


Introduction
Formulating a recovery plan for an endangered species requires a thorough understanding of source-sink dynamics because unidentified sink populations may delude management into imposing unachievable delisting requirements (Pulliam Reference Pulliam1988). This could ultimately lead to sink populations preventing a species from being delisted even if surrounding source populations have met recovery requirements and are viable. Therefore an understanding of the viability of sink populations within the context of source populations is needed to develop appropriate metapopulation conservation objectives (Harveson et al. Reference Harveson, Lopez, Silvy and Frank2004). An accurate assessment of viability requires long-term data capturing environmental variation across all life history stages (White Reference White, Boitani and Fuller2000). In the past, this data requirement had been lacking for many endangered species, but recent efforts to collect vital rate data and improve databases offer the opportunity to evaluate the recovery potential and viability of populations with greater confidence. This is especially the case for shorebirds (suborder Charadrii), which can be banded and monitored relatively easily to estimate survival, reproductive success, and dispersal using mark-recapture methods (Sandercock Reference Sandercock2003).
We conducted a population viability analysis (PVA) on a small, geographically isolated population of the Snowy Plover Charadrius nivosus that breeds in coastal northern California. The Snowy Plover (hereafter “plover”) is a small shorebird whose listed population segment along the Pacific Coast of the United States extends from southern Washington to the Mexico border in southern California (USFWS 2007). The plover mating system has been described as serial polygamy, because both sexes acquire new mates for the second or third successful breeding attempt over the six month breeding season, which begins in March and ends in September (Warriner et al. Reference Warriner, Warriner, Page and Stenzel1986). Female plovers lay a clutch of three eggs that are incubated for ∼28 days (Warriner et al. Reference Warriner, Warriner, Page and Stenzel1986). Upon hatching, females usually leave males to search for another mate, while males provide parental care of the brood; as a result, males father a maximum of six young in two broods each season (Warriner et al. Reference Warriner, Warriner, Page and Stenzel1986). In northern California, plovers are partial migrants, with a proportion of the subpopulation residing year-round and another proportion either wintering or breeding (Colwell et al. Reference Colwell, McAllister, Millet, Transou, Mullin, Nelson, Wilson and LeValley2007). Permanent emigration has been documented to occur as far as 790 km from natal sites but most yearlings disperse much shorter distances to their first breeding site (Colwell et al. Reference Colwell, McAllister, Millet, Transou, Mullin, Nelson, Wilson and LeValley2007, Stenzel et al. Reference Stenzel, Page, Warriner, Warriner, George, Eyster, Ramer and Neuman2007, Pearson and Colwell Reference Pearson and Colwell2013).
The United States Fish and Wildlife Service (USFWS) listed the Pacific Coast metapopulation of the Snowy Plover as threatened under the United States Endangered Species Act in 1993. The species’ Recovery Plan (USFWS 2007) identified three factors that are thought to limit recovery via negative effects on breeding productivity: 1) predation of eggs, chicks, and adults by native and introduced vertebrates, 2) encroachment of invasive vegetation such as European beach grass Ammophila arenaria, which degrades breeding habitats, and 3) human activity, which causes direct mortality of eggs, chicks, and adults or indirectly affects the distribution of plovers.
Nur et al. (Reference Nur, Page and Stenzel1999) produced a PVA that assessed the demographic response of the plover metapopulation to 19 scenarios that perturbed vital rates such as adult and juvenile survival, reproductive success, and dispersal. The results of this PVA provided guidance for the formulation of delisting criteria published in the species’ recovery plan (USFWS 2007). The recovery objective for the metapopulation was 3,000 breeding adults, with 150 needed in Recovery Unit 2 of coastal northern California (which includes Mendocino, Humboldt, and Del Norte counties; USFWS 2007).
The northern California population has been described as a sink by Mullin et al. (Reference Mullin, Colwell, McAllister and Dinsmore2010), who used an algebraic assessment of population growth using estimates of survival and productivity. We utilised 11 years of survival and productivity data collected from northern California and elsewhere along the Pacific Coast to assess the viability of the northern California population within the context of surrounding source populations. Our study had two main objectives pertaining to the viability of the northern California population: 1) to evaluate the relative importance of specific aspects of plover life history, and 2) to examine various management strategies that address the limiting factors of predation, habitat loss, and human disturbance. With these results, we evaluated the likelihood of population recovery given the current delisting requirements and made management recommendations specific to the northern California population that highlighted the importance of source-sink dynamics operating in the Pacific Coast metapopulation.
Study area
We monitored a colour-marked population of plovers from 2001 to 2011, on gravel bars of the Eel River and ocean-fronting sandy beaches in Humboldt County, California (Colwell et al. Reference Colwell, Burrell, Hardy, Kayano, Muir, Pearson, Peterson and Sesser2010). On beaches, plovers occupied habitat composed of fine homogeneous substrates littered with natural and human debris, and vegetated with large expanses of introduced European beach grass or small restored tracts of native flora. On gravel bars, plovers bred amidst heterogeneous substrates littered with driftwood and vegetated with sparse stands of willow Salix spp. and white sweet clover Melilotus alba. Colwell et al. (Reference Colwell, Burrell, Hardy, Kayano, Muir, Pearson, Peterson and Sesser2010) provided a detailed description of the study area and the distribution of plovers within these habitats.
Methods
Field methods
The population we studied has been intensively monitored each year, with nearly all adults and chicks colour-marked (Colwell et al. Reference Colwell, Burrell, Hardy, Kayano, Muir, Pearson, Peterson and Sesser2010, Mullin et al. Reference Mullin, Colwell, McAllister and Dinsmore2010). Between mid-March and mid-August, observers regularly surveyed known breeding sites and suitable breeding habitats; survey frequency increased when observers detected plovers at a site (Colwell et al. Reference Colwell, Burrell, Hardy, Kayano, Muir, Pearson, Peterson and Sesser2010). During surveys, observers resighted marked individuals, monitored nests, and determined parenthood based on multiple observations of banded plovers near the nest, adults incubating eggs, or tending chicks (Colwell et al. Reference Colwell, Burrell, Hardy, Kayano, Muir, Pearson, Peterson and Sesser2010). Observers conducted research under federal, state, and university permits.
Modelling population viability
We assessed population viability using VORTEX, version 9.99b (Lacy et al. Reference Lacy, Borbat and Pollak2003), a stochastic simulation software package. To investigate source-sink dynamics, we modelled northern California population viability by defining two subpopulations (Figure 1). The focal population was Recovery Unit 2 (hereafter “northern California”); neighbouring populations of Oregon, San Francisco Bay, and Monterey Bay constituted a single source population (hereafter “Pacific Coast”), which produce most immigrants to northern California (M. A. Colwell unpubl. data). VORTEX is an individual-based model that tracks the simulated fate of an individual through its lifetime (birth, survival, reproduction, movement, and death; Figure 2). VORTEX simulated population dynamics stochastically by drawing the annual success and fate of each individual from vital rate distributions we estimated and provided in the model (Table S1 in the online Supplementary Materials). VORTEX also required basic plover life history information such as clutch size, mating system, longevity, etc., which we obtained from published literature (Table S1). We used n = 5 as the quasi-extinction threshold. Therefore, we assumed that if the population declined to five individuals it would result in a major change in management practices (e.g. captive breeding). We projected simulations for 50 years and ran 1000 iterations per simulation to obtain accurate measures of stochastic population dynamics (Caswell Reference Caswell2001).

Figure 1. Map of Snowy Plover distribution along the Pacific Coast of the United States with the delineation of the two populations used in the stochastic model. NCA is the northern California focal population and PAC is a source population consisting of surrounding recovery units contributing immigrants to northern California.

Figure 2. Sequence of life-history events and probabilities (P) used by VORTEX to model population growth of Snowy Plovers in northern California (NCA) in the context of the Pacific Coast population (PAC), where ϕi is the survival of age class i and ψj, k is the movement between population j and population k.
Vital rate estimation
We used 11 years (2001 to 2011) of mark-resight and individual lifetime reproductive success data from northern California to estimate annual apparent survival, productivity, and movement parameters (Figure 2). We defined the resighting interval as the breeding season (i.e. interval over which pairs initiated clutches), a period extending from 15 March to 20 July (M. A. Colwell unpubl. data). See Table S2 in the supplementary materials for methods used to estimate plover vital rates.
Evaluating population viability
Long-term population growth is best predicted by the arithmetic mean of the population growth rates of many simulated iterations, known as the stochastic growth rate (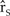 ${\hat r_S}$; Morris and Doak Reference Morris and Doak2002). To quantify the viability of the northern California plover population, we evaluated the
${\hat r_S}$; Morris and Doak Reference Morris and Doak2002). To quantify the viability of the northern California plover population, we evaluated the  ${\hat r_S}$ based on 1,000 iterations each simulated for 50 years. The units of
${\hat r_S}$ based on 1,000 iterations each simulated for 50 years. The units of 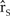 ${\hat r_S}$ are the number of individuals gained or lost per individual per year, with positive and negative values describing an increase and decrease in the population, respectively.
${\hat r_S}$ are the number of individuals gained or lost per individual per year, with positive and negative values describing an increase and decrease in the population, respectively.
There have been no studies assessing carrying capacity or density dependence in Snowy Plovers across the study area, and thus it was difficult to consider its role in population viability. Nur et al. (Reference Nur, Page and Stenzel1999), assumed a carrying capacity of 200 for northern California in their coast-wide PVA. Biologically this value may not be realistic, as plovers breed at low density (Page et al. Reference Page, Stenzel, Winkler and Swarth1983) and there appears to be plenty of unoccupied breeding and wintering habitat available in northern California (Brindock and Colwell Reference Brindock and Colwell2011, Burrell and Colwell Reference Burrell and Colwell2012). Nevertheless, we kept the carrying capacity consistent with Nur et al. (Reference Nur, Page and Stenzel1999) because the delisting requirement of 150 individuals for northern California was based on their findings.
The baseline model included vital rate distributions observed in both populations from 2001 to 2011. We evaluated the accuracy of the baseline model by comparing the coefficient of variation in population size observed between 2001 and 2011 with the 95% confidence interval of the mean coefficient of variation simulated across 30 ten year iterations, using the 2001 counts as the initial population sizes (N NCA = 57, N PAC = 437).
Elasticity analysis for stochastic population growth
A common approach to determine the extent to which vital rates affect population dynamics is to conduct an elasticity analysis, which quantifies the relative change in stochastic population growth resulting from a change in a vital rate (Morris and Doak Reference Morris and Doak2002). To estimate the elasticity of vital rates, we perturbed the vital rates of the baseline model by 10, 20, and 50% in the direction that would most likely increase 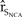 ${\hat r_{{S_{NCA}}}}$. We calculated elasticities as the proportional increase in
${\hat r_{{S_{NCA}}}}$. We calculated elasticities as the proportional increase in 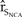 ${\hat r_{{S_{NCA}}}}$ divided by the proportional change in the vital rate (Morris and Doak Reference Morris and Doak2002), a method commonly used to calculate elasticities in PVAs (Benton and Grant Reference Benton and Grant1999, Finkelstein et al. Reference Finkelstein, Wolf, Goldman, Doak, Sievert, Balogh and Hasegawa2010, Hudgens et al. Reference Hudgens, Beaudry, George, Kaiser and Munkwitz2011).
${\hat r_{{S_{NCA}}}}$ divided by the proportional change in the vital rate (Morris and Doak Reference Morris and Doak2002), a method commonly used to calculate elasticities in PVAs (Benton and Grant Reference Benton and Grant1999, Finkelstein et al. Reference Finkelstein, Wolf, Goldman, Doak, Sievert, Balogh and Hasegawa2010, Hudgens et al. Reference Hudgens, Beaudry, George, Kaiser and Munkwitz2011).
Assessment of limiting factors
We developed hypothetical scenarios that evaluated the effectiveness of methods managing one or a combination of the three limiting factors affecting plover viability as identified by the recovery plan (USFWS 2007). Additionally, we assessed the influence of a prolonged cold winter weather catastrophe on population viability and recovery. We evaluated the results of these scenarios by comparing the 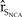 ${\hat r_{{S_{NCA}}}}$ to that of the baseline model.
${\hat r_{{S_{NCA}}}}$ to that of the baseline model.
Predation
Predation of eggs and chicks by Corvus brachyrhynchos and C. corax is the most influential factor affecting Snowy Plover productivity in northern California (Burrell and Colwell Reference Burrell and Colwell2012). Lethal predator removal and nest exclosures are two methods used to increase productivity that have been implemented at other populations across the Pacific Coast (USFWS 2007). Nest exclosures were used on 135 occasions in northern California between 2001 and 2006 and twice in 2010, and were designed to keep predators from eating eggs. There was no lethal removal of predators in northern California between 2001 and 2011. We simulated two scenarios that utilised lethal predator removal or nest exclosures in northern California to evaluate how these two methods may influence population viability. We implemented this by substituting the relevant northern California vital rates with those of the Pacific Coast where lethal predator removal has been practiced and nest exclosures were used to protect eggs and enhance hatching success since 2002 (unpubl. data from: D. J. Lauten et al., Oregon Biodiversity Information Center; G. W. Page et al., Point Reyes Bird Observatory; C. Robinson-Nilson et al., San Francisco Bay Bird Observatory). Likewise, to assess the use of only nest exclosures, we used northern California productivity rates estimated from 2001 to 2006 productivity data. In northern California, although nest exclosures are shown to increase chick fledgling rates (Watts et al. Reference Watts, Cao, Panza, Dugaw, Colwell and Burroughs2012), they are known to compromise the survival of incubating adults (Hardy and Colwell Reference Hardy and Colwell2008). To acknowledge this, we used survivorship vital rates estimated between 2001 and 2006 in northern California.
Habitat loss
The invasion of European beach grass has decreased the amount and quality of plover breeding habitat by increasing beach slope and creating less open space (Wiedemann Reference Wiedemann1987, Muir and Colwell Reference Muir and Colwell2010). Agencies occasionally restore plover habitat by removing European beach grass, which enhances recovery of the native dune ecosystem. With the creation of open habitat, we assumed that carrying capacity increased (USFWS 2007). To assess the influence of habitat restoration on population viability in northern California, we ran six management scenarios of the stochastic simulation that perturbed the baseline carrying capacity (K = 200) in both directions by 10, 20, and 50%.
Human activity
Human activity compromises plover productivity (Ruhlen et al. Reference Ruhlen, Abbott, Stenzel and Page2003, Lafferty et al. Reference Lafferty, Goodman and Sandoval2006, Wilson and Colwell Reference Wilson and Colwell2010) and survival (Brindock and Colwell Reference Brindock and Colwell2011), and affects the distribution of plovers on a beach (Lafferty et al. Reference Lafferty, Goodman and Sandoval2006). Fencing, consisting of a roped boundary with signs posted to inform the public that the delineated area is restricted, is commonly used to reduce human disturbance (USFWS 2007). In northern California, plover fledging success was almost twice as high within a fenced area than outside the fence (Wilson and Colwell Reference Wilson and Colwell2010). Additionally, Lafferty et al. (2006) observed a more than 50% reduction in disturbance events after the installation of a fence. Survival may be compromised by human activities, such as driving vehicles through wintering flocks and occasionally killing plovers (Brindock and Colwell Reference Brindock and Colwell2011).
We assessed the response of the population to a reduction in human disturbance by simulating scenarios in which we matched productivity distributions to those observed inside fencing (Wilson and Colwell Reference Wilson and Colwell2010) and increased the survival of adults and juveniles by 2%, 5% or 10%. There are no data to quantitatively assess how varying levels of vehicle use on beaches reduces plover survival probability, therefore we used a range of perturbations that we believe is conservative given anecdotal reports of shorebird mortality associated with vehicle strikes elsewhere within the winter range on the plover (Buchanan Reference Buchanan2011).
Severe winter weather
Severe weather, such as low temperature anomalies, can reduce the over-winter survival of shorebirds (Parr Reference Parr1992, Yalden and Pearce-Higgins Reference Yalden and Pearce-Higgins1997, Durell et al. 2006, Roche et al. Reference Roche, Cohen, Catlin, Amirault-Langlais, Cuthbert, Gratto-Trevor, Felio and Fraser2010) and plovers in northern California (Eberhart-Phillips Reference Eberhart-Phillips2012). Estimates from the vicinity of Monterey Bay indicate especially low overwinter juvenile and adult survival in 1987 and 1998 (Stenzel et al. Reference Stenzel, Page, Warriner, Warriner, George, Eyster, Ramer and Neuman2007, Reference Stenzel, Page, Warriner, Warriner, Neuman, George, Eyster and Bidstrup2011), which may be related to climatic events like the winter of 1998 during which there were seven consecutive days with a minimum temperature at or below freezing.
Severe winter weather catastrophes were estimated to reduce survival by 26% in northern California and by 7% in the Pacific Coast, with a 15% probability of occurring in any one year (Eberhart-Phillips Reference Eberhart-Phillips2012). We used these catastrophe statistics as parameters in VORTEX to simulate the influence of a catastrophe on the viability of the northern California plover population.
Results
Vital rate estimation
At the end of the 2011 breeding season, our dataset included 224 males and 244 females; 176 were immigrants originating from the Pacific Coast, 64 were recruited locally within northern California, and 228 were individuals that hatched in northern California but did not return in subsequent breeding seasons. Table S3 gives results pertaining to plover vital rate estimates.
Modelling
The observed coefficient of variation in population size between 2001 and 2011 (NCA = 0.366, PAC = 0.207) fell within the 95% confidence intervals of the coefficient of variation of the 2001 to 2011 baseline model 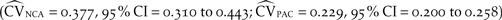 $\left( {{{\widehat {CV}}_{NCA}} = 0.377,\;95\% \,CI = 0.310\;to\;0.443;\;{{\widehat {CV}}_{PAC}} = 0.229,\;95\% \,CI = 0.200\;to\;0.258} \right)$, demonstrating that the structure of the baseline model accurately predicted the observed stochasticity. The observed trend of northern California deviated positively from the simulated projection between 2001 and 2006, but between 2007 and 2011 the model effectively predicted the observed trend (Figure 3a). Between 2001 and 2011, the simulated projection of the Pacific Coast population slightly underestimated the observed population size, but the observed trend fell within the 95% confidence interval of the simulation’s average projection for all years (Figure 3b).
$\left( {{{\widehat {CV}}_{NCA}} = 0.377,\;95\% \,CI = 0.310\;to\;0.443;\;{{\widehat {CV}}_{PAC}} = 0.229,\;95\% \,CI = 0.200\;to\;0.258} \right)$, demonstrating that the structure of the baseline model accurately predicted the observed stochasticity. The observed trend of northern California deviated positively from the simulated projection between 2001 and 2006, but between 2007 and 2011 the model effectively predicted the observed trend (Figure 3a). Between 2001 and 2011, the simulated projection of the Pacific Coast population slightly underestimated the observed population size, but the observed trend fell within the 95% confidence interval of the simulation’s average projection for all years (Figure 3b).

Figure 3. Comparison of observed and predicted population projections for (a) northern California and (b) the Pacific Coast between 2001 and 2011 using the 2001 initial population sizes and the baseline vital rates. Baseline model population projections for (c) northern California and (d) the Pacific Coast. Error bars represent the 95% confidence interval of the average population size of 1,000 iterations.
When we projected the model into the future using the 2011 counts as the initial population sizes (N NCA = 36, N PAC = 799), the baseline model experienced slightly positive growth 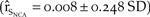 $\left( {{{\hat r}_{{S_{NCA}}}} = 0.008 \pm 0.248\;SD} \right)$ such that by the fiftieth year, northern California had an average of 59 individuals with a 0.2% probability of quasi-extinction (Figure 3c). Similarly, the baseline model of the Pacific Coast exhibited positive growth (
$\left( {{{\hat r}_{{S_{NCA}}}} = 0.008 \pm 0.248\;SD} \right)$ such that by the fiftieth year, northern California had an average of 59 individuals with a 0.2% probability of quasi-extinction (Figure 3c). Similarly, the baseline model of the Pacific Coast exhibited positive growth (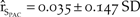 ${\hat r_{{S_{PAC}}}} = 0.035 \pm 0.147\;SD$; Figure 3d) and increased to an average of 1054 individuals after 50 years with a 0% probability of quasi-extinction.
${\hat r_{{S_{PAC}}}} = 0.035 \pm 0.147\;SD$; Figure 3d) and increased to an average of 1054 individuals after 50 years with a 0% probability of quasi-extinction.
The elasticity of 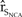 ${\hat r_{{S_{NCA}}}}$ was greater for perturbations of Pacific Coast vital rates than for perturbations in the vital rates of northern California, with the productivity of the Pacific Coast having the largest influence on
${\hat r_{{S_{NCA}}}}$ was greater for perturbations of Pacific Coast vital rates than for perturbations in the vital rates of northern California, with the productivity of the Pacific Coast having the largest influence on 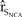 ${\hat r_{{S_{NCA}}}}$ (Figure 4). For example, a 10% increase in the expected brood and fledging parameters of the Pacific Coast increased the
${\hat r_{{S_{NCA}}}}$ (Figure 4). For example, a 10% increase in the expected brood and fledging parameters of the Pacific Coast increased the 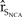 ${\hat r_{{S_{NCA}}}}$ 1.81 times more than a 50% increase in northern California adult survival, 10 times more than a 50% increase in the expected brood and fledging parameters of northern California, and 20 times more than a 50% increase in the carrying capacity of northern California.
${\hat r_{{S_{NCA}}}}$ 1.81 times more than a 50% increase in northern California adult survival, 10 times more than a 50% increase in the expected brood and fledging parameters of northern California, and 20 times more than a 50% increase in the carrying capacity of northern California.

Figure 4. Elasticity of the stochastic growth rate of the northern California population for 21 model components specific to northern California (NCA), the Pacific Coast (PAC), or globally affecting both populations simultaneously (immigration, emigration, the winter cold weather catastrophe, and inbreeding). Total productivity represents a perturbation of both the broods per male and fledglings per brood distributions. All perturbations were made in the direction that would hypothetically favour growth in northern California.
Immigration was an important component of the model. A 20% increase in the immigration rate had a greater effect on northern California growth than all other northern California productivity vital rates (Figure 4). A scenario in which no movement occurred between the two populations resulted in 100% probability of quasi-extinction of northern California after an average of five years and a 46% increase in growth for the Pacific Coast population 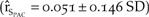 $\left( {{{\hat r}_{{S_{PAC}}}} = 0.051 \pm 0.146\;SD} \right)$.
$\left( {{{\hat r}_{{S_{PAC}}}} = 0.051 \pm 0.146\;SD} \right)$.
Our estimate of northern California juvenile apparent survival 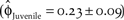 $\left( {{{\hat \phi }_{Juvenile}} = 0.23 \pm 0.09} \right)$ was 50% of the true juvenile survival estimated in Monterey Bay
$\left( {{{\hat \phi }_{Juvenile}} = 0.23 \pm 0.09} \right)$ was 50% of the true juvenile survival estimated in Monterey Bay 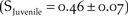 $\left( {{S_{Juvenile}} = 0.46 \pm 0.07} \right)$ by Stenzel et al. (Reference Stenzel, Page, Warriner, Warriner, George, Eyster, Ramer and Neuman2007). This suggests our estimate of juvenile apparent survival in northern California was biased low because it is confounded with permanent emigration (Sandercock Reference Sandercock2003). To evaluate the relative difference in the two estimates of juvenile survival, we simulated a model in which juvenile survival and variation of northern California was equal to that of the Pacific Coast. This resulted in a 50% increase in the growth of northern California over the baseline
$\left( {{S_{Juvenile}} = 0.46 \pm 0.07} \right)$ by Stenzel et al. (Reference Stenzel, Page, Warriner, Warriner, George, Eyster, Ramer and Neuman2007). This suggests our estimate of juvenile apparent survival in northern California was biased low because it is confounded with permanent emigration (Sandercock Reference Sandercock2003). To evaluate the relative difference in the two estimates of juvenile survival, we simulated a model in which juvenile survival and variation of northern California was equal to that of the Pacific Coast. This resulted in a 50% increase in the growth of northern California over the baseline  $\left( {{{\hat r}_{{S_{NCA}}}} = 0.012 \pm 0.23\;SD,\;{N_{50}} = 72} \right)$; however, the delisting criterion of 150 breeding adults was not achieved within the 50-year simulation.
$\left( {{{\hat r}_{{S_{NCA}}}} = 0.012 \pm 0.23\;SD,\;{N_{50}} = 72} \right)$; however, the delisting criterion of 150 breeding adults was not achieved within the 50-year simulation.
Assessment of limiting factors
Nest and brood predation
To simulate the use of lethal predator removal in northern California, we increased the expected successful broods per breeding male from 0.44 to 0.84 and the expected fledglings per successful brood from 1.82 to 1.87, which were the productivity rates observed across the Pacific Coast where such management has been practiced. This simulation resulted in a 25% increase in the growth of northern California over the baseline. However the population did not reach the recovery objective of 150 individuals (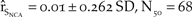 ${\hat r_{{S_{NCA}}}} = 0.01 \pm 0.262\;SD,\;{N_{50}} = 68$; Figure 5a).
${\hat r_{{S_{NCA}}}} = 0.01 \pm 0.262\;SD,\;{N_{50}} = 68$; Figure 5a).

Figure 5. Population projections for northern California under four management approaches addressing either predation or human disturbance: (a) lethal predator removal, (b) the use of nest exclosures, (c) fencing, and (d) fencing with three levels of hypothetical reductions in mortality related to vehicle strike. Error bars represent the 95% confidence interval of the average population size of 1,000 iterations.
Likewise, to simulate the use of nest exclosures we increased the expected successful broods per breeding male from 0.44 to 0.54 and decreased adult survival from 0.62 to 0.57, which were the brood and survival rates observed and estimated in northern California between 2001 and 2006 when nest exclosures were used. This simulation resulted in a 12.5% decrease in the growth of northern California  $\left( {{{\hat r}_{{S_{NCA}}}} = 0.007 \pm 0.255\;SD} \right)$ over the baseline (Figure 5b).
$\left( {{{\hat r}_{{S_{NCA}}}} = 0.007 \pm 0.255\;SD} \right)$ over the baseline (Figure 5b).
Habitat loss
All perturbations of carrying capacity in northern California that simulated habitat restoration did not result in a difference in 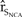 ${\hat r_{{S_{NCA}}}}$ from the baseline. However, increasing the carrying capacity of the Pacific Coast by 10% from 1,300 to 1,450 individuals resulted in a 12.5% increase in the growth in northern California.
${\hat r_{{S_{NCA}}}}$ from the baseline. However, increasing the carrying capacity of the Pacific Coast by 10% from 1,300 to 1,450 individuals resulted in a 12.5% increase in the growth in northern California.
Human disturbance
We assessed management of human disturbance using two models. The first model simulated a doubling in productivity through the use of fencing throughout all suitable breeding habitat in northern California. This model differed from the current management regime, which has used fencing sparingly at a few sites. Doubling productivity resulted in a 50% increase in the growth of northern California 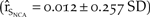 $\left( {{{\hat r}_{{S_{NCA}}}} = 0.012 \pm 0.257\;SD} \right)$ from that of the baseline (Figure 5c).
$\left( {{{\hat r}_{{S_{NCA}}}} = 0.012 \pm 0.257\;SD} \right)$ from that of the baseline (Figure 5c).
The second model simulated the use of fencing and prohibition of recreational vehicle use near known plover habitat. We simulated this management scenario by using the productivity rates of the first model and by increasing adult and juvenile survival by 2%, 5% or 10%. With fencing and a 2% increase in survival, there was a 50% increase in the population growth 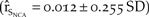 $\left( {{{\hat r}_{{S_{NCA}}}} = 0.012 \pm 0.255\;SD} \right)$ over the baseline (Figure 5d). However with fencing and a 10% increase in survival, there was a 140% increase in the baseline growth of northern California (
$\left( {{{\hat r}_{{S_{NCA}}}} = 0.012 \pm 0.255\;SD} \right)$ over the baseline (Figure 5d). However with fencing and a 10% increase in survival, there was a 140% increase in the baseline growth of northern California (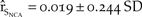 ${\hat r_{{S_{NCA}}}} = 0.019 \pm 0.244\;SD$; Figure 5d).
${\hat r_{{S_{NCA}}}} = 0.019 \pm 0.244\;SD$; Figure 5d).
Severe winter weather
The severity of the cold winter catastrophe was an important component of the model. A 10% reduction in severity had a greater elasticity than all perturbations of northern California vital rates, except a 50% reduction in adult mortality (Figure 4). Reducing the catastrophe severity by 10% resulted in a 25% increase in the growth of northern California 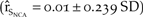 $\left( {{{\hat r}_{{S_{NCA}}}} = 0.01 \pm 0.239\;SD} \right)$. The elasticities of northern California growth to perturbations in the frequency of catastrophes were not as important as those of the severity (Figure 4). A 50% reduction in the frequency resulted in a 12.5% increase in growth over the baseline
$\left( {{{\hat r}_{{S_{NCA}}}} = 0.01 \pm 0.239\;SD} \right)$. The elasticities of northern California growth to perturbations in the frequency of catastrophes were not as important as those of the severity (Figure 4). A 50% reduction in the frequency resulted in a 12.5% increase in growth over the baseline 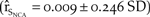 $\left( {{{\hat r}_{{S_{NCA}}}} = 0.009 \pm 0.246\;SD} \right)$. Removing the catastrophe from the model resulted in a 37.5% increase in population growth in northern California
$\left( {{{\hat r}_{{S_{NCA}}}} = 0.009 \pm 0.246\;SD} \right)$. Removing the catastrophe from the model resulted in a 37.5% increase in population growth in northern California 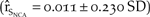 $\left( {{{\hat r}_{{S_{NCA}}}} = 0.011 \pm 0.230\;SD} \right)$.
$\left( {{{\hat r}_{{S_{NCA}}}} = 0.011 \pm 0.230\;SD} \right)$.
Discussion
Four main findings emerged regarding the viability of the plover subpopulation in northern California: 1) the population is a sink that relies upon immigrants originating from adjacent populations; 2) population persistence depends on continued management aimed at increasing plover productivity at source populations elsewhere on the Pacific Coast; 3) local management should consider the risks associated with the use of nest exclosures and acknowledge the benefits of reducing human disturbance and implementing predator removal; and 4) decreased survival associated with cold winter weather strongly influenced population growth.
Model performance
Baseline projections forecasting 50 years indicated that both the northern California and Pacific Coast populations will increase. The structure of the baseline model reflected the observed stochasticity accurately; however, the simulated trajectory of northern California underestimated the observed population size between 2001 and 2006. The shortfall of the model between 2001 and 2006 most likely resulted from two things. First, the 2001–2006 interval did not experience winters with prolonged cold weather (Eberhart-Phillips Reference Eberhart-Phillips2012); whereas on average, simulations included one or more catastrophic winters during this interval which depressed simulated growth rates. Second, productivity parameters of the baseline model were based upon productivity data averaged over the entire 11-year period; whereas between 2001 and 2006, nest exclosures were used to manage nest predators which resulted in higher nest success during that interval.
Population viability
The productivity and survival rates used in the model do not support a self-sustaining plover population in northern California. Consequently, immigration from source populations elsewhere along the Pacific Coast is necessary to maintain a breeding population in northern California. Our study confirmed the role of immigration in northern California using state-of-the-art methods and updated demographic data. Mullin et al. (Reference Mullin, Colwell, McAllister and Dinsmore2010) classified the northern California population as a sink based on an algebraic assessment of population growth using estimates of productivity and survival. And in the PVA by Nur et al. (Reference Nur, Page and Stenzel1999), when dispersal was reduced to zero, the northern California population became extinct. Plissner and Haig (Reference Plissner and Haig2000) derived similar conclusions from their PVA of the Atlantic Coast breeding metapopulation of Piping Plovers C. melodus: persistence time increased with increased movement rates and greater connectivity increased the total size of the metapopulation. For Snowy Plovers, this emphasises the importance of dispersal to counterbalance regional low productivity or survival, such as that experienced in northern California (USFWS 2007).
The influence of inbreeding on population growth was relatively low compared to other components of the model. This suggests that during simulations, immigration of individuals from the Pacific Coast replenished genetic variation in northern California, subsequently reducing the impact of inbreeding depression. This phenomenon, known as ‘genetic rescue’ (Ingvarsson Reference Ingvarsson2001), highlights another reason why immigration is crucial for the viability of the northern California population.
Our estimates of variation in adult and juvenile survival in northern California were greater than those of the Pacific Coast, which meant that simulations of northern California experienced large annual fluctuations in population growth given the high elasticity of adult and juvenile mortality. In addition to its low productivity, the large stochasticity of influential vital rates (e.g. survival) characterises the northern California population as a sink. This was further supported by the “no movement” scenario, which demonstrated that the growth of the Pacific Coast population increased by 46% when immigration to northern California ceased. Viewed another way, the northern California population is a burden to the recovery of the rest of the Pacific Coast metapopulation.
Delisting of the plover requires a metapopulation of 3,000 breeding adults maintained for 10 years distributed disproportionately among the recovery units and an average annual productivity of one fledged chick per breeding male for five years (USFWS 2007). Recovery of the northern California population requires 150 breeding adults and Recovery Units 1, 3, and 4 (which make up the Pacific Coast population in this study) collectively require 1,150 breeding adults (USFWS 2007). The Pacific Coast population will likely achieve the delisting requirements within the next 30 years given that the population currently produces an annual average of 1.32 fledged chicks per breeding male and the baseline simulation had an average of 1,060 ± 269 SD individuals after 20 years. However, the northern California population is unlikely to reach the recovery objectives given that the population currently produces an annual average of 0.68 fledged chicks per male and the baseline simulation had an average of 60 ± 23 SD individuals after 20 years. Even those simulations with 50% reductions in the mortality of adults or juveniles in northern California resulted in insufficient population growth needed to achieve 150 breeding adults. This finding highlights that the delisting of the entire Pacific Coast Snowy Plover metapopulation will likely be precluded by sink populations such as northern California, given the current delisting requirements.
Although northern California is a sink, its persistence may be important for the viability of the entire metapopulation. Howe et al. (Reference Howe, Davis and Mosca1991) argued that a metapopulation with a large proportion of individuals residing in sinks can still be viable. Furthermore, Howe et al. (Reference Howe, Davis and Mosca1991) found through simulations that in many cases sink populations can 1) increase the overall size of a metapopulation, 2) increase the size of source populations, and 3) extend the persistence of metapopulations in decline. Howe et al. (Reference Howe, Davis and Mosca1991) argued that sink populations offer available habitat for emigrants from crowded source populations to persist, albeit under conditions that are unsustainable in isolation. Therefore, the protection and maintenance of productive populations such as those in Oregon and Monterey Bay must be a priority for Snowy Plover management, but the preservation of marginal populations such as northern California should still be part of the conservation plan. The 4(d) rule proposed by the U.S. Department of the Interior (2006) would have allowed regions to relax management if they had met the recovery criteria. However, without a productive source population, the northern California population would likely be pushed to extinction within the next 20 years.
Management of limiting factors
Management scenarios addressing human disturbance were the most beneficial for the growth of the northern California population. Population growth increased by 50% through the use of fencing and a further 25% with the prohibition of recreational vehicle use on beaches (assuming a 5% reduction in mortality). It is important to note that our fencing scenario assumed that fencing would be implemented across all beaches in northern California, which is unrealistic. However, our results demonstrate the benefits of managing human disturbance. Wilson and Colwell (Reference Wilson and Colwell2010) found that fencing in northern California increased productivity through a reduction in brood movement. Because of human disturbance, chicks that were raised outside of the fenced area travelled greater distances than those raised in it, potentially exposing broods to higher risks of mortality (Wilson and Colwell Reference Wilson and Colwell2010). The influence of human disturbance on plover productivity in northern California is almost certainly augmented by corvid predation of broods, which coincides with areas of human activity (Marzluff and Neatherlin Reference Marzluff and Neatherlin2006).
Recreational vehicle use in plover habitat is a complicated issue, considering that there is only anecdotal evidence to support its negative influence on survival in northern California (Brindock and Colwell Reference Brindock and Colwell2011). However, the potential effects of vehicle use on population growth in northern California must be acknowledged by management because survival, especially that of adults, had the greatest effect on plover population growth; this result is common to other shorebirds (Sandercock Reference Sandercock2003). This may be especially important during the non-breeding season, when plover survival is likely compromised by severe winter weather (Yalden and Pearce-Higgins Reference Yalden and Pearce-Higgins1997, Eberhart-Phillips Reference Eberhart-Phillips2012). It may be politically unfeasible to enforce a coast-wide prohibition of recreational vehicle use on the beaches in northern California, but regional prohibition at locations where plovers predictably over-winter (see Brindock and Colwell 2010) could be an alternative. After a regional ban of vehicle use on beaches in South Africa, Williams et al. (Reference Williams, Ward and Underhill2004) observed a doubling in populations of Crowned Plover Vanellus coronatus, Blacksmith Plover V. armatus, and Sanderling Calidris alba, suggesting that such management was effective.
Predation of eggs and chicks, mainly by corvids, is the most important factor limiting productivity in northern California (Burrell and Colwell Reference Burrell and Colwell2012). Simulations demonstrated that managing predation with lethal methods was 43% more effective at improving population growth in northern California than the use of nest exclosures. The elasticity analysis revealed that the expected number of successful broods per nesting male was the most influential productivity vital rate of northern California affecting growth. This provides support for the use of lethal predator removal in northern California because lethal methods improve both egg and chick survival, whereas exclosures only protect nests and are known to compromise the survival of incubating adults (Hardy and Colwell Reference Hardy and Colwell2008), the most elastic vital rate. This finding is supported by Watts et al. (Reference Watts, Cao, Panza, Dugaw, Colwell and Burroughs2012) who concluded that a 10% increase in adult mortality resulting from nest exclosures would result in a declining population in northern California.
Habitat restoration scenarios revealed that for the current population size of northern California (N 2011 = 36), the assumed carrying capacity (K = 200) is not restricting population growth. All simulations that perturbed carrying capacity (positively or negatively) did not strongly affect population growth. This finding relies upon the assumption of the model that habitat restoration only increases carrying capacity and not vital rates such as survival and productivity. There is no evidence to suggest an increase in survival associated with habitat restoration; however, restoration may indirectly enhance nest survival through the creation of open habitat which facilitates an early detection of predators by incubating plovers (Koivula and Rönkä Reference Koivula and Rönkä1998). Muir and Colwell (Reference Muir and Colwell2010) found that breeding plovers selected nest sites that were more open than random sites; however there was no evidence to suggest that restored habitat was of higher quality because measures of enhanced productivity associated with open habitat were not evaluated. Although our findings suggest that habitat restoration is not a priority for the recovery of plovers in northern California, it is important to stress that other species benefit from restoration of the native dune ecosystem.
The simulated cold winter weather catastrophe strongly influenced population growth in northern California and is likely a factor contributing to its status as a sink. Evidence suggests that cold winter anomalies caused decreased survival (Eberhart-Phillips Reference Eberhart-Phillips2012), a vital rate with the highest elasticity in northern California. Winter weather severity was highly variable between 2001 and 2011 and is thus a probable source for the large variation in both adult and juvenile survival observed over the 11-year period (Eberhart-Phillips Reference Eberhart-Phillips2012). Stochastic weather events make small isolated populations like northern California especially vulnerable to extinction (Crick Reference Crick2004). The influence of climate highlighted by this study provides evidence that factors beyond direct control of humans affect population viability.
Pulliam (Reference Pulliam1988) argued that sink populations could easily mislead management into formulating unrealistic population size expectations if the underlying source-sink dynamics of the metapopulation were not fully understood. The delisting criteria for plovers in northern California was formulated based upon a metapopulation viability analysis (Nur et al. Reference Nur, Page and Stenzel1999), that made large assumptions pertaining to population-specific vital rates because of data limitations at the time of analysis. For example, in their model, Nur et al. (Reference Nur, Page and Stenzel1999) included 1) higher estimates of adult 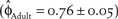 $\left( {{{\hat \phi }_{Adult}} = 0.76 \pm 0.05} \right)$ and juvenile
$\left( {{{\hat \phi }_{Adult}} = 0.76 \pm 0.05} \right)$ and juvenile 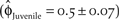 $\left( {{{\hat \phi }_{Juvenile}} = 0.5 \pm 0.07} \right)$ survival with less variation, and 2) a catastrophe that reduced productivity by 50% every 20 years. Nur et al. (Reference Nur, Page and Stenzel1999) also concluded that northern California is a sink; however, the differences between the parameters used in their model and those of this study are a likely source for the differing conclusions regarding an achievable recovery population size in northern California.
$\left( {{{\hat \phi }_{Juvenile}} = 0.5 \pm 0.07} \right)$ survival with less variation, and 2) a catastrophe that reduced productivity by 50% every 20 years. Nur et al. (Reference Nur, Page and Stenzel1999) also concluded that northern California is a sink; however, the differences between the parameters used in their model and those of this study are a likely source for the differing conclusions regarding an achievable recovery population size in northern California.
Results of this study draw attention to active source-sink processes working within the Pacific Coast Snowy Plover metapopulation. Managers must collaborate to focus conservation efforts on maintaining viable source populations but not neglect sinks. Our study suggests that some of the delisting criteria in the Snowy Plover Recovery Plan may be unrealistic, which could prevent the Pacific Coast metapopulation from being delisted. However, because this study focused specifically upon northern California, our model was limited to a simple source-sink structure and thus we must be cautious about drawing inferences about the status of other subpopulations and their relationship to surrounding sinks or sources. Therefore, given the coast-wide efforts by plover biologists to collect ample vital rate data since the analysis by Nur et al. (Reference Nur, Page and Stenzel1999), we recommend that future work should be focused on a thorough reassessment of metapopulation structure and viability of Snowy Plovers across the Pacific Coast. This will provide valuable information pertaining to the recovery potential of Snowy Plovers given the current delisting requirements.
Supplementary Material
The supplementary materials for this article can be found at journals.cambridge.org/bci
Acknowledgements
Many individuals assisted us with fieldwork in northern California; in particular, we thank K. Brindock, N. Burrell, W. Goldenberg, J. Hall, M. Hardy, J. Harris, D. Herman, A. Hoffmann, S. Hurley, R. LeValley, A. Liebenberg, S. McAllister, J. Meyer, C. Millett, S. Mullin, J. Muir, Z. Nelson, A. Patrick, W. Pearson, S. Peterson, K. Ross, K. Sesser, R. Thiem, A. Transou, J. Watkins, and C. Wilson. This study would not be possible without the fieldwork and reports from many biologists across the Pacific Coast; in particular we thank K. Castelein, D. Lauten, G. Page, C. Robinson-Nilson, and L. Stenzel. S. Dinsmore, T. L. George, B. Hudgens, and an anonymous reviewer assisted with revisions of the modelling procedure and provided constructive comments to this manuscript. Field work was funded by the California Department of Fish and Game, California Department of Parks and Recreation, Chevron Oil Corporation, Humboldt State University Sponsored Programs Foundation, Marin Rod and Gun Club, MRB Research, Inc., U.S. Bureau of Land Management, U.S. Fish and Wildlife Service, and California Department of Fish and Game’s Oil Spill Response Trust Fund through the Oiled Wildlife Care Network at the Wildlife Health Center, School of Veterinary Medicine, University of California, Davis.







