Introduction
The onset of male reproductive system development in rats occurs during intrauterine life with the beginning of testosterone production, extending up to 60 d after birth, and divided into the following phases: neonatal (postnatal day – PND 1–7), infant (PND) 8–21), juvenile (PND 22–25), and peri-pubertal (PND 36–55 or 60). Reference Clegg1,Reference Ojeda, Andrews, Advis and White2 There is little production of testosterone during the neonatal phase, but during the infantile and juvenile phases, primary androgens are produced. In addition, between days 28 and 56, Leydig cells differentiate into adult cells with an increase in testosterone production. Puberty begins with the pulsatile release of gonadotropins into the central nervous system for further stimulation of steroid hormone production by the gonads, Reference Ojeda and Skinner3 culminating in physical, behavioral, and hormonal alterations which lead to final sexual maturation and reproductive capacity. Reference Golub, Collman and Foster4 The four phases of development of the male reproductive system in rats are controlled by genetic and hormonal mechanisms, which are the periods most vulnerable to disturbance in both control mechanisms, leading to changes that last into or manifest during adulthood. Reference Norgil Damgaard, Maria Main, Toppari and Skakkebæk5 The developmental period is sensitive to environmental factors such as drugs and environmental pollutants. During the prenatal and early postnatal life, cells differentiate, and specific tissue is formed, thus, plasticity is particularly prevalent. It is known that environmental influences during the developmental period affect etiology and susceptibility to diseases, for example, neurodevelopmental and neurodegenerative diseases/dysfunctions, changes in the timing of puberty and infertility. Reference Heindel, Balbus and Birnbaum6,Reference Barouki, Gluckman, Grandjean, Hanson and Heindel7 Knowing that prenatal and early childhood events influence the probability of developing diseases, allows for interventions or prevention to be targeted at the most effective time.
Exogenous substances that interfere with the synthesis, secretion, transport, binding, or elimination of endogenous hormones responsible for homeostasis are called endocrine disruptors (ED), for example, synthetic substances (pesticides, phthalates, polychlorinated biphenyls, bisphenol A, and pharmaceutical substances, among others) and natural substances, as well as natural estrogens and phytoestrogens. 8 Topiramate (TOP) is a psychotropic drug approved by the Food and Drug Administration for the treatment of epilepsy in children older than 2 years of age, Reference Ben-Menachem, Sander, Stefan, Schwalen and Schäuble9,Reference Chung and Eiland10 as well as being used since 2014 for migraine prophylaxis in adolescents aged 12 to 18 years. 11,Reference Sakulchit, Meckler and Goldman12 In humans, a 39-year-old patient diagnosed with partial epilepsy and treated with TOP 100 mg/d for 2 months experienced loss of libido and sperm every 2–3 d, which ceased approximately one week after the end of treatment. Reference Wu, Hao and Zhou13 Nonetheless, another case report suggested that erectile dysfunction observed in a man induced by TOP may be related to a blockage of AMPA receptors leading to the inhibition of the glutamatergic pathway in predisposed patients, since sex hormones levels were normal and libido was subjectively increased, Reference Calabrò, Bramanti, Italiano and Ferlazzo14 showing contradictory data in the literature. A study with adult Sprague Dawley rats treated with TOP (100 mg/kg) for 60 d, showed a reduction in reproductive organ weight, sperm motility, spermatocyte and spermatid numbers, and Leydig cells, Reference Sameer, Hamed and Zuheir15 thus suggesting that TOP may act as an ED.
Therefore, given the evidence that TOP can affect the reproductive system, its wide use during periods of reproductive system development, and the lack of studies that elucidate the possible effects of TOP used during childhood and adolescence, this work is justified by the need to evaluate the immediate and late effects of treatment with TOP during childhood and adolescence on the rat reproductive system. Moreover, the major goal of the present study was to reproduce the exposure to TOP during those periods and due to the lack of experimental studies on rats, as a result this study was based on clinical studies on children and adolescent human beings.
Material and methods
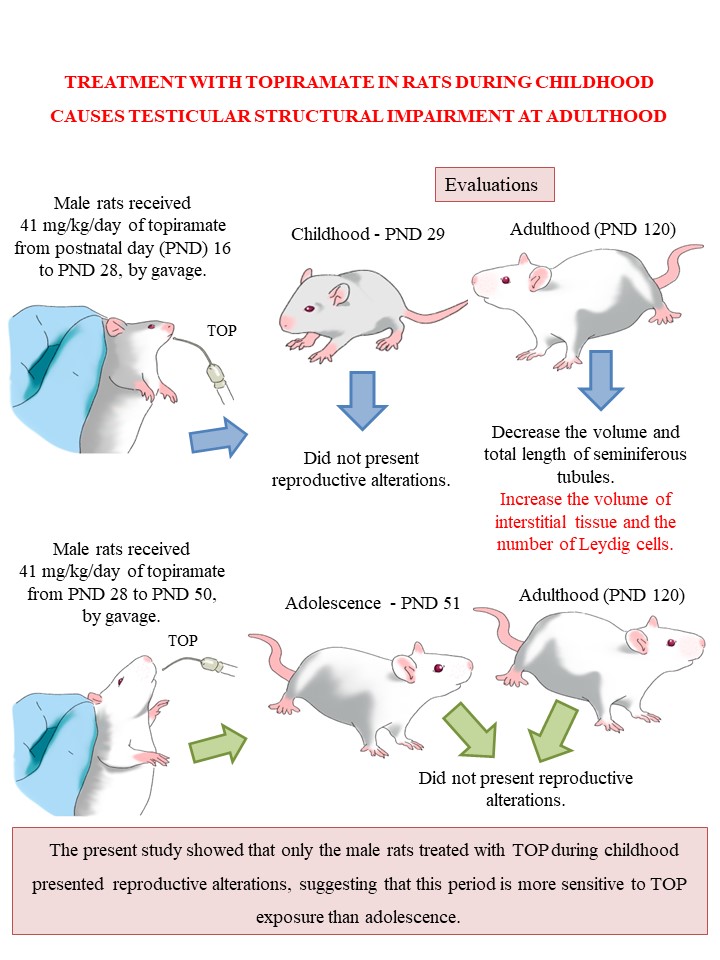
All protocols were approved by the Animal Ethics Committee of UEL (CEUA / UEL 100/2018). The experimental design is depicted in Fig. 1.

Fig. 1. Diagram of the experimental design. PND: postnatal day.
Experiment 1
Male and female Wistar rats (75 d) from the colony of the State University of Londrina (UEL), were used as the parental generation. The animals were transferred to the vivarium of the Department of Physiological Sciences where they were kept in a controlled environment with a temperature of 23 ± 2 °C, 12 h light/dark cycle (lights on at 6:00 a.m.) and had free access to regular lab chow (Nuvilab, Brazil) and tap water. The animals were given a 15-d period of adaptation to the environment before the beginning of mating. Two females were mated with one male and gestational day (GD) 0 was determined as the day of sperm detection in vaginal smears. The pregnant rats were individually housed in cages. Pregnancy monitoring was performed, and the day of birth of the pups was denominated postnatal day 0 (PND). The pups were counted, the sex identified, and the litter weighed. At PND 4, litters were reduced to 10 animals (5 males and 5 females whenever possible, and the females were used in another study), so that breastfeeding was homogeneous between the litters. At PND 16, male offspring were distributed into 2 experimental groups that were weaned at PND 21, and received the following treatments:
–CTRc group: Water daily, by gavage (0.3 mL/100g body weight), from PND 16 to PND 28 (n = 20)
–TOPc group: 41 mg/kg/d of TOP (Topamax®, Janssen-Cilag, Brazil) suspended in water, daily, by gavage, from PND 16 to PND 28 (n = 20)
These groups, treated during childhood, were subdivided into four subgroups (n = 10 animals/ subgroup). Two subgroups (CTRc and TOPc) were evaluated 24 h after the end of treatment (PND 29), and the other two subgroups (CTRc and TOPc) were evaluated in adulthood (PND 120). Thus, the immediate and late repercussions of TOP exposure were evaluated.
Antiepileptic treatment with TOP is approved in children over 2 years of age. 16,Reference Cella, Knibbe, Danhof and Della Pasqua17 In rodents, PND 16 corresponds to 3 years of age in humans, Reference Marco, Adriani, Ruocco, Canese, Sadile and Laviola18 in this way, the males were treated from PND 16 to PND 28, to cover the period equivalent to that of childhood in rats, and in order to cover the critical developmental stages of the reproductive systems. In children and adolescents, the effective dose range is 3-9 mg/kg/d. Applying the BW3/4 scaling, 19 the equivalent dose in rats would be 41 mg/kg/d.
Experiment 2
The rats were obtained from the colony of the State University of Londrina at PND26 and transferred to the vivarium of the Department of Physiological Sciences where they were kept under the same conditions as Experiment 1. At PND28, adolescent rats were assigned to 2 experimental groups, which received the following treatments daily:
–CTRa group: Water daily, by gavage, from PND 28 to 50 (n = 20)
–TOPa group: 41 mg/kg/d of TOP (Topamax®, Janssen-Cilag, Brazil) suspended in water, daily, by gavage, from PND 28 to 50 (n = 20)
These groups, treated during adolescence, were subdivided into four subgroups (n = 10 animals/group) as early described and were and evaluated 24 h after the end of treatment (PND 51) or in adulthood (PND 120).
Anogenital distance
The anogenital distance (AGD, distance from the anus to the genital tubercle) was obtained using a vernier caliper and normalized through its division by the cube root of body weight, Reference Gallavan, Holson, Stump, Knapp and Reynolds20 obtained on PND 16 and 120 in the childhood group, and PND 28 and 120 in the adolescence group.
Collection of plasma and organs
On the specific days in each group (PND 29, 51, and 120), the animals were euthanized with sodium thiopental overdose (40 mg/kg, i.p.), a thoracotomy was performed for blood draw using cardiac puncture, from the left ventricle, into tubes containing heparin, between 8:00 am and 10:00 am. Immediately after, blood samples were centrifuged (10,584×g for 20 min at 4 °C) and the plasma was frozen until assayed. After blood collection, the right vas deferens was removed and used for the analysis of sperm motility and counting in the vas deferens. The left vas deferens was removed for further analysis of sperm morphology. In addition, the reproductive organs (testis, epididymis, seminal vesicles, and prostate) were removed, dissected, and their wet weights obtained using an analytical balance. The left testis and epididymis were frozen for sperm count, and the right testis and epididymis were stored for histological analysis.
Sperm evaluation
Sperm motility
Sperm were obtained from the right vas deferens and diluted in1 ml of GV HEPES (Ingamed, Brazil) pre-warmed at 34°C. A 10 µl aliquot was placed in a Makler chamber (Sefi-Medical, Haifa, Israel) and analyzed under a phase-contrast microscope (OSM-223287, Olympus) at 100× magnification. One hundred spermatozoa were evaluated per animal and classified as motile or immotile. Reference Perobelli, Martinez, Da Silva Franchi, Fernandez, De Camargo and De Grava Kempinas21,Reference Favareto, Fernandez, da Silva, Anselmo-Franci and Kempinas22
Sperm count in the vas deferens
After sperm motility analysis, the sperm count was performed in the vas deferens analysis (million/ml). The average number of sperms counted was obtained in 10 different squares, chosen randomly in four different fields in the Makler chamber (Sefi-Medical, Haifa, Israel). Reference Piffer, Garcia, Gerardin, Kempinas and Pereira23
Daily sperm production per testis, sperm number and transit time in the epididymis
The left testis was decapsulated and the caput/corpus and cauda segments from the epididymis were separated. Homogenization resistant testicular spermatids (step 19 of spermiogenesis) and sperm in the caput/corpus epididymis and cauda epididymis were assessed as described previously by Robb et al. Reference Robb, Amann and Killian24 with adaptations of Fernandes et al. Reference Fernandes, Arena, Fernandez, Mercadante, Barbisan and Kempinas25 Mature spermatids were counted in a Neubauer improved chamber (Optik Labor, 0.100 mm Tiefe Depth Profondeur). To calculate daily sperm production (DSP), the number of spermatids at step 19 was divided by 6.1, which is the number of days in one seminiferous cycle when these spermatids are present in the seminiferous epithelium. Sperm transit time through the epididymis was determined by dividing the number of sperm in each segment by the DSP.
Sperm morphology
The left vas deferens was carefully flushed, with the aid of a syringe and needle, with 1 ml of formol-saline (10%) to remove the sperm and this content was prepared on histological slides that were left to dry for 90 min. A total of 200 spermatozoa were analyzed per animal in a phase-contrast microscope (OSM-223287, Olympus, 400× magnification). Morphological abnormalities were classified into two general categories: head abnormalities and tail abnormalities. Reference Fernandes, Arena, Fernandez, Mercadante, Barbisan and Kempinas25
Histomorphometric parameters of the testis
The right testis of male rats was dissected, weighed, and fixed by immersion in Methacarn solution for 4h before being stocked in ethanol at 70°, until processing. The testis was cut into tissue fragments which were routinely processed for embedding in Paraplast® (McCormick Scientific). Blocks were sectioned at 5 µm intervals for tissues collected from each animal and stained with hematoxylin–eosin. For testis morphometry, the average diameter of fifteen round sex cord cross sections/seminiferous tubules per animal was obtained with a photo camera (Moticam 5, Motic) coupled to an ocular microscope with 100× final magnification, using the software ImagesPlus 2.0 (Motic). The volume densities of various testicular tissue components were determined by light microscopy using a 100-intersection grid placed in the ocular of the light microscope. Ten fields, chosen randomly (1000 points), were scored for each animal at 400× magnification. The volume of each component of the testis was determined as the product of the volume density and testis volume. Because the testis density is nearly 1.0 (∼1.03–4), for subsequent morphometric calculations the testis weight was considered equal to testis volume. To obtain a more precise measure of testis volume the testis capsule (∼6.5%) was excluded from the testis weight. Reference Attal, Courot, Richetin and Pisselet26,Reference Dorst and Sajonski27 To count the Leydig cell nuclei, 10 random fields of interstitial tissue in each testis section per rat were counted, under a light microscope at 400× magnification. Reference Guerra, Sanabria and Leite28 The Leydig cell morphology was identified as described by Teerds and Huhtaniemi (2015). Reference Teerds and Huhtaniemi29
Plasma testosterone quantification
Plasma testosterone was measured by chemiluminescence microparticle immunoassay (ARCHITECTTM, 2nd Generation Testosterone, Abbott Laboratories, IL, USA) according to the manufacturer’s instructions. The intra-assay coefficient of variation and the minimum sensitivity of the assay were 4,6% and 0.15 nmol/L, respectively.
Statistical analysis
Initially, an exploratory analysis was conducted to evaluate normal distribution (Shapiro–Wilk test) and homogeneity of variance (Levene’s test) of each variable. Variables that presented normal distribution and homogeneity of variance were analyzed by the Student’s t-test. Conversely, for other variables, the Mann–Whitney test was performed. Analysis of covariance (ANCOVA) was used to detect the effect of treatment on organ weight using the final body weight as the covariate. The data on sperm count were analyzed by the Fisher’s test and are presented as a percentage. All statistical procedures were performed using SPSS version 19 for Windows (SPSS Inc.).
Results
Results from experiment 1 – Childhood
Bodyweight, weight of reproductive organs, anogenital distance, and testosterone level were not influenced by treatment with TOP during childhood at PND 29 (Supplementary Table S1).
The seminiferous tubule of the testis of rats treated with TOP during childhood are shown in Fig. 2.

Fig. 2. Photomicrographs of seminiferous tubule (ST) sections of rats from the postnatal day (PND) 29 CTRc (A), TOPc (B); at PND 120 CTRc (C) and TOPc (D); at PND 51 CTRa (E) and TOPa (F); and at PND 120 CTRa (G) and TOPa (H). Histological slides were stained with hematoxylin–eosin. Experimental groups received 41 mg/kg of topiramate daily, by gavage, from PND 16 to 28 (TOPc group) or from PND 28 to 50 (TOPa group). Control groups (CTRc group or CTRa group) received tap water daily. Half of the animals were evaluated 24 h after the end of treatment (PND 29 and PND 51, respectively) and the remainder were evaluated in adulthood (PND 120). Scale = 100 μm. It was analyzed in a phase-contrast microscope (100× magnification). ST: seminiferous tubule; I: Interstice.
The results of the histomorphometry of the testis are presented in Table 1. There were no statistical differences between the groups in any parameters.
Table 1. Histomorphometry of the testis from rats at PND 29 treated during childhood

The data were analyzed by the Student’s t-test and are means ± SEM (p > 0.05). Numbers in brackets represent the number of animals/group. CTRc: Received water by gavage during childhood. TOPc: Received TOP 41 mg/kg/d during childhood.
Bodyweight, weight of reproductive organs, anogenital distance, and testosterone level in adulthood were not influenced by TOP treatment during childhood, as indicated in (Table 2).
Table 2. Bodyweight, weight of reproductive organs, anogenital distance, and testosterone level from rats in PND 120 treated during childhood

The data were analyzed by the Student’s t-test and are means ± SEM (p > 0.05). Numbers in brackets represent the number of animals/group. CTRc: Received water by gavage during childhood. TOPc: Received TOP 41 mg/kg/d during childhood.
The results of the histomorphometry of the testis from the rats in experiment 1 at PND 120 are shown in Table 3 and Fig. 2 shows the seminiferous tubule of the testis. Treatment with TOP during childhood increased the volume of interstitial tissue and the number of Leydig cells and decreased the volume of seminiferous tubules and the total length of seminiferous tubules.
Table 3. Histomorphometry of testis of rats at PND 120 treated during childhood

The data were analyzed by the Student’s t-test and are means ± SEM (*p < 0.05 vs CTRc). Numbers in brackets represent the number of animals/group. CTRc: Received water by gavage during childhood. TOPc: Received TOP 41 mg/kg/d during childhood.
The data of the sperm parameters of rats submitted to treatment with TOP during childhood were not altered (Supplementary Table S2 and S3).
Results from experiment 2 – Adolescence
Bodyweight, weight of reproductive organs, anogenital distance, and testosterone level were not influenced by treatment with TOP during adolescence (Supplementary Table S4).
The seminiferous tubule of the testis of rats treated with TOP during childhood are shown in Fig. 2 and the results of histomorphometry of the testis are presented in Table 4. There were no statistical differences in TOPa animals when compared to the control group at PND 51.
Table 4. Histomorphometry of the testis from animals at PND 51 treated during adolescence

The data were analyzed by the Student’s t-test and are means ± SEM (p > 0.05). Numbers in brackets represent the number of animals/group. CTRa: Received water by gavage during adolescence. TOPa: Received TOP 41 mg/kg/d, by gavage, during adolescence.
Body weight, weight of reproductive organs, anogenital distance, and testosterone level at adulthood were not influenced by treatment with TOP during adolescence (Supplementary Table S5).
At PND 120, histomorphometry of the testis was not influenced by the treatment with TOP during adolescence as shown in Table 5 and Fig. 2.
Table 5. Histomorphometry of testis from animals at PND 120 treated during adolescence

The data were analyzed by the Student’s t-test and are means ± SEM (p > 0.05). Numbers in brackets represent the number of animals/group. CTRa: Received water by gavage during adolescence. TOPa: Received TOP 41 mg/kg/d, by gavage, during adolescence.
The data on sperm parameters of rats submitted to TOP treatment during adolescence were not altered (Supplementary Table S6 and S7).
Discussion
Traditional antiepileptic drugs, for example valproate and gabapentin, are related to sexual dysfunction in humans, including decreased libido and erectile dysfunction. Reference El Makawy, Ibrahim, Mabrouk, Ahmed and Fawzy Ramadan30 These drugs may induce sexual dysfunction by reducing bioactive testosterone, accelerating sexual hormone metabolism, and stimulating hormone binding globulin production. Reference Wu, Hao and Zhou13 However, studies are emerging on new antiepileptic drugs, such as TOP, and relating them to adverse effects on the reproductive system in mice, such as decreased plasma testosterone, decreased spermatogenesis, and decreased mRNA related to germ cell genes. Reference El Makawy, Ibrahim, Mabrouk, Ahmed and Fawzy Ramadan30 However, there is a lack of studies investigating the effects of antiepileptic drugs on the reproductive system during initial development periods, since these drugs are indicated for children over two years old and adolescents.
The results obtained in the present study revealed a decrease in the volume and the total length of seminiferous tubules and an increase in the volume of interstitial tissue as well as, Leydig cell numbers at PND 120 in rats treated with TOP during childhood, while the other parameters were not affected by TOP exposure.
AGD, an effective biomarker to detect fetal androgen insufficiency, is applied in strategies for detecting endocrine disrupting effects. Reference Schwartz, Christiansen, Vinggaard, Axelstad, Hass and Svingen31 Drugs can cause abrupt hormonal changes may interfere with AGD. Reference Schwartz, Christiansen, Vinggaard, Axelstad, Hass and Svingen31 In the present study, TOP treatment during childhood and adolescence did not compromise the AGD in male rats, suggesting no abrupt alteration in the sexual hormone levels during development, also confirmed by the testosterone level.
It has been demonstrated that the growth pattern of an animal is responsive to the presence of a toxic substance and this may occur in a dose-dependent way. Reference Johnson32 Treatment with TOP (41 mg/kg/d) did not alter the final body weight and reproductive organ weights in either the childhood or adolescence groups. In the literature, a decrease in the weight of the reproductive organs (testis, epididymis, seminal vesicle, ventral prostate, and vas deferens) is demonstrated in adult rats treated with TOP (100 mg/kg) for 60 d. Reference Sameer, Hamed and Zuheir15 The divergence in results might be related to the differences in doses and treatment periods between the studies.
In addition, the treatment with TOP did not alter sperm parameters (count, production, motility, and morphology) of adult rats treated during childhood and adolescence. Spermatogenesis begins concomitantly with puberty under the stimulation of LH and FSH, which act on gonads, stimulating the production of testosterone, which initiates and maintains spermatogenesis. Reference Zirkin, Santulli, Awoniyi and Ewing33 In our study, although the testosterone levels were increased (not statically significant) in the TOP group, exposure during childhood ensured the complete development of spermatogenesis, as this depends entirely on testosterone activity. Reference Pakarainen, Zhang, Mäkelä, Poutanen and Huhtaniemi34 Though the sperm parameters were not affected by the TOP treatment, it is important to highlight that these results do not assure fertility, moreover their gametes could present epigenetic alterations which could compromise the next generation. Further studies are necessary to investigate the reproductive ability of these males and their offspring.
There is evidence that TOP leads to a decrease in the thickness of the germinative epithelium of adult mice at doses of 200–400 mg /kg/d, Reference El Makawy, Ibrahim, Mabrouk, Ahmed and Fawzy Ramadan30 highlighting the ability of this drug to impair testicular tissue. In the present study, treatment with TOP during childhood (PND 16 to 28) decreased the volume and total length of seminiferous tubules in animals at PND 120. The decrease in the volume filled by seminiferous tubules is correlated with a decrease in the total length of seminiferous tubules. The longitudinal growth of seminiferous tubules is directly influenced by the plasma concentration of FSH. Reference Leidl, Bentley and Gass35 In adult Sprague Dawley rats treated with TOP (100 mg/kg) there is a reduction in plasma concentration of FSH. Reference Sameer, Hamed and Zuheir15 Although, the plasma testosterone level was not statistically different from the control, it is important to highlight that testosterone levels in the TOP group exposed during childhood were almost double compared to the control, which could be extremely high inside the testes. Such high testicular testosterone would interfere with the hypothalamus–pituitary–testicular axis. Thus, our hypothesis is that TOP compromised the testosterone levels, which may have led to oscillation in the plasma concentration of FSH, reflected in a decrease in the total length of the seminiferous tubules in adulthood. Moreover, the increase in testosterone levels is related to the rise in Leydig cell numbers, causing an increase in the volume of interstitial tissue. The absence of immediate changes in testicular tissue at PND 29 suggests that TOP may have led to changes in the programming of FSH producing cells, since the treatment period (from PND 16 to 28) covered a critical development window during which the plasticity of the cells is higher and they are, therefore, more vulnerable to environmental influences. Reference Godfrey, Lillycrop, Burdge, Gluckman and Hanson36
TOP treatment during adolescence (PND 28 to 50) did not cause changes in the histological parameters of the testis. The postnatal development of this organ begins at birth. At PND 15, Sertoli cells are mature and stop dividing, and at the same age, the spermatogonia divide into spermatids through meiosis. From PND 32 to 55, the peri-pubertal period occurs, which activates spermiogenesis, culminating in sperm development. Reference Picut, Remick, de Rijk EP c., Simons, Stump and Parker37 Thus, the significant period of testicular growth occurs from PND 21–32. Reference Picut, Remick, de Rijk EP c., Simons, Stump and Parker37 Thus, the group of adolescents received treatment for only 5 d of that period, while treatment in childhood covered a period of 13 d, which may explain the absence of impairment in the group of adolescents.
In summary, the present study revealed that treatment with TOP affects the histomorphometry of the testis of male rats in adulthood, specifically shown by an increase in the Leydig cell number. These effects were especially seen in the group exposed only during the childhood period. It is suggested that this period is more sensitive to TOP. It is important to highlight, that this is the first study to examine the immediate and long-term effects of treatment using this drug, in doses based on what would be suitable for children and adolescent human beings, during sensitive periods of their development. In this regard, it is important to emphasize the importance of studying the safety of treatment using TOP on reproductive functions in children as the results found do not assure male fertility. Moreover, epigenetic alterations could be present in the gametes which can compromise the next generation. Therefore, further investigation is needed to better understand the mechanisms by which this drug promotes testicular changes and their consequences.
Supplementary materials
For supplementary material for this article, please visit https://doi.org/10.1017/S2040174422000587
Acknowledgments
This work was supported by CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior), Brazil (Master fellowship to Lorena I. Borges) and to Bruno H. Forcato for designing the Graphical abstract.
Financial Support
Part of this study was financed by Fundação Araucária de Amparo à Pesquisa: grant 215/2022-PBA.
Conflicts of Interest
The authors declare no conflicts of interest.
Ethical Standards
The authors assert that all procedures contributing to this work comply with the ethical standards of the relevant national guides on the care and use of laboratory animals (Brazilian College of Animal Experimentation) and has been approved by the institutional committee (State University of Londrina Ethics Committee for Animal Research).