


Introduction
In ungulates, two main types of mother–young relationships have been described, depending on the behaviour of the neonate. On the one hand, some species give birth to neonates that start to follow their mother as soon as she moves from the birth site, and which are therefore classified as ‘followers’ (Lent, Reference Lent1974). In other species, neonates do not follow their mother when she goes foraging. Rather, for the first few days they lie down and hide, waiting for the mother to return for nursing, hence their classification as ‘hiders’. Sheep belong to the first category, and the system of mother–young recognition has been extensively studied in this species (Lévy et al., Reference Lévy, Kendrick, Keverne, Porter and Romeyer1996; Kendrick et al., Reference Kendrick, Da Costa, Broad, Ohkura, Guevara, Lévy and Keverne1997; Nowak et al., Reference Nowak, Breton and Mellot2001). In contrast, work on goats, especially regarding the neonate, is rather scarce and little information is available concerning the establishment of maternal recognition by the newborn kid. In the sheep, the neonate develops a preference for its dam within 12 to 24 h (Nowak et al., Reference Nowak, Poindron, Le Neindre and Putu1987; Val-Laillet et al., Reference Val-Laillet, Simon and Nowak2004), initially on the basis of the acceptance behaviours displayed by the mother (Terrazas et al., Reference Terrazas, Nowak, Serafin, Ferreira, Lévy and Poindron2002), followed later by true recognition of her individual physical features. Thus, lambs become able to choose between their mother and an alien dam from a distance of several metres from day 3 of age onwards (Nowak, Reference Nowak1990) and can recognise her bleats (Sèbe et al., Reference Sèbe, Aubin, Nowak and Poindron2007).
Goats, in contrast to sheep, belong to the hider category, whether under feral or domestic conditions (Lent, Reference Lent1974; Lickliter, Reference Lickliter1985). Studies concerning the discrimination of the kid by its dam have shown that the mechanisms involved are similar to those found in sheep (Romeyer et al., Reference Romeyer, Porter, Lévy, Nowak, Orgeur and Poindron1993, Reference Romeyer, Poindron and Orgeur1994a and Reference Romeyer, Poindron, Porter, Lévy and Orgeur1994b; Poindron et al., Reference Poindron, Gilling, Hernandez, Serafin and Terrazas2003). In contrast, it is not known whether the hiding behaviour of kids is associated with marked differences in their discrimination abilities compared with that of lambs. In particular, it is not known whether the hiding behaviour of kids is associated with a later display of discrimination of the dam than in lambs. There is some evidence that kids can show a preference for their mother at 2 days of age (Lickliter and Heron, Reference Lickliter and Heron1984; Addae et al., Reference Addae, Awotwi, Oppong-Anane and Oddoye2000) but not earlier, whereas lambs already discriminate their dam at 12 h (Nowak et al., Reference Nowak, Poindron, Le Neindre and Putu1987; Terrazas et al., Reference Terrazas, Nowak, Serafin, Ferreira, Lévy and Poindron2002; Val-Laillet et al., Reference Val-Laillet, Simon and Nowak2004). While this difference may be related to the fact that kids hide and lambs do not, it may also depend on other factors. Thus in sheep, breed is well known to influence the early ability of the mother and of the neonate to recognise each other (Shillito-Walser et al., Reference Shillito-Walser, Willadsen and Hague1982; Nowak et al., Reference Nowak, Poindron, Le Neindre and Putu1987). It cannot be excluded that the same applies in goats, especially since Addae et al. (Reference Addae, Awotwi, Oppong-Anane and Oddoye2000) suggested that mothers of the breed they studied had poor learning abilities. Indeed, they found that recognition was not clearly established before 48 h post partum, whereas in a recent study using a similar testing situation we found that crossbred dairy-type goats already recognised their kids at 4 h post partum (Poindron et al., Reference Poindron, Gilling, Hernandez, Serafin and Terrazas2003). Therefore, we investigated whether 8-, 12- and 24-h-old kids displayed a preference for their mother in a two-choice test and, if so, whether this initial preference depended on differences in maternal selective behaviour between the own and the alien dam, as reported in sheep (Terrazas et al., Reference Terrazas, Nowak, Serafin, Ferreira, Lévy and Poindron2002). To assess this question, we studied the response of 12-h-old kids given the choice between two anosmic mother goats, both of which displayed acceptance behaviours, since peripheral anosmia induced before parturition by irrigation of the nostrils with zinc sulphate prevents the development of maternal selectivity (Romeyer et al., Reference Romeyer, Poindron and Orgeur1994a; Hernández et al., Reference Hernández, Delgadillo, Serafin, Rodriguez and Poindron2004). Finally, we also investigated whether the size of the testing pen could influence the display of an early preference for the mother; we compared the performances of 8-h-old kids tested in a pen similar to that used for lambs, with that of kids tested in a smaller pen and placed closer to the dams at the beginning of the test.
Material and methods
General conditions of study and management before the test
The experiment was carried out in June 2000 and April 2001 at the experimental unit of the Institute of Neurobiology (UNAM), located on the grounds of the farm belonging to the Veterinary Faculty of the Autonomous University of Querétaro (Mexico). Different animals were used each year. The subjects were 87 kids (45 females and 42 males; 12 singles, 49 twins and 26 triplets) and their 47 multiparous Mexican mixed-breed mothers (dairy type). The animals were kept under intensive conditions at a density of approximately 10 m2 per mother, with a section of the pens provided with a sheltered area for protection from sun and rain. Animals were fed lucerne hay, minerals and concentrate pellets (18% crude protein; Generaleche, Purina®, Mexico) according to their gestation status, and water ad libitum. Oestrus was synchronised in the does using vaginal sponges (fluorogestone acetate, 45 mg; Intervet®, Holland) for 19 days, followed by an injection of pregnant mare serum gonadotropin (Folligon, Intervet®, 300 IU intramuscular). Females were fecundated by active bucks introduced in the group of females at the time of sponge withdrawal.
From gestation day 148 (mean duration of pregnancy: 150 days), animals were kept under supervision from 0600 to 2200 h. Only kids with a known exact time of birth were used. When a goat showed signs of imminent birth, a 2 m× 2 m individual pen was built where the female was located and the goat placed in it. She remained in it until 4 h after parturition to allow adequate development of the mother–young relationship. After this time, mothers were transferred to another corral, together with other mothers and their kids until the time of testing, so that interactions between unrelated females and kids could occur.
Experimental groups and testing procedure
To test the ability of kids to discriminate between their own mother and an alien equivalent one, we used a two-choice test paradigm that had already been used previously with success in lambs and kids (Lickliter and Heron, Reference Lickliter and Heron1984; Nowak et al., Reference Nowak, Poindron, Le Neindre and Putu1987; Addae et al., Reference Addae, Awotwi, Oppong-Anane and Oddoye2000; Awotwi et al., Reference Awotwi, Oppong-Anane, Addae and Oddoye2000). Kids were given the choice between their mother and an alien equivalent goat having given birth at about the same time, each mother being placed in one corner of a triangular (experiments 1 and 2, 1st and 2nd year) or rectangular (experiment 3, 2nd year) testing pen. The 1st year, the aim of the study was to test the ability of kids to discriminate between their mothers in three age groups (8, 12 and 24 h) and to investigate if their performance would be impaired by the experimental suppression of maternal olfaction before parturition, as reported in sheep (Terrazas et al., Reference Terrazas, Nowak, Serafin, Ferreira, Lévy and Poindron2002). In both species, pre-partum-induced anosmia induced surgically by olfactory bulb ablation, nerve severing or destruction of the olfactory mucosa by zinc sulphate prevented the establishment of exclusive nursing of the own young by its mother that relies on olfactory recognition by the dam (see Lévy et al., Reference Lévy, Keller and Poindron2004 for review). It is important to note that in the present paper, maternal selectivity refers specifically to this aspect of maternal recognition, which relies on maternal olfaction. Indeed, while anosmic ewes and goats are able to recognise their young in a two-choice tests by sight and/or hearing, they do not display rejection behaviours towards alien young that intent to suck (Ferreira et al., Reference Ferreira, Terrazas, Poindron, Nowak, Orgeur and Lévy2000; Poindron et al., Reference Poindron, Gilling, Hernandez, Serafin and Terrazas2003; Lévy et al., Reference Lévy, Keller and Poindron2004). Also, it is important to note that in the present study, in all cases the kids had full access to visual, acoustic and olfactory cues from the dams.
As the number of subjects studied in 2000 was only about 60% of the number originally planned (12 at 8 h, 7 at 12 h, 9 at 24 h with intact mothers and 11 at 12 h with anosmic mothers), the experiment was repeated the following year. The maintenance conditions and feeding of the animals, the location of living and testing pens, as well as the testing and observation procedures were the same in both years. In addition, results from the 1st year suggested that 8-h-old kids failed to show a preference for their mother. Since kids may be less active than lambs due to their hiding tendency, in the 2nd year we completed the initial groups and we constituted an additional group to verify whether the lack of preference could be due to the size of the testing pen being too large, associated with a low mobility of the kids. In this latter group, 8-h-old kids were tested in a smaller pen.
Discrimination of intact/selective mothers by their kids. Twenty-two kids (3 singles, 12 twins and 7 triplets) were tested 8 h after birth, 11 (1 single, 7 twins, 3 triplets) at 12 h and 18 (3 singles, 10 twins, 5 triplets) at 24 h, using different animals in each group, balancing the order of testing between the three groups over time as evenly as possible. The testing pen was located in a roofed building adjacent to the holding pens of the pregnant and post-parturient animals. The tested subjects could only see the two mothers in the testing pen, but acoustic isolation from the rest of the flock was not possible. The testing pen consisted of a triangular area (6 m × 6 m × 6 m), delimited by a 1-m-high metal barrier (Figure 1). Thirty minutes before the test, the two stimulus mothers to be used in the test and their kids were introduced in the central part of the testing pen for habituation, and remained there until the time of test. The alien mother was an unrelated female from the same living pen, having given birth at most 24 h before the mother of the tested kid. For the test, all kids except the one to be tested were removed and kept in a pen outside the building and out of the mothers’ sight. The two mothers were placed in their respective holding zone by one experimenter (Figure 1), the side of the own mother being alternated between tests and unknown to the two observers recording the behaviour of the kid. The kid to be tested could not see in which pen its dam was placed. The holding pens for the mothers were made of open-work metal panels allowing the animals fully to see and hear each other, as well as smell each other when at close quarters. Also in all cases, even though the exact number of vocalisations of the two stimulus mothers could not be recorded, it was noted that both goats were repeatedly bleating during the test, thus ensuring that kids had access to vocal cues. The kid was then placed in the starting zone of the pen and kept there for 30 s, so it could see and hear the does, before being released for the 5-min test. Two trained observers, using preformatted recording sheets and chronometers, recorded the following four variables. (a) Latency to leave the waiting zone (s). If it took the kid longer than 30 s to leave this zone, it was gently pushed out with a broom and given a 31-s latency. (b) Latency to reach a mother: time at which the kid first reached a contact zone after leaving the waiting zone. Kids that did not reach a contact zone during the test were given a latency of 301 s and not included in the analyses of this variable. (c) Identity of mother (left or right) that the kid reached first. (d) Time spent in each contact zone (s). The kid was considered to be in a given zone when it has at least the two front feet in that zone.
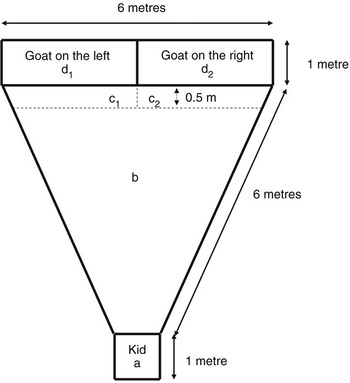
Figure 1 Large pen used for maternal discrimination tests in 8- to 24-h-old kids: (a) starting area for the kid; (b) neutral zone; (c1 and c2) zones of contact with the mothers.
The proportions of kids leaving the waiting pen or reaching a mother, as well as the latency to leave the waiting pen, provided information on the general activity of the kids and was analysed in all kids. The proportion of kids that reached their mother first and the time spent near the mothers gave indications about their discrimination ability and were analysed only in kids that reached at least one dam during the test.
Discrimination of anosmic/non-selective mothers by their 12-h-old kids. Nineteen 12-h-old kids (2 singles, 11 twins, 6 triplets) born from anosmic goats were tested in the pen shown in Figure 1. In this group, each kid was given the choice between its own and an alien equivalent mother, both of which had been rendered anosmic 1 year before this study by irrigation of the nostrils with a 1.5% solution of zinc sulphate. It is well documented that in ewes and does, anosmia induced by zinc sulphate during pregnancy prevents the establishment of olfactory recognition of the neonate by its mother and the development of selective maternal care. As a consequence, in both species, anosmic mothers display maternal acceptance towards an alien young as well as towards their own young (Romeyer et al., Reference Romeyer, Poindron and Orgeur1994a; Hernández et al., Reference Hernández, Delgadillo, Serafin, Rodriguez and Poindron2004; Lévy et al., Reference Lévy, Keller and Poindron2004). The goats had been rendered peripherally anosmic by irrigation of the olfactory mucosa with a 1.5% zinc sulphate solution 1 year before this study, according to the method of Romeyer et al. (Reference Romeyer, Poindron and Orgeur1994a). All females were anosmic and non-selective at the time of the test, as assessed by behavioural tests performed just after the two-choice tests (see Hernández et al., Reference Hernández, Serafín, Terrazas, Marnet, Kann, Delgadillo and Poindron2002 for details of anosmia and selectivity tests). In the present paper, we used the term anosmia, on the grounds that zinc-sulphate-treated animals were behaviourally anosmic to the food preference and maternal selectivity tests used to evaluate the efficacy of the treatment. For animal care and experimental treatments, including anosmia, guidelines A37801 for animal experimentation of the French Ministry of Agriculture were used. Anosmia did not induce observable changes in the general behaviour of the females, their body weight or the body weight of the kids. All testing procedures and recorded variables were the same as with intact mothers.
Influence of the size of the testing pen on maternal discrimination in 8-h-old kids. Seventeen 8-h-old kids (3 singles, 9 twins, 5 triplets) were tested in a pen that consisted of a rectangular corral 4 m wide × 3 m deep, the kids being placed at a distance of 1.5 m from the contact zones (Figure 2). As there was no waiting zone, the test started as soon as the kid was placed in the testing pen. Other variables were the same as for the other groups. Allocation of the 17 kids to the test small pen group and of the 10 kids tested the 2nd year in the large pen was balanced over time at the rate of about one kid tested in the large pen every 3rd test.
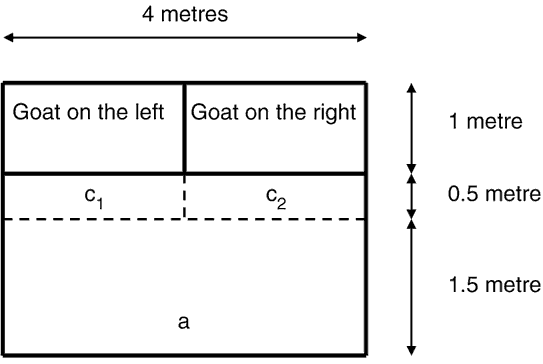
Figure 2 Small pen used for maternal discrimination tests in 8-h-old kids: (a) starting point for the kid; (c1 and c2) zones of contact with the mothers.
Data analysis
The results are presented as medians and quartiles. Within each group, data were compared with the Wilcoxon test, while the comparisons between groups were carried out with Kruskal–Wallis or Mann–Whitney U-tests. Between-group comparisons of proportions were carried out using the χ 2 and Fisher’s exact probability tests. The binomial test was used to test the hypothesis of a choice at random. Bilateral probabilities were used, except for the binomial test. Possible differences in performances due to sex of the kids or litter size were tested globally and within each group with Mann–Whitney U-tests. As no significant effects of these variables were found (P > 0.17 in all cases), these variables were not taken into account in further analyses. Statistics were carried out using the statistical package Systat 10 of Statistical Packages for the Social Sciences (2000).
Results
Preference for the mother on the 1st day of life
The proportion of kids that did not enter a contact zone was significantly higher in 8-h-old kids than in the two other groups (Table 1). Among the kids that reached a mother, the latency to reach a doe did not differ significantly between groups (Table 1). The proportions of kids that reached a mother but had to be pushed out of the starting area did not differ between groups (Table 1). In the groups of kids tested 8 and 12 h after birth, the proportion of kids that reached their mother first did not differ from a random choice (binomial test, P = 0.19 and 0.17, respectively). However, kids tested 24 h after birth reached their mother first more frequently than expected by chance (binomial test, P = 0.023). The proportion of first correct choice also differed significantly between 8- and 24-h-old kids (Fisher’s exact P = 0.045).
Table 1 Performances of kids in a 3-min two-choice test, when given the choice between their own mother and an alien equivalent mother. Kids were tested at 8, 12 or 24 h of age in a triangular 6 m× 6 m× 6 m pen or a rectangular 3 m× 3 m pen (8-h-old kids, short testing pen). Twelve-hour-old kids were tested with their own and an alien mother having both either their sense of smell fully functional (12-h-old kids, intact mothers), or impaired by pre-partum peripherally induced anosmia (12-h-old kids, anosmic mothers)

First column of results: a v. d + e: Fisher’s exact P = 0.036; a v. b: Fisher’s exact P = 0.09; c v. d, Fisher’s exact P = 0.10.
Second column of results: no significant differences between groups (χ 2 = 1.83, 2 d.f., P = 0.40).
Third column of results*: different from a choice at random (binomial test, P = 0.023); a v. b: Fisher’s exact P = 0.045.
Fourth column of results: values are medians with inter-quartiles in brackets. Latencies did not differ between groups (Kruskall–Wallis, P > 0.10).
For kids that reached a mother, the total time spent in the contact zone near the two mothers did not differ significantly between the three groups (Kruskal–Wallis H = 1.16, P = 0.56). In 8-h-old kids, there was no significant difference between the time spent near their own v. the alien mother (Wilcoxon z = 0.0; P = 1). In contrast, kids tested 12 or 24 h after birth spent significantly more time near their mother than near the alien dam (P < 0.05; Figure 3). The time spent near the own mother or near the alien dam failed to differ significantly between the three groups (Kruskal–Wallis tests, P ⩾ 0.11). Nonetheless, the percentage of time in the two contact zones c1 and c2 (Figure 1) that the kid spent near their own mother did so (8 h: 32% [0–93]; 12 h: 100% [38–100]; 24 h: 95% [56–100]; Kruskal–Wallis H = 6.32, P = 0.04). The group of 8-h-old kids differed from the two others (Mann–Whitney U-tests, P < 0.04).
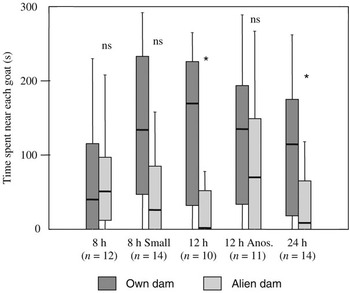
Figure 3 Box plot representation (median, lower and upper quartiles, whiskers) of the time spent by kids next to their own mother or an alien equivalent dam at 8, 12 or 24 h of age in a 5-min two-choice test. Small = test in small pen. Anos. = test with anosmic mothers. ns = P ⩾ 0.14 and * = P < 0.05 for the difference in the time spent with each mother (Wilcoxon). Only the data of kids that reached a mother during the test are included (see proportions in the text).
Preference for the mother by 12-h-old kids when both dams were anosmic and non-selective
The proportion of 12-h-old kids tested with anosmic mothers that did not reach a dam during the test tended to be higher than for kids tested with intact mothers, even though they were gently pushed with a broom (Fisher’s exact P = 0.10; Table 1). Among kids thta reached a goat during the test, the latency to reach a doe did not differ from that found in kids tested with intact mothers (Table 1). The proportion of these same kids that had to be pushed out of the starting area did not differ from that found in kids tested with intact dams (Fisher’s exact P = 0.39) and the proportion of kids reaching their own mother first did not differ from a random choice (binomial test, P = 0.27; Table 1). The total time spent by kids in the contact zones near the two dams did not differ from that found for kids tested with intact mothers (226 s [191.5–249.5]; Mann–Whitney U = 36, P = 0.18). Also, the time spent near their own mother (135 s [33.5–193.5]) did not differ significantly from that spent near the alien dam (70 s [0–149]; Wilcoxon z = 0.71, P = 0.48), contrary to the results of kids tested with intact mothers (Figure 3). Nevertheless, none of the studied variables differed significantly between the group of kids tested with two intact mothers and the group tested with two anosmic mothers (Mann–Whitney U-tests, P ⩾ 0.18 in all cases).
Preference for the mother by 8-h-old kids in a small pen
The proportion of kids reaching a mother tended to be higher in the small pen than in the large one (Fisher’s exact P = 0.09; Table 1). The latency taken by kids to reach a mother did not differ significantly from that found in the large pen (Mann–Whitney U = 67, P = 0.38; Table 1). For those kids that reached a mother, the proportion of them reaching their mother first still did not differ from a random choice (binomial test, P = 0.60) or from the proportion found in the large pen (Fisher’s exact P = 0.45; Table 1). Nonetheless, the total time spent with the two dams tended to be higher for kids tested in the small pen (209 s [156–255], v. 110 s [72–212], Mann–Whitney U = 48.5, P = 0.07). Also, kids that reached a dam tended to spend more time near their mother in the small pen (134 s [47–233] v. 40 s [0–115.5]; Mann–Whitney U = 48.5, P = 0.07), whereas this was not the case for the time near the alien dam (26 s [0–85] v. 51 s [12–97]; Mann–Whitney U = 66, P = 0.35). Besides, in the small pen, there was no statistical difference between the time kids spent with their own mother and with the alien doe (own: 134 s [47–233]; alien: 26 s [0–85]; Wilcoxon z = −1.48, P = 0.14; Figure 3). However, the percentage of time spent near the own mother relative to the total time spent near the two stimulus females was significantly higher than for kids tested in the large pen (84% [45–100] v. 32% [0–93]); Mann–Whitney U = 45.5, P = 0.04).
Discussion
Three main conclusions can be drawn from the results of the present study. First, kids can already display a preference for their own mother at 12 h of age in a two-choice situation, whereas this does not seem yet to be the case at 8 h for a majority of kids, even when they are placed relatively close to the does (2 m). Second, the fact that discrimination is impaired when kids are tested with two anosmic and non-selective dams suggests that the behaviours characterising maternal acceptance play some role in the initial preference shown by 12-h-old kids for their mother. Third, these results do not support the hypothesis that the time at which early recognition is established differs between newborn lambs and kids, despite the fact that lambs are followers and kids are hiders.
The results obtained in 12- and 24-h-old kids indicate without ambiguity that kids are able to discriminate their mother at those ages, even though they only do so at close quarters. This is at variance with the findings of Addae et al. (Reference Addae, Awotwi, Oppong-Anane and Oddoye2000) and Awotwi et al. (Reference Awotwi, Oppong-Anane, Addae and Oddoye2000) in dwarf African goats. The most likely explanation for this difference is that the slow learning abilities mentioned for the mothers in their studies (Addae et al., Reference Addae, Awotwi, Oppong-Anane and Oddoye2000) also applied for the kids. The possibility that some differences in breed exist between the dwarf African goat and the type of goats we used is also supported by the response of the animals to separation. Indeed, both in the present experiment and in a previous study of the recognition of the kid by the mother in the same type of goats (Poindron et al., Reference Poindron, Gilling, Hernandez, Serafin and Terrazas2003), animals were highly responsive to the testing situation, despite the fact that mothers and kids had been separated from each other only for a few minutes before the test. In contrast, in the studies of Addae et al. (Reference Addae, Awotwi, Oppong-Anane and Oddoye2000) and Awotwi et al. (Reference Awotwi, Oppong-Anane, Addae and Oddoye2000), animals did not display sufficient motivation in the test with less than 2 h of separation.
Whether kids are able to discriminate their mother before 12 h of age is not clear. The high proportion of kids that failed to reach a mother at 8 h in the large pen may suggest that discrimination is not yet established at that time, but it is also possible that this poor performance is related to the hiding behaviour of the kids. Thus, the tendency for more kids to reach a mother (regardless of her identity) in the small pen than in the large pen and to spend more time with both dams suggests that seeing the dam from a short distance is important to elicit an approaching response in kids. In addition, testing kids in a small pen seemed to facilitate the display of a preference for those kids that did reach a mother, as these kids tended to spend more time with their own dam than with the alien one. It is possible that because of their hiding behaviour, neonates do not have a strong motivation to walk towards a mother from a distance of several metres. Also, the improvement of the discrimination performances of the kids during the 1st day of life could depend on their motor activity, which at 8 h may still be insufficient for this type of test. Nonetheless, it must be noted that for kids that did reach a mother, latencies to do so did not differ depending on the age of the kids, suggesting that more than motor ability it is the response to the presence of the dams that may change with age. Besides, the criteria used in the present experiment and the experimental testing situations may not have been the most adequate to evidence this discrimination in kids of that age. Hence, we cannot exclude the possibility that 8-h-old kids may discriminate between their own and an alien doe at this early age. Additional studies including other criteria of a preference for the mother in very young kids would help clarify this point.
The impairment of the preference for the own mother in 12-h-old kids, given the choice between two anosmic dams, is similar to the results reported for lambs (Terrazas et al., Reference Terrazas, Nowak, Serafin, Ferreira, Lévy and Poindron2002). The behaviour of anosmic does was not monitored in the present study during the test or at parturition. Therefore, it cannot be excluded that the impairment of the performance of kids tested with anosmic mothers may be partly caused by some early disturbance of maternal behaviour, which in turn would affect the development of the kid’s preference, rather than by the behaviour of the goats during the test. Further studies are warranted to clarify this and it would be particularly interesting to verify whether the latency to first nursing and early frequencies of suckling are affected by anosmia, given the important role of the first suckling activity of the neonate for its bonding to the dam in sheep (Nowak et al., Reference Nowak, Murphy, Lindsay, Alster, Andersson and Uvnäs-Moberg1997). Nonetheless, in sheep, the results of Terrazas et al. (Reference Terrazas, Nowak, Serafin, Ferreira, Lévy and Poindron2002) show that if both the lamb’s own mother and an alien mother are anosmic, they do not behave differently. Furthermore, it is well established that anosmic goats display similar acceptance behaviours towards alien kids as intact goats do towards their own kid (Romeyer et al., Reference Romeyer, Poindron and Orgeur1994a; Hernández et al., Reference Hernández, Delgadillo, Serafin, Rodriguez and Poindron2004). Therefore, our present results suggest that newborn kids first develop a preference for behaviours indicative of maternal acceptance, before developing the ability to recognise individual characteristics of their dam, as it is the case in lambs. However, this phase of preference for acceptance behaviours is probably of short duration, since 24-h-old kids already chose their dam from a distance, which presumably depends on the recognition of individual visual and/or acoustic maternal cues. Which behaviours of the mother may serve as a basis for discrimination has not been investigated in the present study. Results in sheep indicate that maternal vocalisations (Terrazas et al., Reference Terrazas, Nowak, Serafin, Ferreira, Lévy and Poindron2002), the motor behaviour of the female and her posture during the test (Nowak et al., Reference Nowak, Breton and Mellot2001) may serve as a basis for discrimination. Also, studies in sheep indicate that this early preference of the neonate for its mother depends on a learning process, rather than on a spontaneous preference for some key stimuli from the ewe (Nowak et al., Reference Nowak, Murphy, Lindsay, Alster, Andersson and Uvnäs-Moberg1997; Val-Laillet et al., Reference Val-Laillet, Simon and Nowak2004). While the sensory cues used by the neonate to make its choice remain to be identified, it is clear that olfaction does not play a major role in the present two-choice test situation. If this had been the case, kids would have been able to discriminate their mother from the alien doe on the basis of olfactory cues, regardless of the fact that both dams displayed acceptance behaviours as a consequence of maternal anosmia.
As a whole, the results of the present study do not support the hypothesis that newborn kids are slower in developing an early preference for their mother than lambs. Therefore, the following or hiding behaviour of the neonate does not seem critical in that respect. Two main factors may explain this lack of difference. First, the pressure to establish rapidly a preferential mother–young bond ensuring proper maternal investment is probably similarly high in the two species. Indeed, sheep and goats are both gregarious and present seasonal synchronised breeding leading to a high probability of numerous births concentrated over a short period. Second, in both species there is an initial phase of intense interacting between the mother and her neonate(s) that lasts several hours and with a similar behavioural repertoire and pattern in sheep and goats (Hersher et al., Reference Hersher, Richmond and Moore1963). Therefore, factors possibly common to the two species, other than the hiding or following strategy of the neonate, are certainly involved during this early period. For example, a very general factor that may stimulate early learning in the neonate is the experience of labour itself, which has recently been found to facilitate early olfactory exposure learning in human babies, possibly through the stimulation of catecholamines release (Lagerkrantz, Reference Lagerkrantz1996; Wang et al., Reference Wang, Zhang and Zhao1999; Varendi et al., Reference Varendi, Porter and Winberg2002). Another important factor for the establishment of early preferences is the sucking activity of the neonate (Hall, Reference Hall1987). In particular, in the lamb the development of the initial preference for the mother depends on an associative process involving the first nursing bouts (Val-Laillet et al., Reference Val-Laillet, Simon and Nowak2004). Therefore, it would be worthwhile to investigate whether association during early nursing also occurs in kids, given the similarity of the immediate post-partum care of the neonate in the two species.
Acknowledgements
G. Gilling was supported by a scholarship from CONACyT (144835). We thank DGEP and PAPITT (ES212796) for their financial support, the Universidad Autónoma de Querétaro for housing the animals and Raúl and Francisco Paulin for taking care of the animals. We are grateful to Professor Dwight Krehbiel for correction of the English in the final version of the manuscript.




