The effect of light adaptation on the period of photocurrent saturation induced by a bright stimulating flash was examined in rod photoreceptors of the larval-stage tiger salamander (Ambystoma tigrinum). Using suction electrodes, photocurrent responses to brief flashes were recorded from single, isolated rods in the presence and absence of steady background illumination. Background light decreased the saturation period (T) measured at fixed flash intensity (fixed If) and in this respect light-adapted the saturating response. Effects of the background on responses to weak (i.e. subsaturating) and bright flashes were compared with changes in a parameter, 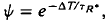 where ΔT is the decrease in saturation period, and where TR* is the slope of the line that relates T and ln If in a given state of adaptation. Dark- and light-adapted responses to flash intensities
where ΔT is the decrease in saturation period, and where TR* is the slope of the line that relates T and ln If in a given state of adaptation. Dark- and light-adapted responses to flash intensities  and
and 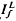 , respectively, exhibited similar absolute peak photocurrent and falling-phase kinetics when
, respectively, exhibited similar absolute peak photocurrent and falling-phase kinetics when 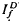 and
and 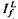 satisfied the relation,
satisfied the relation, 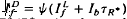 , where Ib is the background intensity. It is argued that ψ approximates the relative PDE*/R* gain of transduction, i.e. the relative peak level of activated cGMP phosphodiesterase (PDE*) produced by a given, small amount of photoactivated visual pigment (R*). Interpreted on this view, the results imply that light adaptation derives largely from a decrease in PDE*/R* gain, rather than from the stimulation of guanylate cyclase activity. The data are consistent with the possibility that modulation of the lifetime of PDE* underlies the background dependence of ψ.
, where Ib is the background intensity. It is argued that ψ approximates the relative PDE*/R* gain of transduction, i.e. the relative peak level of activated cGMP phosphodiesterase (PDE*) produced by a given, small amount of photoactivated visual pigment (R*). Interpreted on this view, the results imply that light adaptation derives largely from a decrease in PDE*/R* gain, rather than from the stimulation of guanylate cyclase activity. The data are consistent with the possibility that modulation of the lifetime of PDE* underlies the background dependence of ψ.
