómez Reference Bayly and Gómez2011). Not all species or individuals follow this pattern and Willow Flycatchers Empidonax traillii, for example, probably make a short refuelling stop before continuing their migration (Gómez and Bayly Reference Gómez and Bayly2011).
Spring migration sees a reduction in species richness in the region, and many transient species on route from South America, appear to be carrying sufficient reserves on arrival to reach North America without refuelling (Bayly and Gómez Reference Bayly and Gómez2011). For a smaller number of Central American wintering species, fuel loads, recapture rates and fuel deposition rates point to a stopover both on the Yucatan Peninsula and in south-eastern Mexico, including species of concern such as Wood Thrush Hylocichla mustelina and Kentucky Warbler Geothlypis formosa (Rogers and Odum Reference Rogers and Odum1966, Bayly and Gómez Reference Bayly and Gómez2011, Shaw and Winker Reference Shaw and Winker2011).
Northern Central America — There are no published studies of autumn stopover site use in northern Central America, despite the potential importance of this region for a large number of species that cross the Gulf of Mexico (Cohen et al. Reference Cohen, Barrow, Buler, Deppe, Farnsworth, Marra, McWilliams, Mehlman, Wilson, Woodrey and Moore2017). A spring study focused on Cerulean Warblers Setophaga cerulea in the mountains of northern Honduras, eastern Guatemala and southern Belize, highlighted the importance of this area for this species of conservation concern (Welton et al. Reference Welton, Anderson, Colorado, Hamel and Calderón-F2012). Cerulean Warblers are rarely recorded anywhere along their migration route between North American breeding grounds and Andean wintering grounds, but Welton et al. (Reference Welton, Anderson, Colorado, Hamel and Calderón-F2012) documented the presence of birds during a two-week period in early April, largely on Caribbean-facing mountains in Honduras and Guatemala. Several other warbler species, including the ‘Near Threatened’ Golden-winged Warbler Vermivora chrysoptera, were also recorded in this study. Given its geographic position and intriguing unpublished records submitted to the online public database eBird (e.g. 400 Bay-breasted Warblers, Setophaga castanea, counted at a single location in Nicaragua in April) (Sullivan et al. Reference Sullivan, Aycrigg, Barry, Bonney, Bruns, Cooper, Damoulas, Dhondt, Diettrich, Farnsworth, Fink, Fitzpatrick, Fredricks, Gerbracht, Gomes, Hochachka, Iliff, Lagoze, La Sorte, Merrifield, Morris, Phillips, Reynolds, Rodewald, Rosenberg, Trautmann, Wiggins, Winkler, Wong, Wood, Yu and Kelling2014), northern Central America will likely prove to be a critical stopover region for many species.
Costa Rica and Panama — Stopover studies from Costa Rica and Panama are also surprisingly rare, and aside from those dating back 50 years (Rogers and Odum Reference Rogers and Odum1966) and a recent study on Swainson’s Thrush Catharus ustulatus (Wilson et al. Reference Wilson, Hobson, Collister and Wilson2008), we could find no reference to the stopover behaviour of Nearctic-Neotropical migrants in this region. There is, however, a long-term migration monitoring station at Tortuguero, Costa Rica (Elizondo-Camacho and Ralph Reference Elizondo-Camacho and Ralph2012), and the data collected there would no doubt shed light on the use of the Caribbean lowlands for stopovers, beyond the habitat selection inferences thus far reported (Wolfe et al. Reference Wolfe, Johnson and Ralph2014). Further, unpublished records in eBird and elsewhere suggest that a variety of species (e.g. Cerulean Warbler) may depend on montane forest habitats in Costa Rica and Panama, especially in early autumn. Studies of autumn migrants on the Caribbean coast of Panama found a mixture of exhausted birds and individuals with sufficient reserves to continue their migration well into South America (Rogers Reference Rogers1965, Rogers and Odum Reference Rogers and Odum1966). There was some evidence that fuel-depleted birds stopped over in Panama to rebuild their energy reserves (Galindo et al. Reference Galindo, Méndez and Adames1963). In a spring study of Swainson’s Thrushes on the Pacific coast of Costa Rica, there was no evidence of birds making prolonged stopovers (Wilson et al. Reference Wilson, Hobson, Collister and Wilson2008).
Northern Colombia — In contrast to the above regions, a growing body of evidence on stopover site use in northern Colombia is emerging, with studies on Empidonax flycatchers in the Darién (Colorado Reference Colorado2010) and on Catharus thrushes, vireos, and warblers in the Sierra Nevada de Santa Marta (SNSM) and the Darién revealing new insights on the importance of this region to migrants (Gómez et al. Reference Gómez, Bayly and Rosenberg2013, Reference Gómez, Bayly and Rosenberg2014). During autumn, Empidonax flycatchers stopping in the Darién did not appear to build energy reserves (Colorado Reference Colorado2010), whereas three species of Catharus thrushes exhibited a mixed strategy, with some individuals passing through rapidly and others making multi-day stopovers to refuel (Gomez et al. 2014). In contrast, migrants passing through the SNSM frequently made prolonged stopovers in both autumn and spring. For example, Veery Catharus fuscescens on stopover in the SNSM stayed for an average of nine days in autumn, during which time they increased their energy reserves by around 30% (Figure 2) and were subsequently capable of flights of up to 2,000 km towards their South American overwintering grounds (Bayly et al. Reference Bayly, Gómez, Hobson, González and Rosenberg2012a). A similar situation was found for Grey-cheeked Thrush Catharus minimus in spring, with birds stopping in the SNSM for up to two weeks and storing sufficient energy to reach North America (>2,500 km) without refuelling (Bayly et al. Reference Bayly, Gómez and Hobson2013). In a third species, the Tennessee Warbler (Oreothlypis peregrina), long spring stopovers in the SNSM resulted in smaller fuel loads compared to Grey-checked Thrushes, but they were still sufficient for flights to northern Central America and, in some individuals, to the southern USA (Bayly et al. Reference Bayly, Gómez, Hobson and Rosenberg2016).
These recent results from studies in northern Colombia suggest that migratory landbirds are capable of adopting a migratory strategy similar to that of several shorebirds (Gómez et al. Reference Gómez, Bayly, Norris, Mackenzie, Rosenberg, Taylor, Hobson and Cadena2017); i.e. using a small number of sites to store large fuel reserves for long-distance flights (Piersma Reference Piersma1987, Gill et al. Reference Gill, Tibbitts, Douglas, Handel, Mulcahy and Gottschalck2009, Klaassen et al. Reference Klaassen, Alerstam, Carlsson, Fox and Lindström2011). The strategy of species like the Grey-cheeked Thrush also has similarities with those adopted by migrating raptors, which pass over Mesoamerica without the need for refuelling stops (Smith et al. Reference Smith, Goldstein and Bartholomew1986, Kochert et al. Reference Kochert, Fuller, Schueck, Bond, Bechard and Woodbridge2011). These strategies do not appear to be restricted to the species studied in Colombia to date, as the results from geolocators described below demonstrate. Another key lesson from studies in Colombia is that migratory landbirds are far more abundant in certain stopover habitats (e.g. pre-montane forest) compared to others and that different habitats support different fuelling opportunities and rates, which can make the difference between individuals successfully crossing a water barrier or needing to take a longer route or make additional stopovers (Gómez et al. Reference Gómez, Bayly and Rosenberg2013, Reference Gómez, Gómez-Bahamón, Cárdenas-Ortiz and Bayly2015, Bayly et al. Reference Bayly, Gómez, Hobson and Rosenberg2016).
Caribbean islands — Despite many long-term studies of overwintering migrants in the West Indies, studies of stopover behaviour in the Caribbean are rare. General texts on Caribbean birds indicate that most species migrating to South America are rarely encountered on Caribbean islands and that most records are thought to be weather-related (Garrido and Kirkconnell Reference Garrido and Kirkconnell2000, Raffaele et al. Reference Raffaele, Wiley, Garrido, Keith and Raffaele2003, Latta et al. Reference Latta, Rimmer, Keith, Wiley, Raffaele and McFarland2006); thus the optimal strategy seems to be to overfly the Caribbean, rather than stopping to refuel on islands. This hypothesis is supported by a study of Blackpoll Warbler Setophaga striata in the Dominican Republic, where birds primarily undertook daytime rest-roost stops, often in association with unfavourable weather (Latta and Brown Reference Latta and Brown1999). Geolocators suggest a similar role for Caribbean islands (see below), but further study may reveal stopover areas, particularly on large islands such as Cuba where information is scarce.
Western Mexico — Few studies of landbird migration stopover have occurred in western Mexico, yet the diversity of habitats, ranging from coastal lowlands to high mountains, combined with the large number of species migrating through or overwintering in Mexico, strongly suggests its importance as a stopover region, especially for species breeding in the western USA and Canada (Howell and Webb Reference Howell and Webb1995). Recent studies of moult migration in north-western Mexico, in which individuals of many species undergo an extended stopover to complete feather moult during and after the summer monsoon season and then continue to overwintering areas (Leu and Thompson Reference Leu and Thompson2002, Rohwer et al. Reference Rohwer, Butler, Froehlich, Greenberg and Marra2005, Pyle et al. Reference Pyle, Leitner, Lozano-Angulo, Avilez-Teran, Swanson and Gómez Limón2009), is one key example showing the importance of western Mexico to a critical life history stage.
Findings from geolocators and other tracking technologies
Geolocators have revolutionised our ability to track small landbirds during migration, revealing a wealth of information that can be used to identify migratory routes and stopover regions. Nonetheless, due to the relatively low precision of light-level geolocators (typically ± 100 km but up to 400 km in the tropics) (McKinnon et al. Reference McKinnon, Fraser and Stutchbury2013a, Reference McKinnon, Stanley, Fraser, MacPherson, Casbourn and Marra2013b), it is only by combining findings from geolocators with detailed on-the-ground field studies that habitat use and quality within broad stopover regions can be assessed. Indeed, the array of habitats present within polygons or confidence ellipses identified by geolocators can be extremely large. For example, in northern Colombia, where geolocators have identified a general region used by Veery for autumn stopover (Heckscher et al. Reference Heckscher, Taylor, Fox and Afanasyev2011, Hobson and Kardynal Reference Hobson and Kardynal2015), it is possible to travel from snow-capped peaks, through humid montane forest, to xeric scrub in less than 100 km. Only through detailed mark-recapture studies within that region has the high value of foothill and pre-montane forest been demonstrated for Veery and other Nearctic-Neotropical migrants (Bayly et al. Reference Bayly, Gómez, Hobson, González and Rosenberg2012a, 2016).
Geolocators have revealed a number of unexpected patterns in the migratory strategies of small landbirds, including faster than expected travel speeds and a general pattern of birds making prolonged stopovers in a small number of regions (McKinnon et al. Reference McKinnon, Fraser and Stutchbury2013a). For example, eastern-breeding Veery were found to make multi-day stopovers in three main regions, the south-eastern USA, northern Colombia, and the northern Amazon, when on route from a Delaware breeding site to overwintering sites in the southern Amazon basin (Heckscher et al. Reference Heckscher, Taylor, Fox and Afanasyev2011). Remarkably, western-breeding Veery also converge on the same northern Colombia stopover region in autumn (Hobson and Kardynal Reference Hobson and Kardynal2015), as results from an analysis of stable isotopes in feathers had predicted (González-Prieto et al. Reference González-Prieto, Hobson, Bayly and Gómez2011). Bobolink use fewer regions still, with many birds making one long stopover in the Llanos (grasslands) of Venezuela and Colombia, while on route between North America and their southern South American overwintering grounds (Renfrew et al. Reference Renfrew, Kim, Perlut, Smith, Fox and Marra2013).
The pattern of long stopovers at a relatively small number of sites is emerging from landbird studies in both the Americas and Europe (Stanley et al. Reference Stanley, MacPherson, Fraser, McKinnon and Stutchbury2012, Kristensen et al. Reference Kristensen, Tøttrup and Thorup2013, Lemke et al. Reference Lemke, Tarka, Klaassen, Akesson, Bensch and Hasselquist2013, DeLuca et al. Reference DeLuca, Woodworth, Rimmer, Marra, Taylor and McFarland2015). This strategy may be used primarily by nocturnally migrating species, as aerial insectivores (Beason et al. Reference Beason, Gunn, Potter, Sparks and Fox2012) and other diurnal migrant birds appear to behave differently (Jahn et al. Reference Jahn, Cueto, Fox, Husak, Kim and Landoll2013): although Fraser et al. (Reference Fraser, Stutchbury, Kramer, Silverio, Barrow and Newstead2013) documented prolonged stopovers in Purple Martin. Presumably, the ability of diurnal migrants to feed while actively migrating explains part of this difference. Aside from revealing the widespread occurrence of long stopovers, geolocators also show why landbirds were previously considered to use numerous sites/regions during migration. Indeed, although birds move rapidly between these major stopover regions, often at rates of 500 km/day (Callo et al. Reference Callo, Morton and Stutchbury2013, Fraser et al. Reference Fraser, Stutchbury, Kramer, Silverio, Barrow and Newstead2013), flights are interrupted at a series of sites along the route, which most likely act as diurnal or nocturnal rest-roost stops in order to avoid migrating at unfavourable times of day (Stutchbury et al. Reference Stutchbury, Tarof, Done, Gow, Kramer and Tautin2009, Delmore et al. Reference Delmore, Fox and Irwin2012).
There are still too few published geolocator studies to draw conclusions about the Neotropical stopover regions used by the wider community of migratory landbirds, or by species with shared wintering grounds, or even those from different breeding regions (e.g. eastern vs. western North America). Nevertheless, several regions are now known to be important for two or more species (Table 3). For example, Purple Martin (Fraser et al. Reference Fraser, Stutchbury, Kramer, Silverio, Barrow and Newstead2013), Swainson’s Thrush (Delmore et al. Reference Delmore, Fox and Irwin2012), and to a lesser extent Wood Thrush (Stutchbury et al. Reference Stutchbury, Tarof, Done, Gow, Kramer and Tautin2009) make an extended autumn stopover on the Yucatan Peninsula, joining the list of species found to make short stops there by banding studies (see above). Northern Colombia hosts extended stopovers by at least three species, Veery (autumn), Red-eyed Vireo Vireo olivaceus (spring), and Swainson’s Thrush (autumn), adding further evidence for this region’s importance to South American wintering species (Heckscher et al. Reference Heckscher, Taylor, Fox and Afanasyev2011, Delmore et al. Reference Delmore, Fox and Irwin2012, Callo et al. Reference Callo, Morton and Stutchbury2013). Geolocators further indicate that both Red-eyed Vireo (spring) and Swainson’s Thrush (autumn and spring) also make long stopovers in northern Central America, confirming our prediction that this region should figure prominently in migration strategies. Finally, it is worth emphasizing the critical importance of the Llanos of Venezuela and Colombia as an autumn and spring stopover site for Bobolink (Renfrew et al. Reference Renfrew, Kim, Perlut, Smith, Fox and Marra2013).
Table 3. Evidence for major stopover regions and bottlenecks for migratory landbirds in the Neotropics. Numbers following region names relate to Fig. 3 & 4.
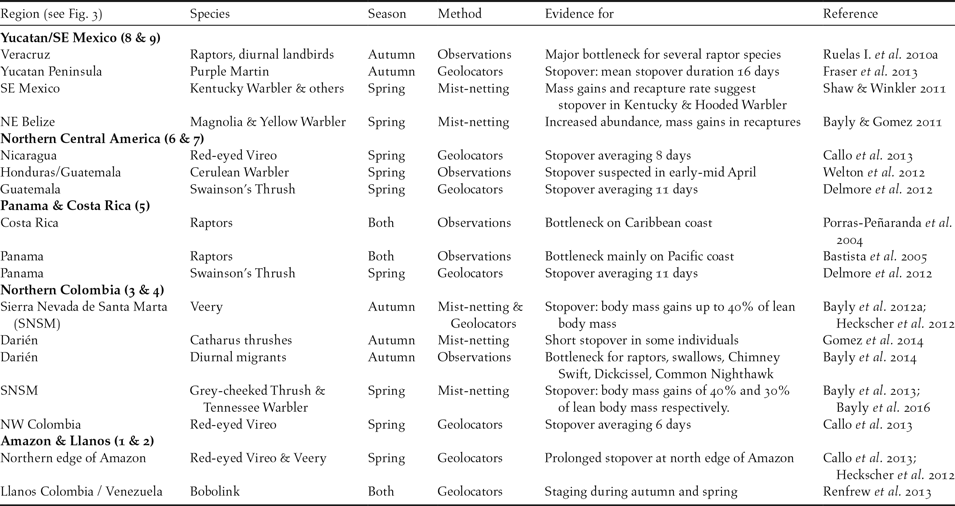
GPS tags and coded nano-tags (radio transmitters) also hold promise for determining the precise location of new stopover sites. Nano-tags, in particular, are an excellent tool for studying stopover behaviour and the outcome of subsequent migratory flights (Taylor et al. Reference Taylor, Mackenzie, Thurber, Calvert, Mills and McGuire2011, Reference Taylor, Crewe, Mackenzie, Lepage, Aubry, Crysler, Finney, Francis, Guglielmo, Hamilton, Holberton, Loring, Mitchell, Norris, Paquet, Ronconi, Smetzer, Smith, Welch and Woodworth2017), such as the confirmation of non-stop flights >3,000 km by Grey-cheeked Thrush on leaving a stopover site in Northern Colombia (Gómez et al. Reference Gómez, Bayly, Norris, Mackenzie, Rosenberg, Taylor, Hobson and Cadena2017).
Emerging picture of stopover regions in the Neotropics
The general pattern emerging from both on-the-ground field studies and the deployment of geolocators is that long-distance migration strategies in many small landbirds are actually similar to those of certain shorebirds (Atkinson et al. Reference Atkinson, Baker, Bennett, Clark, Clark and Cole2007, Gill et al. Reference Gill, Tibbitts, Douglas, Handel, Mulcahy and Gottschalck2009, Lindström et al. Reference Lindström, Alerstam, Bahlenberg, Ekblom, Fox and Råghall2016), involving long flights and prolonged multi-day stopovers at a relatively small number of sites. We are just beginning to understand the generality of this pattern, as well as which regions are of greatest importance to the largest number of migratory birds.
Along the major flyways used by species breeding in eastern North America, a number of major stopover regions are emerging (Table 3; Figure. 4), including areas inland of the Gulf Coast and northern Florida in North America and northern Colombia in South America, both of which are used to store sufficient energy to make unbroken flights across the Gulf of Mexico and/or the Caribbean Sea. In particular, the Sierra Nevada de Santa Marta, an isolated massif on Colombia’s Caribbean coast, is of considerable strategic importance for a number of species (Bayly et al. Reference Bayly, Gómez, Hobson, González and Rosenberg2012a, Reference Bayly, Gómez, Hobson and Rosenberg2016, Gómez et al. Reference Gómez, Gómez-Bahamón, Cárdenas-Ortiz and Bayly2015). Not all birds are capable of such long-distance flights and various points in Central America, such as the Yucatan/south-east Mexico and northern Central America host stopover regions for certain species but we lack sufficient information to highlight regions used by multiple species. No doubt the deployment of geolocators and GPS tags on additional species and the establishment of migration banding stations in little-studied regions such as eastern Nicaragua and Honduras will reveal other critical fuelling regions.
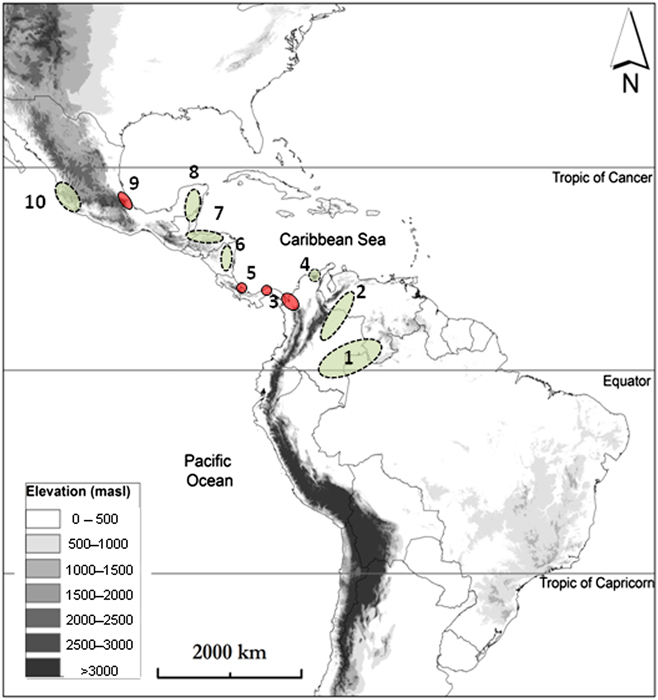
Figure 4. Major Neotropical stopover regions (dashed circles) and bottlenecks (closed circles) for migratory landbirds identified to date. 1. Northern Amazon (Veery, Red-eyed Vireo); 2. Llanos of Venezuela and Colombia (Bobolink); 3. NW Colombia (bottleneck; also stopovers by Red-eyed Vireo & Catharus thrushes); 4. Sierra Nevada de Santa Marta (Tennessee Warbler, Grey-checked Thrush, Veery); 5. Costa Rica Caribbean slope & Panama Canal Zone (bottleneck for raptors); 6. Western Nicaragua (Red-eyed Vireo); 7. Highlands Honduras, Guatemala and Belize (Cerulean Warbler); 8. Yucatan Peninsula (Purple Martin, Red-eyed Vireo, Magnolia Warbler); 9. Veracruz (bottleneck for raptors, Kentucky and Hooded Warblers, Scissor-tailed Flycatcher); 10. SW Mexico (Swainson’s Thrush). See Table 3 for references.
Neotropical stopover regions for western-breeding migrants are almost completely unknown, but sites within south-western Mexico and the Pacific slope of Central America are likely to prove important for species on passage to South America, as they have for western populations of Swainson’s Thrush (Ruegg and Smith Reference Ruegg and Smith2002, Delmore et al. Reference Delmore, Fox and Irwin2012), and may mirror findings in north-west Mexico where moult-migration is integral to migration strategies (Rohwer et al. Reference Rohwer, Butler, Froehlich, Greenberg and Marra2005). Indeed, while this region is known to hold a diverse assemblage of over-wintering migrants (Howell and Webb Reference Howell and Webb1995, Hutto Reference Hutto1997, Fagan and Komar Reference Fagan and Komar2016), it may also host stopover populations of long-distance migrant species such as Yellow-billed Cuckoo Coccyzus americanus, Western Wood-Pewee Contopus sordidulus and Olive-sided Flycatcher Contopus cooperi.
Threats facing landbirds on migration in the Neotropics
Just as we learn about the importance of key regions to long-distance migratory birds within the Neotropics, new concerns are being raised about threats to specific sites and vital habitats within those regions, as well as the predicted effects of climate change. Habitat loss continues to be the leading threat to migratory bird populations throughout their annual cycle (Faaborg et al. Reference Faaborg, Holmes, Anders, Bildstein, Dugger, Gauthreaux, Heglund, Hobson, Jahn, Johnson, Latta, Levey and Marra2010a, Rosenberg et al. Reference Rosenberg, Kennedy, Dettmers, Ford, Reynolds and Alexander2016), and in areas where birds are highly concentrated during migration stopover, loss or degradation of critical habitats may contribute disproportionately to overall population declines (Baker et al. Reference Baker, González, Piersma, Niles, do Nascimento and Atkinson2004, Newton Reference Newton2006). In addition, direct mortality of migrating birds due to collisions with buildings, communication towers and other structures could be magnified in areas where large proportions of species’ populations concentrate. Although astounding numbers of birds are thought to perish through collisions in North America alone (Longcore et al. Reference Longcore, Rich, Mineau, MacDonald, Bert and Sullivan2013), including significant percentages of global populations of species of concern such as Golden-winged Warbler (Arnold and Zink Reference Arnold and Zink2011), equivalent data from south of the USA are few and far between (Agudelo-Álvarez et al. Reference Agudelo-Álvarez, Moreno-Velasquez and Ocampo-Peñuela2010). Nonetheless, urban and resort development along coastlines, and proliferation of wind-energy and communications infrastructure along coasts, narrow landmasses, and mountain passes could pose major challenges for migratory bird conservation (Cohen et al. Reference Cohen, Barrow, Buler, Deppe, Farnsworth, Marra, McWilliams, Mehlman, Wilson, Woodrey and Moore2017).
Research to date indicates that the majority of landbirds stopping over in Central and northern South America rely on native forests, especially pre-montane forests on Caribbean-facing slopes of coastal mountains, such as in the Sierra Nevada de Santa Marta in Colombia, as well as lowland tropical wet and dry forests along the coasts (Johnson and Winker Reference Johnson and Winker2008, Welton et al. Reference Welton, Anderson, Colorado, Hamel and Calderón-F2012, Gómez et al. Reference Gómez, Gómez-Bahamón, Cárdenas-Ortiz and Bayly2015). As elsewhere in the Neotropics, these tropical forests are under severe threat from expanding agriculture and development, and several of the stopover regions identified to date are experiencing high rates of deforestation and are considered hotspots for future deforestation (Wassenaar et al. Reference Wassenaar, Gerber, Verbury, Rosales, Ibrahim and Steinfeld2007). In addition, bird-friendly practices which have the potential to mitigate forest loss, such as shade-grown coffee, may not be as beneficial to birds on stopover (Bayly et al. Reference Bayly, Gómez, Hobson and Rosenberg2016) as they are considered to be for over-wintering migrants (Komar Reference Komar2006).
Much of south-eastern Mexico, such as in the state of Veracruz, has been deforested and although this trend has been reversed in recent years (Ruelas Inzunza et al. Reference Ruelas Inzunza, Goodrich, Hoffman, Martínez, Smith and Peresbarbosa2009), the extremely low percentage of remaining forest cover likely remains a barrier to a successful migration for many species. The expansion of cattle pastures in north-western and eastern Honduras and eastern Nicaragua also represents a major threat to forests that are expected to constitute important stopover habitats. In particular, the montane forests of north-western Honduras that are used by the threatened Cerulean Warbler during spring migration (Welton et al. Reference Welton, Anderson, Colorado, Hamel and Calderón-F2012) are under considerable pressure. Likewise, in Colombia, the Darién bottleneck is a hotspot for deforestation to make way for cattle pastures. The Sierra Nevada de Santa Marta is also under pressure, as both coffee cultivation and cattle pastures move up slope (Bayly et al. Reference Bayly, Paez, Gómez and Mora2012b). In western Mexico, loss of dry tropical forest (Trejo and Dirzo Reference Trejo and Dirzo2000) has likely diminished habitat, where many species have been shown to require primary forest (Hutto Reference Hutto1989).
While populations of most raptor species that migrate through the Neotropics are stable or increasing (Ruelas Inzunza et al. Reference Ruelas Inzunza, Goodrich and Hoffman2010a), dramatic changes along the main flyway could reverse current fortunes, particularly for species concentrating in large proportion in narrow corridors. The development of wind farms, in particular, has been cited as a potential hazard for migrating raptors but mortality rates reported elsewhere do not support this claim (De Lucas et al. Reference De Lucas, Janss, Whitfield and Ferrer2008, Marques et al. Reference Marques, Batalha, Rodrigues, Costa, Pereira and Fonseca2014). Given our excellent knowledge of the routes taken by raptors, however, any potentially detrimental projects can be identified rapidly and measures to mitigate their impact can be proposed based on sound scientific justification.
The next steps: research and conservation needs
The last five years have seen an upsurge in new information on the migratory strategies of a variety of landbirds, largely due to the innovation of geolocator technology (McKinnon et al. Reference McKinnon, Fraser and Stutchbury2013a), but also as a result of on-the-ground field studies and monitoring programmes in northern Central America (Martínez Leyva et al. Reference Martínez Leyva, Ruelas Inzunza, Cruz Carretero, Barr, Peresbarbosa Rojas and Chávez Domínguez2005), Colombia (Bayly et al. Reference Bayly, Gómez and Hobson2013) and Costa Rica (Elizondo-Camacho and Ralph Reference Elizondo-Camacho and Ralph2012). Despite our growing understanding of migratory strategies and stopover site use, we are still only scratching the surface of a topic that will require decades of dedicated research to determine the needs of most species. To increase our understanding of migration strategies, and to identify the most important stopover sites and habitats for declining Nearctic-Neotropical migratory birds, we recommend a coordinated research initiative combining targeted field studies with the latest technologies across key regions of the Neotropics: including regions 1, 2, 3, 6, 7, and 10 in Figure 3, premontane forests (600–1,500 m) in Panama and Costa Rica, Pacific dry forests in Costa Rica and Nicaragua, lowland dry forests in northern Colombia, and a general exploration of regions south of the equator. The goals of this initiative would be to (1) identify new and previously unknown stopover regions for migratory landbirds; (2) evaluate sites and habitats within those regions that are most critical for successful migration; and (3) develop strategies to conserve these sites and habitats.
The deployment of geolocators or GPS tags on a wider range of landbird species, especially those of conservation concern, is a clear research priority in order to identify new stopover regions, and studies are already underway on Cerulean, Golden-winged, Canada Cardellina canadensis, Kirtland’s Setophaga kirtlandii and Prothonotary Protonotaria citrea Warblers. It is also necessary to build on successful geolocator studies by ensuring representative sampling across breeding/wintering ranges, thereby shedding light on how stopover use varies between populations (Jahn et al. Reference Jahn, Cueto, Fox, Husak, Kim and Landoll2013, Fraser et al. Reference Fraser, Stutchbury, Kramer, Silverio, Barrow and Newstead2013, Hobson and Kardynal Reference Hobson and Kardynal2015) or the degree to which populations converge on major stopover regions, as occurs in Purple Martin (Fraser et al. Reference Fraser, Stutchbury, Kramer, Silverio, Barrow and Newstead2013), Veery (González-Prieto et al. Reference González-Prieto, Hobson, Bayly and Gómez2011, Heckscher et al. Reference Heckscher, Taylor, Fox and Afanasyev2011, Hobson and Kardynal Reference Hobson and Kardynal2015) and Bobolink (Renfrew et al. Reference Renfrew, Kim, Perlut, Smith, Fox and Marra2013).
While geolocators enable us to rapidly identify broad stopover regions, their lack of precision and inability to tell us about the habitats and resources that birds are using, means that more precise technology (e.g., GPS tags) and on-the-ground field studies in the Neotropics are essential. Such studies will not only help to pinpoint the actual location of birds within the broad regions identified by geolocators (McKinnon et al. Reference McKinnon, Stanley, Fraser, MacPherson, Casbourn and Marra2013b) but also determine what habitats are used and how their quality varies for birds on stopover (Bayly et al. Reference Bayly, Gómez, Hobson and Rosenberg2016). We therefore recommend that geolocator studies be followed up or complemented by standardised surveys within these broad regions, in combination with multi-year mark-recapture studies, that can: (1) estimate occupancy and persistence of landbirds by region and habitat during migration; (2) determine the energetic contribution of a given stopover region to migration strategies; (3) provide site-specific habitat and resource associations for defining realistic conservation priorities; and (4) provide estimates of use at the population level for communities of migrants.
The recapture of individually marked birds, for example, is essential to estimate stopover durations (Schaub et al. Reference Schaub, Pradel, Jenni and Lebreton2001), rates of energy accumulation (Schaub and Jenni Reference Schaub and Jenni2000), and ultimately the contribution that a given stopover site makes to the overall energetic needs of migration (Bayly et al. Reference Bayly, Gómez, Hobson, González and Rosenberg2012a). By comparing rates of mass gain (a surrogate for energy deposition) between habitats, we can also examine habitat quality (Dunn Reference Dunn2001, Bayly et al. Reference Bayly, Gómez, Hobson and Rosenberg2016). Mark-recapture studies can also make use of innovative techniques to gather data on difficult-to-catch and canopy-dwelling species, including the use of coded nano-tags and tower arrays to determine stopover durations and subsequent movements of tagged individuals (Gómez et al. Reference Gómez, Bayly, Norris, Mackenzie, Rosenberg, Taylor, Hobson and Cadena2017, Taylor et al. Reference Taylor, Crewe, Mackenzie, Lepage, Aubry, Crysler, Finney, Francis, Guglielmo, Hamilton, Holberton, Loring, Mitchell, Norris, Paquet, Ronconi, Smetzer, Smith, Welch and Woodworth2017).
Once we know where to focus conservation resources, the scale of action required to ensure adequate habitat availability in major stopover regions throughout the Neotropics is potentially overwhelming. To be successful, strategies to conserve stopover habitats need to be integrated within regional conservation efforts, including those focused on Neotropical resident and threatened species, protection of watersheds, reducing deforestation and socio-economic development (Barlow et al. Reference Barlow, Lennox, Ferreira, Berenguer, Lees, Mac Nally, Thomson, de Barros Ferraz, Louzada, Oliveira and Parry2016). Any approach must go above and beyond the strategic purchase of private reserves, given the considerable areas occupied by migratory birds, and include activities such as lobbying for changes in detrimental agricultural and land-use policies, community-based conservation efforts, ecotourism activities, promotion of migrant friendly agricultural practices such as shade-grown coffee and silvopastures, and the use of tools such as Payment for Environmental Services. Indeed, as we identify new stopover sites, the actions required to ensure that they continue to function as key links in the lives of migratory birds will have to be tailored to the unique conditions in each.
As bird conservation plans have focused largely on breeding habitat, and to a lesser extent on overwintering sites, new information on the specific needs of birds on migration must be incorporated into full life-cycle bird conservation plans (Berlanga et al. Reference Berlanga, Kennedy, Rich, Arizmendi, Beardmore and Blancher2010, Rosenberg et al. Reference Rosenberg, Kennedy, Dettmers, Ford, Reynolds and Alexander2016). Only through large-scale and highly collaborative efforts, such as the GoMAMN (https://www.gomamn.org/, Cohen et al. Reference Cohen, Barrow, Buler, Deppe, Farnsworth, Marra, McWilliams, Mehlman, Wilson, Woodrey and Moore2017) or the Neotropical Flyways Project (http://selva.org.co/research-programs/migratory-species/nfp/) will we be able to understand and conserve the spectacle of migration throughout the Western Hemisphere.
While our suggestions for continued and future research will yield important advances that can help guide conservation, some habitats and regions are so threatened that we can’t afford to wait for the next generation of data. Thus, we want to emphasise the importance of continuing ongoing habitat conservation and restoration efforts throughout the region, especially in natural habitats already in decline, as these efforts will undoubtedly benefit Neotropical migrant and resident species alike.
Acknowledgements
This review grew out of the Migration Stopover Special Session at the Partners in Flight V Conference and Conservation Workshop in Snowbird, Utah in August 2013, and we thank the organisers of Partners in Flight V, especially American Bird Conservancy, for including the session within the programme. Having begun as an overview of our knowledge prior to the special session, the discussions during the session highlighted the need for this review. We thank all participants in the session for their contributions and insights and subsequent discussions, especially, Rosa M. Vidal, Elisa Peresbarbosa, C. J. Ralph, Melinda Welton and Pablo Elizondo. This is contribution No. 1 of the Neotropical Flyways Project and Hawk Mountain conservation science contribution number 285. We thank an anonymous reviewer for suggestions that improved this manuscript. Attendance at PIFV by NB and KVR was supported by the Cornell Lab of Ornithology, and travel support for other conference participants was provided by American Bird Conservancy.







