



INTRODUCTION
The development of ethnobotanical studies in pre-Hispanic Central America has been slower than other regions, despite important foundational research completed at sites from southern Mexico through Panama. Critical work has been accomplished on the origins and spread of cultigens such as maize (Perry et al. Reference Perry, Dickau, Zarrillo, Holst, Pearsall, Piperno, Berman, Cooke, Rademaker and Ranere2007; Pickersgill Reference Pickersgill2007; Piperno Reference Piperno2009; Ranere et al. Reference Ranere, Piperno, Holst, Dickau and Iriarte2009), the relationship between plants and artifact use (Lentz et al. Reference Lentz, Beaudry-Corbett, Reyna de Aguilar and Kaplan1996; Piperno and Holst Reference Piperno and Holst1998), the importance of root crops in ancient societies (Cagnato and Ponce Reference Cagnato and Ponce2017; Piperno Reference Piperno and Zeder2006; Piperno et al. Reference Piperno, Ranere, Holst and Hansell2000; Sheets et al. Reference Sheets, Dixon, Guerra and Blanford2011; Simms et al. Reference Simms, Berna and Bey2013), agricultural practices and landscape (Baldi Reference Baldi2011; Dickau Reference Dickau, Van Derwarker and Peres2010; Lentz et al. Reference Lentz, Magee, Weaver, Jones, Tankersley, Hood, Islebe, Ramos Hernandez, Dunning, Lentz, Dunning and Scarborough2015; Matthews Reference Matthews1984; Piperno Reference Piperno, Bush, Flenley and Gosling2007; Rue Reference Rue1987, Reference Rue1989), and forest management and ecology (Anchukaitis and Horn Reference Anchukaitis and Horn2005; Dussol et al. Reference Dussol, Elliott, Michelet and Nondédéo2017; Goldstein and Hageman Reference Goldstein, Hageman, Staller and Carrasco2009; Lentz et al. Reference Lentz, Ramírez and Griscom1997, Reference Lentz, Magee, Weaver, Jones, Tankersley, Hood, Islebe, Ramos Hernandez, Dunning, Lentz, Dunning and Scarborough2015; McNeil et al. Reference McNeil, Burney and Burney2010; McNeil Reference McNeil2012). In recent years, researchers have used new approaches that greatly expand the potential for recovery of evidence of plants and their uses (Anchukaitis and Horn Reference Anchukaitis and Horn2005; Hageman and Goldstein Reference Hageman and Goldstein2009; Kraft et al. Reference Kraft, Brown, Nabhan, Luedeling, Ruiz, d'Eeckenbrugge, Hijmans and Gepts2014; Lentz et al. Reference Lentz, Magee, Weaver, Jones, Tankersley, Hood, Islebe, Ramos Hernandez, Dunning, Lentz, Dunning and Scarborough2015; McClung de Tapia Reference McClung de Tapia1985; McNeil et al. Reference McNeil, Burney and Burney2010; Morehart et al. Reference Morehart, Lentz and Prufer2005; Morell-Hart Reference Morell-Hart2011; Morell-Hart et al. Reference Morell-Hart, Joyce and Henderson2014; Pickersgill Reference Pickersgill2007; Piperno Reference Piperno2009; Piperno et al. Reference Piperno, Ranere, Holst and Hansell2000; Sheets et al. Reference Sheets, Dixon, Guerra and Blanford2011, Reference Sheets, Lentz, Piperno, Jones, Dixon, Maloof and Hood2012; Whitney et al. Reference Whitney, Rushton, Carson, Iriarte and Mayle2012).
These advances bring us to a point where we can begin to understand ethnoecology (Conklin Reference Conklin1954), defined here as the dynamic relationship between affordances provided in a botanical landscape and the impacts of human activities on that botanical landscape. This paper uses the case study of Puerto Escondido, located in the lower Ulúa River valley of Caribbean coastal Honduras, to demonstrate how we can explore ethnoecology in Central America. It builds on the effectiveness of using multiple methods in concert to interpret ethnobotanical practice in the past (Morell-Hart Reference Morell-Hart2011; Morell-Hart et al. Reference Morell-Hart, Joyce and Henderson2014). By examining chipped-stone tools, ceramics, sediments from artifact contexts, and macrobotanical remains recovered from Puerto Escondido (Henderson and Joyce Reference Henderson and Joyce1998, Reference Henderson, Joyce and Emery2004a, Reference Henderson and Joyce2004b; Joyce and Henderson Reference Joyce and Henderson2001, Reference Joyce and Henderson2002, Reference Joyce, Henderson, Laporte, Arroyo, Escobedo and Mejía2003, Reference Joyce and Henderson2010, Reference Joyce, Henderson, Blomster and Cheetham2017), research on botanical practices “in the home,” such as foodways, medicinal practices, fiber crafting, and ritual activities, can be complemented by studies of practices “close to home,” such as agricultural and horticultural practices, forest management, and other engagements with local and distant ecologies. Such complementary studies grant us a broader sense of the ways that people negotiated with the world of plants in which they were embedded.
Multiple recovery methods produced a large inventory of plants from Puerto Escondido that no single method would have recovered, as several key plants are identifiable through only one of the available methods. A wide set of ethnobotanical practices is represented by the recovered residues, including food preparation and consumption, fiber and thatch processing, medicinal activities, and ritualized practice. This broad range of ethnobotanical activities indicates a rich knowledge of the surrounding landscape, and possession of skills associated with the collection, management, and cultivation of diverse plant species. Furthermore, such activities would have directed the ways that residents of Puerto Escondido molded the landscape around them to provide these resources, including water management, replanting select species in homegardens, and soil fertilization and fallowing.
Previous research at Puerto Escondido has already demonstrated the use of early pottery bottles and bowls to contain cacao-based foods (Henderson and Joyce Reference Henderson, Joyce and McNeil2006; Henderson et al. Reference Henderson, Joyce, Hall, Hurst and McGovern2007; Joyce and Henderson Reference Joyce, Henderson, Staller and Carrasco2009). The current study shows that such early bowls contained an even wider range of foods, including maize and squash. Obsidian blades provide more evidence of use in processing fibers and wood, and relatively less use in food plant processing. Nonetheless, our analysis of obsidian blades shows the use of arrowroot, manioc, maize, and chile peppers at an early period. Such plants are virtually unrecoverable or frequently unidentifiable in macrobotanical residues recovered through flotation in this region.
Obsidian blades and flakes from later periods (after a.d. 200) also show more evidence of use in fiber and wood processing than in food plant processing. Microbotanical remains in the surrounding sediments from which these objects were recovered reveal a much wider range of food plants, including arrowroot, hackberry, bean, and maize, suggesting that the obsidian tools were being used primarily in non-culinary processing but discarded in locations where other plants, including food scraps, were deposited. Macrobotanical remains of plants recovered from trash-filled pits dating to this later period attest to the prevalence of maize and beans, as well as the presence of several other food taxa. Considering the botanical assemblage present at the site as a whole, what is most striking is the many different plants that are evident, representing uses beyond culinary consumption.
Without recovering residues from ceramics, obsidian tools, and adhering sediments, the true breadth of plant use at Puerto Escondido would not have been evident from the macrobotanical remains alone. No single method employed would have detected all the plants identified using the suite of methods used in this study. Moreover, plants that participated in multiple kinds of practices would not have been visible across artifacts and contexts. The evidence obtained from the direct examination of artifacts and their adhering sediments provides a basis for understanding not just which plants were present, but how assemblages of tools were used to process different plants and how different plants were processed using a variety of tools. Researchers working in areas where plant remains are detected using only one method may want to broaden the gamut of methods employed—even if some additional methods can only be applied to samples of limited size—in order to arrive at richer plant inventories. Sampling residues from artifacts is critical, as even the most widely accepted expectations about tool use may be based on ethnographic analogues. These analogues best represent recent plant practices which may be different from earlier practices that have dramatically narrowed or radically shifted over time (Perry Reference Perry2005).
Based on this case study, we advocate a move toward socially foregrounded and broad-based ethnobotanical approaches, where research is framed from the beginning in terms of a wide scope of plant uses and explicit definition of the kinds of practices that would have engaged humans and plants in complicated relationships. We necessarily draw on ethnographic and ethnohistoric analogues to consider likely plant practices at this ancient community, and infer impacts of these practices on the landscape based on the preferred growing conditions of various plants. By contributing to a broader understanding of foodways, medical practice, construction of homes, and ethnoecology, analysis of ancient plant remains can become an even more important contribution to understanding human communities in the past, as well as their impacts on surrounding botanical communities.
PUERTO ESCONDIDO: ARCHAEOLOGICAL RESEARCH, SAMPLE CONTEXTS, AND LOCAL ECOLOGY
Archaeological research at Puerto Escondido began in 1993, when employees of the Honduran Institute of Anthropology and History (Instituto Hondureño de Antropología e Historia, or IHAH) received reports that bulldozing for a new housing development had disturbed a human burial containing ceramic vessels and a group of obsidian nodules. Preliminary recording of bulldozer-cut road faces by IHAH employees led the IHAH to ask Henderson and Joyce to focus research in 1994 on this site, as mitigation before construction continued. Fieldwork carried out during six- to eight-week seasons from 1994 to 1997 demonstrated that what was initially reported as a Late Classic (a.d. 500–1000) settlement actually was a multi-component site with occupation beginning in the Early Formative period (Henderson and Joyce Reference Henderson and Joyce1998). A continuous sequence of occupation from before 1600 b.c. through to a.d. 650 to 750 was defined based on analysis of 42 calibrated radiocarbon dates reported in previous publications (Joyce and Henderson Reference Joyce and Henderson2001, Reference Joyce and Henderson2007, Reference Joyce and Henderson2010, Reference Joyce, Henderson, Blomster and Cheetham2017). Ceramics collected from surface deposits indicate that occupation actually continued later, to ca. a.d. 1000, in contexts destroyed by bulldozing.
Following a break in fieldwork caused by Hurricane Mitch, which flooded the IHAH repository where collections were stored, a final season of excavation at the site in 2001 aimed to investigate the extent of early village occupation through test excavations of a stratified random sample of zones of the bulldozed site (Early and Middle Formative periods, through 400 b.c.). Analyses of the stratigraphic sequence of architecture from this early period (Joyce Reference Joyce and Beck2007), burials (Joyce Reference Joyce, Adams and King2011), and the execution of symbolic designs in ceramics and stone sculpture (Joyce and Henderson Reference Joyce and Henderson2002, Reference Joyce, Henderson, Laporte, Arroyo, Escobedo and Mejía2003, Reference Joyce, Henderson, Blomster and Cheetham2017) demonstrated that Puerto Escondido was a village that participated in the use of Olmec-related symbolism and practices (Joyce and Henderson Reference Joyce and Henderson2010), with differentiation among residents in different sectors of the village. This included differences in the use of imported raw materials such as jade and obsidian (Joyce et al. Reference Joyce, Shackley, McCandless and Sheptak2004) and luxury items made from locally available marble (Luke et al. Reference Luke, Joyce, Henderson, Tykot and Lazzarini2003).
The study of plant remains was part of the project from its beginning. In 1994, when the deposits investigated dated to the late Middle Formative through Classic periods (400 b.c.–a.d. 800), Margie van de Guchte undertook flotation of samples and established protocols which continued to be used through 1997, calling for collection of 2-L sediment samples. During the 2000 field season, Natasha Lyons reviewed field procedures for collection and processing of sediment samples for flotation. Over these multiple seasons, the volume of sediment processed for individual samples ranged from 2 to 10 L. Many features were of small overall volume and thus yielded smaller volumes of sediment for flotation.
Lab processing of samples and identification of plants began in 2000, when Cane (Reference Cane2001) analyzed flotation samples from 32 excavation units processed in 1995. As part of her doctoral dissertation research, Morell-Hart (Reference Morell-Hart2011) included data from these 32 flotation samples, and also conducted research to recover phytoliths and starches from six pieces of worked obsidian. In postdoctoral research, Morell-Hart undertook analysis of an additional group of artifacts, including five additional chipped obsidian objects (for a total of 11 obsidian artifacts analyzed) and four ceramic sherds. In addition to these macrobotanical and microbotanical analyses, a sample of ceramic sherds was tested for the presence of chemical residues potentially indicative of the presence of cacao-based foods (Henderson and Joyce Reference Henderson, Joyce and McNeil2006; Henderson et al. Reference Henderson, Joyce, Hall, Hurst and McGovern2007; Joyce and Henderson Reference Joyce and Henderson2007, Reference Joyce, Henderson, Staller and Carrasco2009). These analyses led to the proposal that early use of cacao fruit involved the production of fermented drinks.
The archaeological contexts for recovered plant remains at Puerto Escondido date as early as the Ocotillo phase (1400–1100 b.c.) based on 16 radiocarbon determinations (Joyce and Henderson Reference Joyce and Henderson2007:Table 1) through the early Ulúa phase (a.d. 450–650) based on five radiocarbon determinations (Joyce and Henderson Reference Joyce and Henderson2007:Table 1). Samples come from four out of five sectors in the site that were extensively excavated (Operations 2/8, 4 North, 4 South, and 6; Table 1). Three of these sectors (Operations 4 North, 4 South, and 6) can be correlated with one of the two large earthen mounds truncated by bulldozing in 1993, and the fourth sector (Operation 2/8) is located where the second, somewhat smaller, mound once stood. In each excavation area, the bulldozer-cut surface left parts of features visible below a thin layer of redistributed sediments. The features the bulldozer truncated included thick clay walls of rectangular structures (in Operation 2/8, often with stone features), postholes, hearths, subsurface bell-shaped pits, and human burials (in Operation 2/8, two stone cist graves), indicating that the tested areas were used for dwelling during their most recent preserved periods of occupation. The sequences of features of this latest occupation level from which plant remains were sampled dated to between a.d. 200 and 650.
Table 1. All samples (all types), ordered by area of site and time period.

Preceding this sequence of residential remains in all excavated areas were sequences of buried ancient surfaces which yielded plant remains. Blocks were excavated to depths of as much as 3.5 m, in one location in Operation 4 North, until there were no new traces of cultural materials or constructions. In this area, buried surfaces starting at depths of 2.2 m revealed a sequence of wattle-and-daub building construction and remodeling (Joyce Reference Joyce and Beck2007). Early wattle-and-daub buildings were replaced by more substantial, more permanent buildings, with thick clay wall foundations supporting wattle-and-daub superstructures, with the use of stone for platforms in one area, and plaster floors in another. In two of the sectors providing samples discussed in this paper (Operations 2/8 and 4 North), such residential buildings were replaced after about 900 b.c. by wide earthen platforms with a stepped profile or stairway with stone facing (Operation 2/8) or with traces of plaster surfacing (Operation 4 North). Excavations within the latter construction identified multiple buried vessels, one containing jade ornaments, and remains of at least two human burials.
The excavated areas of the site did not produce identifiable features dating to the period between 200 b.c. and a.d. 200, although pottery recovered in mixed fills can be identified as pertaining to types made during those centuries. Instead, there is an apparent discontinuity in use of space with the building of new groups of residential structures that were truncated by bulldozing. In Operation 2/8, a larger building with a built-in bench dating to the period after a.d. 200 may be a new kind of public structure. Buildings in all areas occupied after a.d. 200 (Operations 2/8, 4 North, and 4 South) are oriented at right angles to each other, framing open patios or yards. Hearths and subterranean storage pits are located in the spaces surrounding the houses. Burials were located along the walls of the houses and occasionally in reused storage pits in the yards around houses. One burial in Operation 4 North contained a very elaborate assemblage (Joyce Reference Joyce, Adams and King2011).
This pattern of deep stratigraphy, with multiple remodeling events creating episodic depositional histories for mainly residential structures, provides a very productive setting for the exploration of plant use. Samples explored in this study came from a number of well-defined depositional contexts and features (Table 1). The earliest samples (dating 1400–1100 b.c.) include obsidian chipped-stone artifacts examined for starch and phytoliths and sherds tested for the presence of theobromine. These come from two areas (Operations 4 South and 6), both with well-preserved remains of thick clay building walls forming rectangular rooms presumed to be residential construction. Samples of ceramics tested for theobromine come from one of these areas (Operation 4 South) in sequences of remodeling that six radiocarbon dates suggest took place between 1150 and 900 b.c. (Joyce and Henderson Reference Joyce and Henderson2007:Table 1).
Samples of ceramics assigned to the period from 900–700 b.c. were tested for phytoliths and starches. These samples all come from Operation 4 North, where a house with thick clay wall bases supporting wattle-and-daub upper walls was burned, apparently deliberately, and filled in as part of the building of a large platform used for human burials and burial of cached pots. Before the burned walls of the house collapsed inward, a group of bowls and bottles were broken and their sherds deposited on the interior floor. Six radiocarbon dates suggest this event took place around 900 b.c. (Joyce and Henderson Reference Joyce and Henderson2007:Table 1). The sherds sampled are typical of vessels produced beginning around 1100 b.c. The botanical remains recovered from these sherds should be differentiated from other samples, an obsidian flake and flotation samples, recovered from contexts that post-date these events. One flotation sample (4AW-12) represents the contents of one of the pots placed in the newly constructed platform (ca. 900–700 b.c.). Another flotation sample (4AM-13) came from the sediments around clay hearths postdating the platform. The third sample, an obsidian flake (4AW-19/20), came from the matrix between the platform and the level associated with the hearth. Both can be assigned to the period from 700 to 400 b.c. A final early botanical sample is represented by another sherd tested for theobromine, deposited in an area dated to 400–200 b.c. in the Operation 2/8 area (based on three radiocarbon dates; Joyce and Henderson Reference Joyce and Henderson2007:Table 1).
The remaining 36 samples, light fractions from flotation or residues from obsidian flakes and blades, come from the Operation 4 North area on the larger mound, in houses built much later over the platform, which dates to 900 b.c. These samples are derived from features associated with residential compounds with buildings laid out at right angles to each other around open spaces with hearth features, and large subterranean pits on other sides. The samples (Table 1) can be divided into two groups: one dating from the late Chamelecon phase (a.d. 200–450; 22 samples) the other from the early Ulúa phase (a.d. 450–650; 14 samples), with dates based on eight radiocarbon samples (Joyce and Henderson Reference Joyce and Henderson2007:Table 1). These samples represent two intervals in the continuing history of residential occupation of one area of the village of Puerto Escondido.
In terms of affordances offered by the local ecology of Puerto Escondido, there would have been a fairly wide range of plants to collect, manage, and cultivate. Henderson and Joyce (Reference Henderson, Joyce and Emery2004a:3) have even hypothesized that “high soil fertility and habitat diversity…were among the factors that allowed a large agricultural population to live in rural farming settlements in situations of relative affluence.” The primarily riverine ecology of this ancient community was marked by some vertical differences in microenvironments, as well as seasonal variations in hydrology. Broadly, the vegetation in the immediate region is marked by tropical rainforest and wet tropical highlands (Marshall Reference Marshall2007). The riverine zone is characterized by highly agriculturally productive soils, similar to those of the lower Usumacinta in Guatemala, and the area encompasses fairly broad range of habitats (Pope Reference Pope and Robinson1987). Pope (Reference Pope and Robinson1987:115) identified the intensive use of alluvial and colluvial fan and upland soils in the Classic period, but noted less intensive use of the northern gallery forest and tributary plain soils. He suggested that the best soils may have been reserved for commercial use, or potentially avoided due to their propensity for flooding. Contemporary agricultural uses of land in this region have reduced overall taxa richness, but there is still a high number of plant species that continue to thrive in the region.
PLANT USE AT PUERTO ESCONDIDO: RESEARCH METHODS
The paleoethnobotanical data from Puerto Escondido encompass nine kinds of contexts and five kinds of spaces, consisting of both light fraction macrobotanical samples analyzed by Cane (Reference Cane2001), and microbotanical extractions from obsidian artifacts and ceramic sherds analyzed by Morell-Hart (Reference Morell-Hart2011). The methods and reasoning for laboratory subsampling are outlined below.
Lithic Subsampling Strategy
During excavation, every piece of obsidian was collected individually, bagged separately, and left unwashed unless selected for compositional or other analysis. The objective of subsampling lithic artifacts was to obtain plant residues related to tool use, provide complementary lines of evidence for contexts where cacao residues had been recovered, and maximize the overall quantity of recoverable plant material. After first selecting for particular contexts (e.g., time period orassociation with cacao residue vessels), the selection criteria were as follows, in order of highest preference first: (1) formal tool or fragment; (2) retouching evident; (3) obvious usewear; (4) large size; and (5) visibly dirty, with adhering sediment.
Where a preferred attribute was not available for a particular context, we skipped this attribute and went on to the next criterion. Criteria 1–3 were directed toward obtaining plant residues related to tool use, while criteria 4–5 were directed toward maximizing the amount of adhering sediment. We targeted plant use and human-environmental interactions through analysis of the residues from adhering sediments, and targeted specific tool uses through sonicated residues. A total of 10 obsidian artifacts were included in this analysis. Six obsidian artifacts were analyzed in the first set (first described in Morell-Hart Reference Morell-Hart2011), and four additional obsidian artifacts were selected for the second set, described for the first time here (Figure 1).

Figure 1. Obsidian artifacts selected for 2013–2014 analysis: (a) 4DC-130 flake fragment, (b) 4DK-136 flake fragment, (c) 6C-67 flake fragment, and (d) 4AW-19&20 retouched flake. Photographs by Morell-Hart.
Blade fragments analyzed came from lined-pit fills (04AH-17 and 04DZ-02) and high-density midden material (04DP-25) dating between a.d. 200 and 650, as well as from matrices between defined occupational surfaces (04F-06 and 04G-04). Two obsidian retouched flakes from a similarly dated deposit associated with a hearth (04AL-07) were also studied.
Early obsidian flake fragments studied come from two contexts (04DC-130 and 04DK-136) where cacao residues were chemically recovered from ceramics dating to between 1100 and 1000 b.c. A third flake fragment from a structure in a separate excavation area (06C-67) dates to the same period. These three flakes represent rapidly produced expedient tools, with no evidence of retouching. More effort appears to have been expended on a fourth retouched flake recovered from a deposit (04AW-19&20) dating between 700 and 400 b.c.
All of the lithic artifacts from which microbotanical residues were obtained are made from obsidian. Analysis of the techniques of manufacture, as part of a study of chemical composition of obsidian at the site over time (Joyce et al. Reference Joyce, Shackley, McCandless and Sheptak2004), showed that before 1100 b.c. obsidian was worked using bipolar reduction, producing flakes and distinctive cores from material mostly obtained within 100 km of the site. Some of the flakes were worked into blade-like forms. After 1100 b.c., prismatic blade production is represented, primarily in obsidian from the newly used Ixtepeque source. Over time, Ixtepeque obsidian and the prismatic blade industry increased to the majority of obsidian tools used at the site, but bipolar reduction of more local material never disappeared. All of the obsidian artifacts selected for analysis have evidence of being utilized, though some in a more specialized way and others in a more expedient way. As the results indicate, while differences in formality of tool production may be culturally significant in other ways, they do not provide a good basis from which to extrapolate assumptions about tool use in plant processing.
The laboratory extractions of residues from lithic tools involved standard protocols (as detailed in Morell-Hart Reference Morell-Hart2011; Morell-Hart et al. Reference Morell-Hart, Joyce and Henderson2014). Stone artifacts were first dry brushed, if sediments still visibly adhered. All artifacts underwent a wet wash to remove additional adhering residues, then a sonicating wash to remove embedded materials in the fissures and pores of the artifact. The dry wash and first wet wash were carried out using sterile non-powdered gloves (two unique sets per artifact; one unique set per sample). Both wet washes were carried out in a distilled water medium. The sonicating wash was carried out using a Baxter large capacity ultrasonic cleaner. In this process, artifacts were sonicated for 5 minutes, and only partially immersed in water to isolate the usewear portion of the artifact.
The effluvium from each of the 2–3 washes per artifact was concentrated using centrifugation, at 3,000 rpm for 5 minutes. The concentrated residues were mounted on slides and viewed under a polarizing 100–1000× Nikon microscope.
Ceramic Subsampling Strategy
All of the ceramics from which microbotanical residues were obtained were recovered from a single deposit extending at least 6 × 4 m on the floor of a demolished house. Reconstructable vessels were recovered from this very large deposit, indicating deliberate in situ smashing of whole vessels, likely the remains of a feasting event. Ceramics in the sealed ceramic deposit had a higher frequency of carved/incised iconography than contemporary ceramics elsewhere at the site. Rubble from the collapse of burned daub walls effectively sealed the context of the floor deposit. The vessels in the sealed deposit were primarily bowls, with some small bottles, and there were no traces of cooking or storage vessels.
The objective of subsampling ceramic artifacts was to identify potential use differences associated with ceramic types, identify plant use associated with feasting events, and maximize the quantity of residues recovered. As with the lithic artifacts, we targeted evidence of human-environmental interactions generally. We also sought to identify potential associations between plants and particular ceramic types. We first selected from one context associated with a feasting event, drawing all the sherds from this same locus. We also selected for the same vessel form, in this case flaring-wall bowls. Through this sampling strategy we hoped to identify the different and similar uses of a particular form of ceramic across ceramic types at a single feasting event. After selecting for these attributes, the criteria were as follows, in order of highest preference first: (1) type (not duplicated with others); (2) basal sherd where available, body/rim sherd where basal sherd unavailable; (3) large size; (4) interior surface intact; and (5) visible interior residues.
As with the lithic artifacts, where a preferred attribute was not available for a particular context, we skipped this attribute and went on to the next criterion. Criterion 1 targeted differences associated with type, while criteria 2, 3, and 5 maximized the potential for adhering residues, and criterion 4 reduced the possibility for paste/temper contamination during the extraction process. Ultimately, of the available collection, only four sherds proved suitable for analysis (Figure 2).

Figure 2. Ceramic sherds selected for analysis from context 4BQ-6 (1100–900 b.c.). From top to bottom: Fia Metallic Grey, Boliche Black, Bonilla Yellow-Brown, and Sukah Differentially Fired. Exterior (left) and interior (right). Photographs by Morell-Hart.
Laboratory extractions of residues from the ceramic samples were both complicated and simplified by the fact that all of the ceramics had previously been washed. For this reason, the ceramics were first wet-washed again to remove potential contamination, using a distilled water medium and independent sets of sterile powder-free gloves. The second wet wash, as with the lithic tools, involved sonication. In this case, however, a hand-held sonicating device was used, in place of a water bath sonicator. The sherds were positioned to maximize interior holding volume, filled with distilled water, then sonicated using a Junghoon Corp La Belle Ultrasonic cleaner (30–40 kHz; Figure 3). As with the lithic artifacts, the effluvium from each of the wet washes was concentrated using centrifugation at 3,000 rpm for 5 minutes. The concentrated residues were mounted on slides and viewed under a polarizing 100–1000× Nikon microscope.

Figure 3. Demonstrating method of ceramic sherd sonication using Junghoon Corp La Belle Ultrasonic cleaner. Photograph by Morell-Hart.
Macrobotanical Subsampling Strategy
The macrobotanical samples analyzed are light fractions from sediment flotations of 32 excavated loci (Table 1) collected during the 1995 field season, represent a total volume of approximately 64 L, and present a diverse selection of taxa (Table 2; Cane Reference Cane2001; Morell-Hart Reference Morell-Hart2011). Few taxa present could be identified due to poor preservation conditions, but botanical remains were classified as wood, parenchymous tissue, cupules from Zea mays, seeds, and other unidentifiable tissues. Samples were taken from selected loci based on the visible concentration of organic matter or association with particular features, such as burials and possible hearths. Sediment samples of 2 L were taken from most loci. Some samples consisted of the entire contents of a locus, whether it was greater or less than 2 L.
Table 2. All taxa ubiquities, tabulated by sample type. Minimum number of taxa represented = 23. Numbers do not indicate raw counts of taxa, but rather reflect the number of instances in which each taxon was recovered. An individual sample type from an individual locus is counted as a single instance.
The sediment samples were processed in the field using a simple hand-held flotation system. The light fraction was skimmed off the top of the water in the bucket with a sieve. Both the light and heavy fractions were dried on cloth squares. The corners of the cloth squares were tied to make small bags to store the contents of the light and heavy fractions. These light fractions are the focus of our analysis.
In the laboratory, samples were selected by Cane and Joyce for greater recovery of charred plant remains as well as to explore specific contexts. During sorting and identification, Cane divided the light fractions of the floated sediment samples according to particle size using geological sieves. Charred materials, and other materials such as bone or snails, were removed from the sample once identified under the microscope. Basic initial identifications designated wood, parenchymous tissue, seeds, and other materials. Subsequent identifications classified specimens according to more specific morphological attributes, generally including size, the shape of the macrobotanical specimen's “footprint,” surface patterning, and other related morphological characteristics such as presence or absence of testa. As is common with archaeobotanical specimens, morphologies were often drastically altered through taphonomic processes, complicating identification (Boardman and Jones Reference Boardman and Jones1990; Hubbard and al Azm Reference Hubbard and Azm1990; Smith and Jones Reference Smith and Jones1990; Stewart and Robertson Reference Stewart and Robertson1971). For this reason, many remains were left unidentified. Other unknown or unrecognized yet potentially diagnostic specimens were drawn and photographed by Cane, Morell-Hart, and several undergraduate student assistants, with the expectation that they may eventually be identified.
Specimens were compared with materials in the modern reference collections housed in the U.C. Berkeley Paleoethnobotany Laboratory, and with images in books (e.g., Lentz and Dickau Reference Lentz and Dickau2005; Martin and Barkley Reference Martin and Barkley1973) and online in various databases (e.g., Kew Royal Botanic Gardens 2011; United States Department of Agriculture 2011). Once identification was complete, surviving portions of the macrobotanical specimen and taphonomic transformations were noted, for additional clues as to potential processing regimens (Fosberg Reference Fosberg1960; Pearsall Reference Pearsall2008). After cataloging the macrobotanical specimens, Cane curated all removed materials in gelatin capsules and boxes, labeled with accompanying site and contextual information.
The greatest number and greatest diversity of organic remains came from locus 04DP-25, a deep pit filled with trash dating to a.d. 200–450, located beneath a later adult burial in the Operation 4 South area. A total of 334 parenchymous tissue fragments and 276 cupules were recovered from this locus. Unfortunately, the condition of materials in 04DP-25 prohibited identification of many of the materials. The materials in 04DP-25, especially compressed maize cupules, showed evidence of heat-related and mechanical deformation. Locus 04AH-21 represented the next highest number of organic remains. This locus is the most deeply buried level sampled of mottled and lensed contents of a bell-shaped pit (locus 04AH-11), possibly used originally for subterranean storage, located in the Operation 4 North area. The pit was filled with trash in primary deposition, including elaborately decorated ceramics interpreted as brewing and drinking cups dating to a.d. 450–650. Several other loci processed come from levels of the contents of this pit deposited above this locus. They also contain a significantly high number of botanical remains.
RESULTS
In total, a minimum of 23 and a maximum of 60 distinct taxa were recovered from 36 excavated loci at Puerto Escondido (Table 2). This minimum number accounts for the possibility that some taxa were identified through one type of residue analysis (e.g., Poaceae seeds recovered from the light fraction) but redundant in another type of residue analysis with specimens that were unidentifiable, non-diagnostic, or diagnostic to a broader or narrower taxonomic clade (e.g., Panicoideae phytoliths recovered from adhering sediment). The maximum number of distinct taxa accounts for the possibility that no overlap exists between differentially identified residues. The actual number of recovered taxa is likely somewhere in between these two figures.
Of the minimum 23 species, at least 15 were recoverable only from artifact residues as microremains (e.g., Manihot [manioc]), at least four were recoverable only as macrobotanical remains from light fraction flotation samples (e.g., Nicotiana [tobacco]), and 2–4 overlapped across several types of samples (e.g., Fabaceae and Phaseolus and Poaceae and Zea mays) (Table 3). We record the total counts of each taxon in the final column of Table 2 to reflect the combined number of instances of each taxon recovered. Sediments adhering to artifacts are likely more representative of surrounding matrices and activities, whereas sonicated materials are likely more representative of actual artifact use (Pearsall Reference Pearsall2008; Perry Reference Perry2004). We calculated final ubiquity values using only the adhering sediments of the artifact dry wash and the light fractions of flotation samples (Table 2). This way, each location was counted only once in “overlapping” loci from which both types of samples were taken. Integrating the botanical data in this way allows us a better overall view of taxon presence across the site, in spite of the fact that each recorded presence might represent a different anatomical part (phytolith, starch grain, fruit, or seed).
Table 3. Taxa ubiquities in sonicated residue, tabulated by artifact type. Only taxa recovered from sonicated material are tallied. Taxa recovered from adhering sediment are not included in these totals. Numbers do not indicate raw counts of taxa, but rather reflect the number of instances in which each taxon was recovered. An individual artifact type from an individual locus is counted as a single instance.
A wide set of ethnobotanical practices is represented by the recovered residues, as inferred from diverse ethnohistoric and ethnographic sources. There are plants commonly associated with food use, plants more commonly associated with fiber and thatching purposes, plants frequently associated with medicinal use, and plants more commonly associated with ritualized practice. There is also a great deal of wood represented, which is to be expected given its almost universal use as fuel and in construction. These represent fluid categories, however, as many plants are utilized in a number of ways (described in the section Discussion below). This typological fluidity has been marked historically and in contemporary times, and is likely to have been evidenced in the ancient past, as well.
Common food plants recovered at Puerto Escondido, aside from the ubiquitous maize (Zea mays) include amaranth grain (Amaranthus sp.), palm fruits (Arecaceae spp.), squash fruits (Cucurbitaceae and Cucurbita spp.), hackberry (Celtis sp.), peppers (Capsicum sp.), goosefoot grain (Chenopodium sp.), beans (Fabaceae and Phaseolus spp.), and, potentially, bromeliads (Bromeliaceae family) and sagebrush (Artemisia sp.). Previous residue analyses confirmed the use of cacao (Theobroma sp.) as well. There were several root and tuber crops represented in the archaeological assemblage, including arrowroot (Maranta sp.), manioc (Manihot sp.), and possibly léren (Calathea sp.), along with plant remains generally categorized in the arrowroot family (Marantaceae spp.). Although spurge family (Euphorbiaceae) species have wildly differing properties, ranging from the simply painful to toxic, there are several species recorded as having food uses (Atran Reference Atran1993; Rico-Gray 1991), including chaya (Cnidoscolus spp.) and the already mentioned manioc. Outside of its use as edible grain, goosefoot leaves (recovered here as seeds) have also been widely cited as a condiment and an edible green (Standley and Steyermark Reference Standley and Steyermark1946). Hirtella, less commonly consumed, has been recorded as having edible fruit (Nelson Sutherland Reference Nelson Sutherland1986).
Plants that were likely used at Puerto Escondido primarily for fiber and thatching include the various grass species (Poaceae family; Arundinoideae, Bambusoideae, Panicoideae, and Pooideae subfamilies), as well as palm fronds (Arecaceae spp.), sedges (Cyperaceae and Cyperus spp.), and potentially bromeliad (Bromeliaceae family) leaves. Unidentified wood charcoal, along with an enormous abundance of unidentified wood phytoliths, mark construction and fuel use. Wood from several identified taxa, including hackberry, palm, and the bean family, may also have been used as fuel and in construction.
Ritualized practice may be represented in the archaeological samples by bromeliads, including those of the Tillandsia species. Species in this genus were historically used by the Honduran Lenca in rituals, particularly the construction of perishable altars decked with Tillandsia plants (Chapman Reference Chapman1985; Meluzín Reference Meluzín1997). Ritual use of plants merges with potential medicinal uses for many plant taxa. A wide range of plants—some recovered at Puerto Escondido—are mentioned in the translation of the colonial Yucatec Maya Ritual of the Bacabs (Roys Reference Roys1965), where they are embedded in incantations intended to restore health from conditions described generically as “seizures” or convulsions. Plants recovered from Puerto Escondido that are noted in the Ritual of the Bacabs include tobacco (Nicotiana spp.), species in the spurge (Euphorbiaceae) family (such as Tragia), and grasses (including Arundo spp. and Phragmites spp.). These plants were invoked to fight fainting, hiccups, tarantula-eruption, and tarantula-seizure, while Tillandsia was recommended to relieve fainting spells.
The plants recovered from Puerto Escondido that potentially index medical practices could have been applied to a wide variety of conditions as well as the general maintenance of good health. Aside from the Ritual of the Bacabs (Roys Reference Roys1965), plants with known medicinal uses are documented in ethnobotanical studies across southeastern Mesoamerica (Anderson Reference Anderson2003a, Reference Anderson, Pompa, Allen, Fedick and Jiménez-Osornio2003b; Arellano Rodríguez et al. Reference Arellano Rodríguez, Guido, Garrido and Bojorquez2003; Atran Reference Atran1993, Atran et al. Reference Atran, Lois and Ucan Ek2004; Balick and Arvigo Reference Balick and Arvigo2015; Cook Reference Cook2016; de Becerra Reference de Becerra2002; Kufer et al. Reference Kufer, Forther, Poll and Heinrich2005; Lentz Reference Lentz1986; Shanti Morell-Hart, personal observation 2006; Nelson Sutherland Reference Nelson Sutherland1986; Rico-Gray et al. 1991; Standley and Steyermark Reference Standley and Steyermark1946; Ticktin and Dalle Reference Ticktin and Dalle2005). The selection below links potential medical treatments with plants identified at the family or genus level at Puerto Escondido.
Some recovered plants may have been used to treat general maladies and conditions. Goosefoot (Chenopodium) leaves have been placed below pillows to induce sleep (Standley and Steyermark Reference Standley and Steyermark1946) and ingested for a variety of purposes (Anderson Reference Anderson2003a, Reference Anderson, Pompa, Allen, Fedick and Jiménez-Osornio2003b; Atran et al. Reference Atran, Lois and Ucan Ek2004; Balick and Arvigo Reference Balick and Arvigo2015; Cook Reference Cook2016). The juice from Cassia grandis legumes (bean family) has been consumed fresh to strengthen the blood (Lentz Reference Lentz1993). Plants in the palm family (Arecaceae) have been noted as having a variety medicinal uses (Anderson Reference Anderson2003a; Balick and Arvigo Reference Balick and Arvigo2015; Lentz Reference Lentz1986, Reference Lentz1993; Rico-Gray et al. 1991). More specific uses of plants for recognizable diseases have also been documented in these same ethnographic and historic sources. Tobacco (represented by a recovered seed) and the Panicoid grass Imperata contracta (potentially represented by phytoliths) are recognized as useful in treatment of chills, fever, and asthma. Spurge family genera such as Tragia, Cnidoloscolus, Croton, and Euphorbia have all been documented historically and ethnographically to treat rheumatism, “bone aches,” and coughs, and several of these may be represented in the assemblage at Puerto Escondido. Grasses such as Arundo and Phragmites have been prescribed as treatments for “phthisis,” an archaic term for wasting diseases with symptoms like tuberculosis.
Historic and ethnographic sources provide an equally rich range of potential medicinal uses of plants recovered at Puerto Escondido for digestive and urinary system diseases. Tobacco, the Panicoid grass Imperata contracta, and Tillandsia are generally cited for use in “bowel complaints,” with Imperata also cited as medicine for biliousness and jaundice (Roys Reference Roys1965) and tobacco (Nicotiana spp.) for urinary conditions. Arundinoid grasses, including Arundo and Phragmites, have been used to treat kidney pains (Arellano Rodríguez et al. Reference Arellano Rodríguez, Guido, Garrido and Bojorquez2003; Atran Reference Atran1993; Nelson Sutherland Reference Nelson Sutherland1986; Rico-Gray et al. 1991), dysentery, retention of urine, and ulcers (Roys Reference Roys1965). Tragia has relieved abdominal pains caused by sorcery (Roys Reference Roys1965). White sagebrush (Artemisia ludoviciana) has been used in southeastern Mesoamerica as a digestive tranquilizer (noted in the Centro de Investigación Científica de Yucatán [CICY] Jardín Botánico in Mérida, Yucatán [Shanti Morell-Hart, personal observation 2006]). Some arrowroot family (Marantaceae) species have been used as foods for invalids (Atran Reference Atran1993). Representatives of these genera and families were all recovered from the Puerto Escondido botanical assemblage.
The treatment of injuries, infections, and general pain would also have been possible with plants recovered from Puerto Escondido. Some plants in the spurge family (Euphorbiaceae), such as Jatropha curcas and Acalypha arvensis, have measurable antimicrobial properties (Lentz et al. Reference Lentz, Clark, Hufford, Meurer-Grimes, Passreiter, Cordero, Ibrahimi and Okunade1998), and may have been used to treat or prevent infections. Goosefoot leaves have been used in poultices for infections (Standley and SteyermarkReference Standley and Steyermark1946). Arundinoid grasses, including Arundo spp. and Phragmites spp., have been used to treat wounds, inflammations, and general pain (Arellano Rodríguez et al. Reference Arellano Rodríguez, Guido, Garrido and Bojorquez2003; Atran Reference Atran1993; Nelson Sutherland Reference Nelson Sutherland1986; Rico-Gray et al. 1991). Tillandsia and the Panicoid grass Imperata contracta were used to relieve headache and neuralgia (Roys Reference Roys1965). Eye conditions and skin diseases were both treated with tobacco, while Imperata contracta was also used for skin diseases. Tragia was specifically identified historically as a pimple medication (Roys Reference Roys1965). Ethnographically, the sap of Jatropha curcas is noted as sometimes applied to cold sores of the mouth (Lentz Reference Lentz1993).
Another wide area of medical practice includes plants used to treat discomfort from bites, stings, and parasites. These practices include treating tick bites and colmoyote worms with tobacco and spider bites with the flowers of Phyllanthus niruri (in the spurge family). Ethnographically, goosefoot leaves have been used as vermifuge when mixed with garlic infusion (Atran Reference Atran1993). Leaves of Gliricidia sepium (in the bean family) are boiled in water and used to bathe the head for lice infestations (Lentz Reference Lentz1993). Tobacco applied to clothing has been used to ward off snakes.
The many unknown phytoliths recorded in this study correspond with species undocumented so far, and would expand our ideas of plant use at Puerto Escondido if ever identified. Even with the abbreviated list of potential uses associated with identified taxa, however, a wide range of ethnobotanical practice is potentially revealed. Such diversity of plants indicates a rich knowledge of the surrounding landscape, and skills associated with the collection, management, and cultivation of diverse plant species (detailed below). Our recognition of this understanding would have been substantially narrower if only one of the analytical methods had been used.
Plant Taxa Represented on Artifacts and in Sediments: An Overview of Differences
The analysis of sonicated artifact residues allows us to link specific plants directly to artifact use. It is likely that many tools and vessels were also used in the processing and serving of animal products, and these activities may have left diagnostic protein or lipid signatures. Even with the study of botanical residues alone, however, we see a diversity of uses for specific artifacts, and some activities that appear across artifact types. The study of lithic and ceramic artifact residues has provided some unexpected findings, and in some cases, documented the use of specific plants that are identifiable solely through microbotanical analysis.
Ceramic Artifact Residue Samples
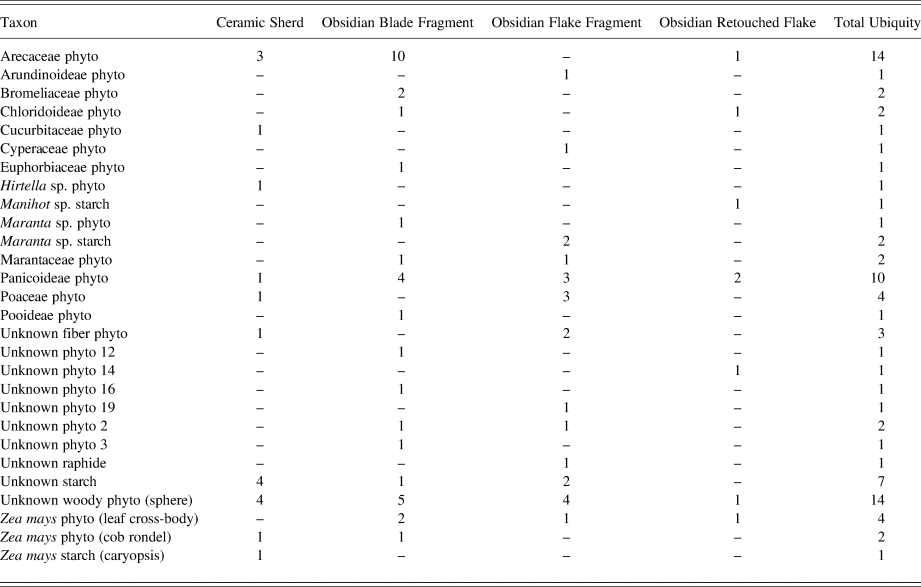
The four ceramic sherds included in this study are flat-bottomed, flaring-wall bowls with complex surface treatments in the types Boliche Black, Sukah Differentially Fired, Bonilla Yellow-Brown, and Fia Metallic Grey (Joyce and Henderson Reference Joyce and Henderson2007, Reference Joyce, Henderson, Blomster and Cheetham2017). All the sherds had been previously washed, and after rewashing and extraction through sonication, recovery of residues was limited, yet still quite informative (Figure 4). Maize was represented as five starch grains coming from a single bowl (the Boliche Black sherd). One maize rondel was also recovered, perhaps indicating the serving of cut maize cob sections, or the use of maize cobs as cleaning implements. Also of interest were Hirtella genus and squash family phytoliths, both from the Bonilla Yellow-Brown bowl. This may indicate the serving of sliced squash with the rind still attached. A few palm family phytoliths were scattered among the bowls, along with many damaged starch grains. Overall, the bowls contained few identifiable starch grains. This was expected, considering the common use of serving vessels to serve cooked foods that have undergone transformations in starch grain morphologies (Henry et al. Reference Henry, Hudson and Piperno2009), often leaving them fully gelatinized. Many woody species phytoliths were recovered (at least 50 percent of the assemblage in each case), indicating either fuel residues from the firing process, wood implements used to clean the sherds, or perhaps bits of ash incorporated into the slip matrices.

Figure 4. Select microremains identified in sonicated residues recovered from ceramic vessel interiors. (a) cf. Hirtella phytolith from Bonilla Yellow-Brown sherd. (b) Panicoid grass phytolith from Fia Metallic Grey sherd. (c) Unknown fiber from Boliche Black sherd. (d) Palm (Arecaceae) phytolith from Sukah Differentially Fired sherd. Images by Morell-Hart.
While the sample is limited, these results show that highly decorated bowls—of the same types and shapes that previously tested positive for cacao residues—were used to serve a wide range of plant-based foods, including maize, squash, and Hirtella. Differences in the specific residues recovered from individual vessels suggest that these bowls were used in a cuisine which featured different foods or recipes, reinforcing previous suggestions that these vessels were used to serve a complex cuisine appropriate for hosting a feast that included cacao in multiple forms (Joyce and Henderson Reference Joyce and Henderson2007, Reference Joyce, Henderson, Staller and Carrasco2009).
Lithic Artifact Residue Samples
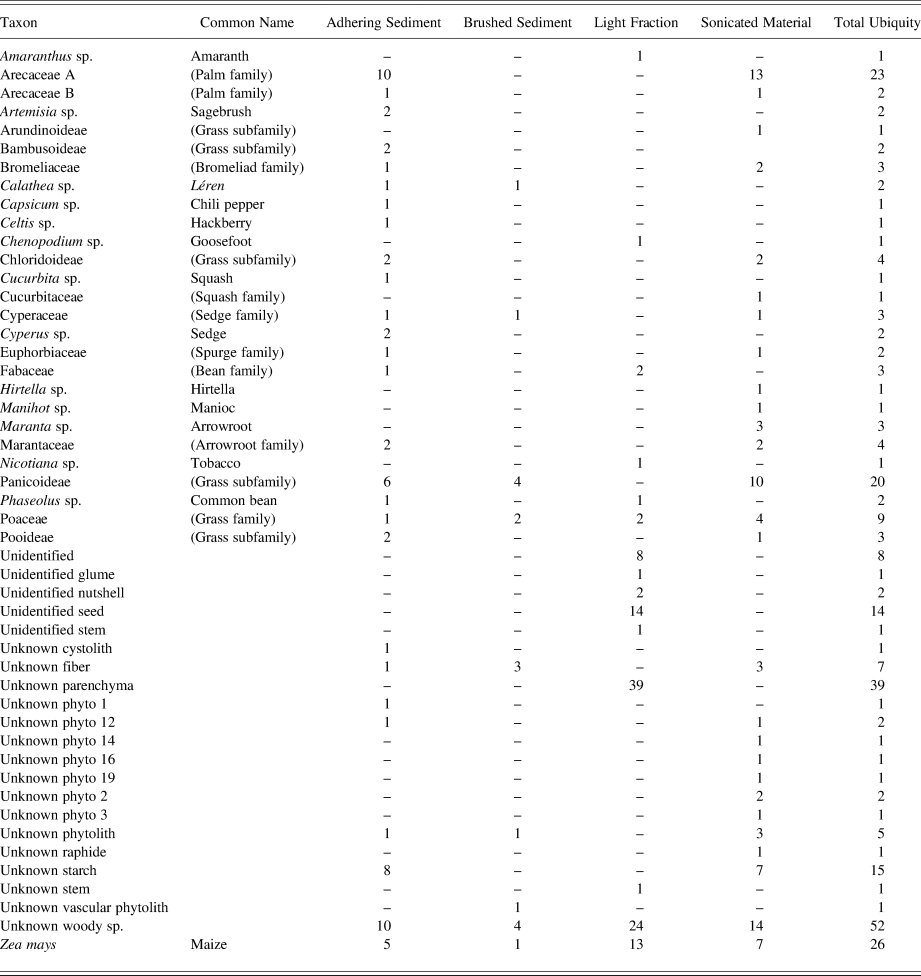
Obsidian blade fragments and flakes represent a variety of different kinds of deposits, different sectors of the site, and different time periods. Of the five obsidian blade fragments analyzed, two (04F-06-A and 04G-04-A) date between a.d. 200–450, while three others (04AH-17-A, 04DP-25-A, and 04DZ-02-A) date between a.d. 450–650. They were recovered from a variety of contexts—lined-pit fills, a high-density midden, and matrices between occupational surfaces—and evidenced a wide variety of uses (Figure 5). Palm family and grass family phytoliths were commonly recovered from the residues, as well as a few bromeliad phytoliths. Grasses present included Chloridoid and Pooid representatives (very rare in the assemblages) and Panicoid representatives (common in the assemblages). Only non-edible parts of maize—leaf and cob phytoliths—are represented, and no kernel starch grains were recovered. This may indicate the processing of maize leaves using obsidian blades and, as with the ceramic sherds, may represent the use of maize cobs to clean sharp implements. Several spurge family (Euphorbiaceae) and arrowroot family (Marantaceae) phytoliths were also recovered from the obsidian blade residues. Although these could be indicators of arrowroot and manioc tuber processing, the lack of starch grains (only a single starch grain was recovered from the sonicated material from all of these blades) would appear to indicate, as with the maize microfossils, fewer uses in processing starchy foods. In contrast, samples from sediments around the obsidian blades yielded residues from a wider variety of food plants, including arrowroot, hackberry, bean, and maize kernels, as well as many of the fibrous taxa previously mentioned (Figure 6). For this reason, there appear to be more general associations between these obsidian blades and fiber processing (of grasses, woods, and palm fronds) than with food processing (of roots, tubers, and other starchy tissues).
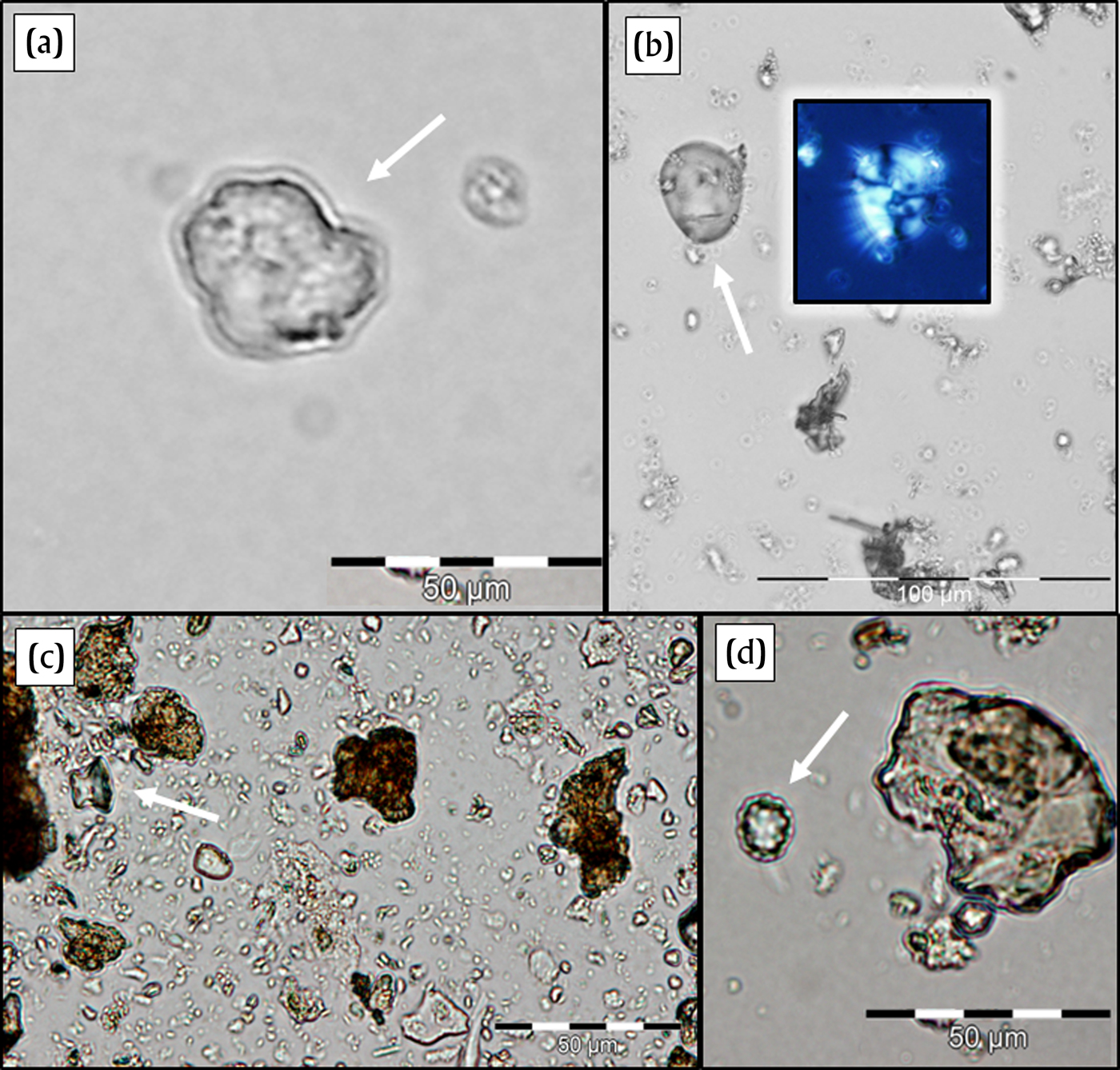
Figure 5. Select microremains recovered in sonicated residues from obsidian implements. (a) Sedge family (Cyperaceae) phytolith from 4DC-130. (b) Arrowroot (Maranta) starch grain under normal and polarized light, from 6C-67. (c) Maize (Zea mays) cob phytolith from 4F-6a. (d) Palm (Arecaceae) phytolith from 4AH-17a. Images by Morell-Hart.

Figure 6. Select microremains recovered from sediments adhering to obsidian implements. (a) Chili pepper (Capsicum) starch grain from 6C-67 under normal and polarized light. (b) Maize cob phytolith from 4AH-17a. (c) Cf. bean family (Fabaceae) starch grain from 4DK-136. (d) Maize (Zea mays) starch grain from 6C-67. Images by Morell-Hart.
Two retouched obsidian flakes were also sampled for residues: 04AL-07-A (dating between a.d. 200 and 400) and 04AW-19&20 (dating between 700 and 400 b.c.). The sonicated residue from 04AL-07-A included a single palm phytolith, two maize leaf phytoliths, and a starch grain that is likely manioc, along with woody species and Chloridoid and Panicoid grasses. Only a few phytoliths from woody species and palms were recovered from the adhering material, along with a single unidentifiable starch grain. These results would indicate a more wood- and fiber-oriented use for this flake (as with the obsidian blades), with perhaps a single use to process manioc starchy tissues. Similarly, the earlier retouched flake 04AW-19&20 was used for fibrous tissues (such as Panicoid grasses), but seems to have been used very infrequently or perhaps more frequently for animal tissues, given that the entirety of the sonicated material from this flake (analyzed fully in two slides) contained only three phytoliths. Wet-washed adhering sediment contained phytoliths of woody species only, whereas the dry-brushed surrounding sediments contained phytoliths from cf. lerén and maize along with many woody species and grasses.
Two other obsidian flake fragments (4DC-130 and 4DK-136) with no visible usewear were also analyzed. Both flake fragments come from contexts dating between 1150 and 1000 b.c., and ceramic sherds collected from these contexts yielded cacao residues (Henderson et al. Reference Henderson, Joyce, Hall, Hurst and McGovern2007; Joyce and Henderson Reference Joyce and Henderson2007). The sonicated residues included phytoliths from grasses (Poaceae: Arundinoid and Panicoid subfamilies), wood, sedges, and potentially a phytolith from maize leaf. As with the retouched flakes and blades already discussed, these implements seem to have been used primarily for fiber, leaf, and wood processing. In spite of the diversity of uses (at least four taxa cut/scraped by each flake) these activities did not leave much trace of usewear, indicating infrequent or very limited use-life for these two artifacts.
One obsidian flake fragment (06C-67) with visible usewear was also analyzed from the Operation 6 area, in deposits dating between 1400 and 1100 b.c. This artifact yielded a rich array of plant remains from the sonicated residues. Identified specimens included arrowroot starch grains, as well as phytoliths from the arrowroot family. There were also a number of other residues including grass family phytoliths, unknown fibers and phytoliths, wood phytoliths, and damaged starch grains. Chile pepper and maize starch grains were recovered from the wet-washed adhering sediments but not from the sonicated residues extracted from the obsidian flake itself. Only grass and wood phytoliths were noted in the exterior dry-brushed sediment attached to the flake. For these reasons, it appears that this flake fragment was either discarded among refuse that included chile and maize remnants, which preserved well in the adhering material but less well in the more exterior dry-brushed sediments, or that this flake fragment was discarded after first uses for arrowroot and fibrous materials and secondary uses for maize and chile. It is also possible that, during the course of cleaning this artifact in antiquity, chile and maize starches were transferred through dishwater or cleaning implements before eventual discard of the flake. In any event, this single early lithic and its adhering sediment provide a rich and complex picture of plant use, and of the way plants were processed, in an early village residential setting.
Comparison with Bulk Flotation Sediment Samples
Given the intensity of analysis required in working with artifacts, bulk flotation of sediment samples continues to provide the most widely feasible method for recovering plant remains. Yet the results presented above clearly show that not all plant remains will be recovered through flotation samples and, without artifact analyses, we run the risk of presuming uses for stone tools based on implement morphology only (cautioned against by Perry Reference Perry2004, Reference Perry2005). Similarly, as we demonstrate, flotation samples provide a rich view of plants that are either unrecoverable from artifact residues or unidentifiable to the same degree of taxonomic specificity.
The bulk flotation sediment samples analyzed in this project date primarily to the period between a.d. 200 and 650 (29 samples, a combined N = 1,245 items), with only a few dating between 900 and 400 b.c. (3 samples, a combined N = 8 items). The light fraction samples come from a variety of contexts, including lined-pit fills, burial matrices, matrices between occupational surfaces, hearths, high-density middens, an exterior surface, burned deposits, and ashy deposits. The early samples contained only unidentifiable plant tissues (an unidentifiable seed and amorphous fragments of parenchymous tissue) and two small wood fragments. In contrast, the diversity of plants represented in the later samples—as expected given the much larger sample volume—is much greater.
Two samples were recovered from matrices between occupational surfaces, yielding unknown wood and parenchymous tissues, as well as a cf. Phaseolus (common bean) cotyledon and a maize cupule. These clearly represent secondary or tertiary deposits of rubbish used as construction fill. Three burial matrices yielded a maize cupule, a bean family seed, and a tobacco seed, as well as many unknown wood and parenchymous tissues. None of the materials recovered from the matrix around burials are presumed to have been placed deliberately. Rather, it is assumed that they are in secondary or tertiary contexts, and incorporated into burial fill (detailed in Morell-Hart Reference Morell-Hart2011).
The majority of the flotation samples analyzed, however, came from contexts with potentially interpretable activities. One exterior surface deposit yielded only unknown parenchymous tissues and wood fragments, likely secondary or tertiary deposits from hearth rake-outs or ash sweeping. An ashy deposit yielded only an unidentifiable fragment of parenchymous tissue. A single burned deposit sample had a similar makeup of unidentifiable parenchymous tissues, along with wood and a maize cupule. Each of these three samples likely represents the in situ remnants of burned fuel, as well as potential detritus from root and tuber processing.
Almost everything recovered from the five hearth contexts was unidentifiable (damaged seeds) or unknown (small fragments of parenchymous tissues and wood). This is likely due to the high degree of fragmentation and vitrification from proximity to the hearth. A maize cupule and a bean family seed recovered were recovered, however, both from the same context (04G-06). This indicates some measure of cooking activity, as expected, near the hearth.
A single high-density midden context (04DP-25) contained enormous quantities of highly fragmented and eroded seeds, nutshell/dry fruits, parenchymous tissues, and wood. Disappointingly, little was identifiable. This sample included 274 maize cupules, however, accounting for 22 percent of the total macrobotanical assemblage across all samples and contexts. The well-preserved cupules in this case may indicate the dumping of hot ash on a midden containing maize cobs, or the light charring of cobs kept near a fire for cleaning or other purposes.
Lined-pit fills presented the greatest diversity of species. As with the midden context, many maize cupules were represented, though not at the density seen in the midden. Also, as with the midden and other contexts, many fragments of parenchymous tissue, wood, and eroded seeds were recovered. Although unidentified, these represent at least 10 taxa, distinguishable through mutually exclusive characteristics (variable general sizes and broad morphologies). Identified seeds included only amaranth (Amaranthus sp.) and goosefoot (Chenopodium sp.). Such clay-lined pits may represent storage or processing locations (similar to those described in Sheets Reference Sheets1998), later filled in with sweepings or other secondary or tertiary fills. The botanical materials recovered would be consistent with such late-stage use.
While the flotation samples provided a good basis to document the presence of dietary staples such as maize and beans, broad culinary plant use would appear very different without the analysis of artifacts. We would lack the confirmation of root crops, chile peppers, and squashes provided by phytoliths and starches, not to mention the absence of indicators of cacao food and drink provided by chemical analyses. Medicinal or ritual use of plants would have been suggested by the recovery of a tobacco seed, but the lack of bromeliad residues would not have allowed potential associations with historical use of Tillandsia by Lenca people, as we are able to do with direct recovery of bromeliad phytoliths from artifacts. Wood and fiber fragments were retrieved using both flotation and artifact sonication, but the specific intensity of obsidian tool use in fiber processing, and the identification of specific fiber materials, required artifact residue analysis. The processing of grasses made visible through the analysis of obsidian artifacts encouraged us to examine the assemblage of plants that might otherwise have been taken as environmental background. This analysis led to the realization of the rich medicinal uses potentially represented by various species.
DISCUSSION
No single method of recovery would have detected all the plants identified at Puerto Escondido, in contrast with the rich results derived from the suite of methods combined in our analysis. Recovering macrobotanical remains through flotation, phytoliths, and starches from artifact residues and distinctive chemical signatures from artifact matrices allows for the construction of a botanical inventory that exceeds any documented or hypothesized in the archaeological literature from Central America, though sites in the nearby Maya area have revealed rich botanical assemblages using both macrobotanical and microbotanical data (Cagnato Reference Cagnato2018; Cagnato and Ponce Reference Cagnato and Ponce2017; Dedrick Reference Dedrick2013; Devio Reference Devio2016; Dussol et al. Reference Dussol, Elliott, Michelet and Nondédéo2017; Farahani et al. Reference Farahani, Chiou, Cuthrell, Harkey, Morell-Hart, Hastorf, Sheets, Sayre and Bruno2017a, Reference Farahani, Chiou, Harkey, Hastorf, Lentz and Sheets2017b; Lentz Reference Lentz2001; Lentz and Hockaday Reference Lentz and Hockaday2009; Lentz et al. Reference Lentz, Woods, Hood, Murph and Robin2012, Reference Lentz, Magee, Weaver, Jones, Tankersley, Hood, Islebe, Ramos Hernandez, Dunning, Lentz, Dunning and Scarborough2015; Morgan Reference Morgan2010; Santini Reference Santini2016; Thompson et al. Reference Thompson, Hood, Cavallaro, Lentz, Lentz, Dunning and Scarborough2015).
Having a rich plant inventory is only the first step in shifting interpretation from pure subsistence reconstruction toward broader and more socially foregrounded ethnobotanical approaches (Morehart and Morell-Hart Reference Morehart and Morell-Hart2015). By examining artifacts to discover what plants they might have processed or contained, the analyses employed here open windows into botanical practices. The importance of processing fiber and wood merits particular attention, as most archaeological reports spend little time on such activities. Plants with medicinal applications also need to be kept in mind when interpreting presence and location of botanical residues. As illustrated above, plants were likely used in a variety of endeavors, and even plants commonly categorized as foodstuffs may have also formed part of medical and spiritual practice.
The wide range of ethnobotanical activities represented by plant residues indexes a deep knowledge of the surrounding landscape, as well as a broad skill set associated the procurement, processing, serving, and disposal of diverse plant species. Some plants and practices fall more frequently within the sphere of house lot and home, while other plants and practices fall more frequently within the sphere of field and forest.
Palm leaves, bromeliads, hackberry fruits, wood from dicotyledonous trees, and some herbaceous medicinal species may have come from wild or managed forests beyond the edge of the fields (Dussol et al. Reference Dussol, Elliott, Michelet and Nondédéo2017; Lentz Reference Lentz and White1999; Lentz and Hockaday Reference Lentz and Hockaday2009; Lentz et al. Reference Lentz, Beaudry-Corbett, Reyna de Aguilar and Kaplan1996; Rico-Gray et al. 1991; Thompson et al. Reference Thompson, Hood, Cavallaro, Lentz, Lentz, Dunning and Scarborough2015). Useful wild plants growing along riverbanks and in wetlands would have included sedges and some grass species (Dunning et al. Reference Dunning, Lentz, Scarborough, Lentz, Dunning and Scarborough2015; Fedick Reference Fedick, Mathews and Guderjan2017), although some sedges may have come from forest assemblages or fallowed milpas. Fields under cultivation likely contained food crops such as manioc, lerén, arrowroot, squash, beans, maize, and chile peppers, and perhaps goosefoot and amaranth (Atran Reference Atran1993; Ford and Nigh Reference Ford and Nigh2009, Reference Ford, Nigh, Morris, Jones, Awe, Thompson and Badillo2010; Lentz et al. Reference Lentz, Beaudry-Corbett, Reyna de Aguilar and Kaplan1996; Sheets et al. Reference Sheets, Dixon, Guerra and Blanford2011, Reference Sheets, Lentz, Piperno, Jones, Dixon, Maloof and Hood2012). During fallowing periods, fields likely contained herbaceous plants such as grass family species and medicinal herbs such as wild sagebrush, alongside fast-growing and nitrogen-fixing wild bean family species (Anderson Reference Anderson, Fedick and Taube1995; Lentz et al. Reference Lentz, Magee, Weaver, Jones, Tankersley, Hood, Islebe, Ramos Hernandez, Dunning, Lentz, Dunning and Scarborough2015). Even adventitious “weeds” pulled from areas under cultivation may have served occasionally as medicine, condiment, or expedient tool (Anderson Reference Anderson, Fedick and Taube1995). The archaeological deposits testify to robust practical knowledge and management of plants in forests, along riverbanks, and in cultivated and fallow fields. This knowledge included not only where plants grew but also how they could be harvested, processed, and used.
Closer to home, palms, fruit trees, shade trees, and medicinal herbs may have been planted in the house lot along with domesticated plants such as manioc and chile peppers (Atran et al. Reference Atran, Lois and Ucan Ek2004; De Clerck and Negreros-Castillo Reference De Clerck and Negreros-Castillo2000; Fedick et al. Reference Fedick, Delgadillo, Sedov, Rebolledo and Mayorga2008; House and Ochoa Reference House, Ochoa and Lok1998; Lentz Reference Lentz, Ramírez and Griscom1997; Lentz et al. Reference Lentz, Beaudry-Corbett, Reyna de Aguilar and Kaplan1996; Montagnini Reference Montagnini, Mohan Kumar and Ramachandran Nair2006). Preliminary processing of plants likely took place outdoors, while finer elaboration of medicines and meals likely took place indoors. Inside structures and next to hearths, implements were used in ways specific to particular plants or expediently for a variety of tasks. Different vessel types were considered appropriate for serving different preparations of foods, with some overlap between them. Disposal of remains including food scraps and ash likely took place at a range of distances in relation to processing areas, from the convenience of the house lot (e.g., swept out the door) to the more distant field (e.g., basketed and removed). These deposited remains could have been used in landforming or as mulch and fertilizer (Fedick et al. Reference Fedick, Delgadillo, Sedov, Rebolledo and Mayorga2008). There is also evidence that plant remains deposited in midden areas were reused as fills in later features and constructions inside the house lot (Morell-Hart Reference Morell-Hart2011). Making use of nearby resources—and depositing debris close to the home—entailed an intimate knowledge of plant uses (e.g., adventitious medicinal herbs) and plant needs (e.g., the use of fertilizer for garden plants), as well as an attentiveness to appropriate disposal areas (e.g., less potent-smelling plant refuse permitted near the home).
The combined botanical deposits reflect a robust practical knowledge of the surrounding landscape, near and far, and reveal affordances provided by decades and even centuries of human-environmental interaction. Cultivation, management, and other ethnobotanical activities transformed the landscape through the promotion of certain plants—intentionally or unintentionally—and had long-term impacts on local botanical communities. Such impacts have been signaled in studies that upend the Pristine Myth (Denevan Reference Denevan1992) to address “escaped species” of homegardens and orchards (Folan et al. Reference Folan, Fletcher and Kintz1979) and visualize the landscape of southeastern Mesoamerica as a “managed mosaic” (Fedick Reference Fedick and Fedick1996) or a “constructed niche” (Lentz et al. Reference Lentz, Magee, Weaver, Jones, Tankersley, Hood, Islebe, Ramos Hernandez, Dunning, Lentz, Dunning and Scarborough2015). Although models of early village life in southeastern Mesoamerica are frequently based on maize cultivation, results for this area of Honduras indicate a much lower ubiquity of maize than anticipated by such models (Morell-Hart Reference Morell-Hart2011; Morell-Hart et al. Reference Morell-Hart, Joyce and Henderson2014).
Instead, our results accord with models of settlements that are not predicated entirely on maize cultivation (e.g., Sheets et al. Reference Sheets, Dixon, Guerra and Blanford2011, Reference Sheets, Lentz, Piperno, Jones, Dixon, Maloof and Hood2012) and indicate ethnobotanical activities with a smaller footprint on the surrounding landscape, given the use of wild and fallow-dwelling species. The higher-impact cultivation practices necessary for intensive production of maize are complemented, and perhaps even counterbalanced by, the lower-impact cultivation strategies necessary for root-crop production. The many non-domesticated species gathered or managed by Puerto Escondido residents implicate the maintenance of places on the landscape that were not used exclusively for cultivation or horticulture. These more limited impacts on local ecology, as evidenced by the wide array of plants consistently cultivated and collected over several hundred years, could also implicate a fairly high degree of resilience over time. Although further paleoethnobotanical (and in particular, palynological) research in the region would help to better assess these ideas, the array of plants and practices revealed in this study nonetheless provide insights into local ethnoecological relationships.
Our analysis suggests that, with a richer inventory of plant remains, researchers are positioned to more thoroughly characterize the plant worlds that enveloped settlements. Such worlds, actively negotiated, maintained, and cultivated by ancient communities, included kitchen gardens, fields planted with staple crops, groves of trees under management, and forested and transitional zones of uncultivated plants. Understanding the complicated dynamic between people and plants requires a full array of analyses to move beyond flora inventories and subsistence strategies toward a more holistic view of ethnobotanical practice.
RESUMEN
En los últimos años, investigadores en Centroamérica han utilizado recursos nuevos para amplificar y enriquecer la evidencia de las plantas antiguas y sus usos. Aquí presentamos un estudio de Puerto Escondido, ubicado en el valle Ulúa de Honduras de la Costa Caribe. Demostramos la eficacia del uso de múltiples métodos para la interpretación de prácticas etnobotánicas en el pasado. Utilizando muestras macrobotánicas obtenidas por flotación de sedimentos, análisis de residuos microbotánicos de artefactos de obsidiana y cerámica, y análisis de residuos químicos de vasijas, proveemos datos nuevos sobre el uso de plantas en el período Formativo de Mesoamérica. Contribuimos al entendimiento de la complejidad del uso de plantas, la importancia relativa del maíz, y las ventajas de utilizar metodologías múltiples para detectar materiales botánicos en sitios arqueológicos.
Nuestros datos sugieren algo más complicado que la dependencia en el cultivo de maíz y otras plantas domesticadas, demostrado por la muy baja frecuencia del maíz, los frijoles y las calabazas. Esta evidencia, añadida a una muestra amplia de raíces comestibles, productos de árboles, y el uso de plantas para otros fines además de los culinarios, indica un conocimiento profundo del medioambiente cerca del solar y a una distancia más larga. Aquí, nos dirigimos a prácticas botánicas “adentro de casa”—tales como actividades culinarias, prácticas medicinales, elaboración de fibra y actividades ritualizadas, y prácticas “cerca de casa,” como las prácticas agrícolas y hortícolas, forestales y otros compromisos con ecologías locales y distantes.
Nos presenta un modelo más amplio, basado en métodos múltiples, para entender mejor el uso de plantas durante los primeros siglos del desarrollo de las sociedades complejas en Honduras, y probablemente, en otras partes de la región. En particular, señalamos la importancia imprevista de productos de palmas y plantas silvestres, y el amplio rango de tubérculos procesados con las herramientas examinadas. Nos permita una oportunidad para desarrollar un entendimiento de la etnoecología en Puerto Escondido, un concepto definido aquí como la relación dinámica entre las habilitaciones presente en un dado medioambiente y los impactos de las actividades humanas en ese medioambiente.
ACKNOWLEDGMENTS
The authors would like to thank the many excavators and apprentices who aided in the recovery and processing of materials from Puerto Escondido. Funding for the research was provided by the Stahl Fund of the Archaeological Research Facility, a Research Assistantship in the Humanities, and a Research Enabling Grant of the Committee on Research of the Academic Senate of the University of California, Berkeley.









