



Introduction
Caffeine, a psychoactive substance, is a methylxanthine found in many beverages, foods, and in some medications. As a result, caffeine exposure occurs throughout life, even during critical developmental periods, such as gestation, childhood, and adolescence. Reference Frary, Johnson and Wang1–Reference Ruiz and Scherr3 Daily caffeine consumption is reported by approximately 70% of pregnant women, and by 75% of children 5 year and older in the United States. Reference Frary, Johnson and Wang1,Reference Ahluwalia and Herrick4 For the last few decades, caffeine consumption by teenagers has become an important issue, particularly due to energy drink consumption. Reference Ruiz and Scherr3,Reference Glatter, Myers and Chiamvimonvat5,Reference Pennington, Johnson, Delaney and Blankenship6 Among juveniles from 9- to 17-year-old, daily caffeine consumption more than doubled since 1980, being highly prevalent among teenagers nowadays. Reference Frary, Johnson and Wang1,Reference Ahluwalia and Herrick4,Reference Temple7 Despite its widespread consumption, the safety of caffeine ingestion during critical periods of brain development is not clear.
In rodents, long-term effects of caffeine exposure during early development have already been described. Prenatal caffeine exposure causes learning and memory impairments in adult rat offspring. Reference Li, Zhang and Shi8 Caffeine exposure during gestation and lactation increases anxiety-like behavior Reference Laureano-Melo, da Silveira and de Azevedo Cruz Seara9 and causes hippocampal GABAergic neuronal loss Reference Silva, Métin and Fazeli10 at adulthood. In adolescent rodents, caffeine impairs decision-making and increases emotional reactivity and anxiety-like behavior. Reference Anderson and Hughes11–Reference da Silva Gonçalves, Leal-Rocha, Manhães, Filgueiras, Abreu-Villaça and Ribeiro-Carvalho14 In this sense, mood disorders and sleep disturbances have been associated with caffeine consumption by human teenagers. Reference Seifert, Schaechter, Hershorin and Lipshultz15–Reference Marmorstein17
Interestingly, both human and animal studies also provide evidence that adolescents that consume caffeine are more likely to use other drugs, such as tobacco. Reference Temple7,Reference Leal and Jackson18–Reference Terry-McElrath, O’Malley and Johnston20 Even though the association between caffeine and tobacco smoking is still under investigation, it is established that nicotine, the main psychoactive component of tobacco, reduces caffeine’s half-life, which explains, in part, the fact that smokers drink more coffee than nonsmokers. Reference Swanson, Lee and Hopp21 Moreover, smokers who have moderate/high caffeine consumption show greater availability of α4β2 nicotinic acetylcholine receptors (nAChRs), one of the primary nicotine’s site of action, Reference Brody, Hubert and Mamoun22 and some animal studies reported increased psychostimulant and reward nicotine effects induced by caffeine, Reference Liu and Jernigan23–Reference Justinova, Ferré and Barnes26 supporting the idea that caffeine increases tobacco consumption due to increased nicotine reward effects. On the other hand, caffeine-induced increase in emotional reactivity could trigger tobacco smoking. Reference Kayir and Uzbay27–Reference Tobias, Templeton, Collings, Tobias, Templeton and Collings29 Despite the worldwide reduction in tobacco consumption over the past decades, the rapid increase in use of electronic nicotine delivery systems (ENDS), particularly during adolescence, Reference Cullen, Gentzke and Sawdey30–Reference Miech, Leventhal, Johnston, O’Malley, Patrick and Barrington-Trimis32 has renewed the relevance of studies that investigate nicotine and caffeine interactions.
Most studies that investigate behavioral and neuropharmacological interactions between these drugs focus on adulthood. However, caffeine is consumed throughout life. Here, we investigated the effects of lifetime (gestational and postnatal) exposure to caffeine on nicotine reward and these drugs interactions on anxiety-like behavior. Since nicotine activation of nAChRs in the ventral tegmental area neurons mediates dopamine release into the striatum and frontal cortex, induces hyperactivity, and modulates anxiety levels, Reference Govind and Vezina33–Reference McGranahan, Patzlaff, Grady, Heinemann and Booker36 we evaluated the effects of lifetime caffeine on nicotine-induced hyperlocomotion as well as on the modulation of anxiety levels in the open field test during adolescence. Considering that lifetime caffeine use may evoke neuroplastic adaptations that play a role in mechanisms of nicotine reward, which may not be identified after acute exposure, we also investigated whether caffeine increases the susceptibility of adolescent mice to the reinforcing effects of repeated exposure to nicotine via the conditioned place preference (CPP) test. Furthermore, since adenosine receptors, caffeine’s primary site of action, regulate the frontocorticostriatal catecholaminergic system, Reference Pandolfo, Machado, Kofalvi, Takahashia and Cunhab37 which is involved in drug addiction, Reference Ceceli, Bradberry and Goldstein38 we evaluated dopamine content, DOPAC, dopamine turnover and norepinephrine levels in the frontal cerebral cortex. The serotonergic system is implicated in anxiety Reference Gottschalk and Domschke39–Reference Asan, Steinke and Lesch41 and the hippocampus is a key structure in the circuitry involved in this behavior. Reference Mah, Szabuniewicz and Fiocco42 Interestingly, caffeine affects the serotonergic system of the hippocampus through its action on adenosine receptors. Reference Okada, Kawata and Murakami43 Therefore, here, we evaluated the hippocampal expression of the serotonin 1A receptor (5HT1A), a subtype abundantly expressed in this region, which is involved in the modulation of the anxiety response. Reference Gross, Zhuang and Stark44,Reference Gordon and Hen45
Method
All experimental procedures were carried out with the approval of the Ethical Committee for Animal Research of the Universidade do Estado do Rio de Janeiro (protocol number: CEUA/038/2014) and in accordance with Brazilian Law #11.794/2008. Swiss mice were bred and maintained on a 12 h light/dark cycle (lights on: 2:00 a.m., lights off: 2:00 p.m.) at a controlled temperature (∼21°C). All animals had free access to food and filtered water, and behavioral tests were carried out in a sound-attenuated room near our animal facility.
Treatment
Caffeine exposure (anhydrous, 1, 3, 7-trimethylxanthine - Proquímios – BR; diluted in filtered water) extended from the first day progenitors mating until the last day of adolescent offspring behavioral testing (Fig. 1). Two doses of caffeine were used based on previous rodent studies, corresponding to low and moderate caffeine intake. Reference Ardais, Borges, Rocha, Sallaberry, Cunha and Porciúncula12,Reference Fredholm, Bättig, Holmén, Nehlig and Zvartau46 Thus, three experimental groups were formed: CAF0.3, which had free access to a caffeine solution of 0.3 g/L (n = 14 litters); CAF0.1, which had free access to a caffeine solution of 0.1 g/L (n = 11 litters); and CTRL, which had free access to filtered water (n = 14 litters). These were the sole fluid sources for each group throughout the experiment. Detailed data on the estimated caffeine dose are provided as Supplementary Material (Tables S4 and S8). Only litters with 8–12 pups at birth (PN1) were used. For dams, body mass, liquid, and food ingestion were measured during lactation (PN2–PN21). Regarding litters, body mass was measured from PN2 until PN30. As for fluid and food intake, data were obtained from PN21 to PN30.

Fig. 1. Experimental timeline. From the first day of mating (M1), dams received a caffeine solution at a concentration of 0.1 or 0.3 g/L, or filtered water as the sole source of liquid until weaning. From that day onwards, pups received the same fluid offered to their mothers. Therefore, exposure extended from the first embryonic day to the last day of behavioral testing. At postnatal day (PN) 28, the offspring were randomly assigned either to the Open Field test (OF), performed at PN30, or to the Conditioned Place Preference test (CPP), which extended from PN28 to PN39. Brain tissues were harvested after completion of the behavioral testing and used for High Performance Liquid Chromatography (HPLC) and Western Blotting (WB) analyses.
Behavioral tests
At PN28, half of the mice of each sex from a given litter was randomly assigned to one of two behavioral tests: open field (OF) or CPP. For both tests, animals from each group were assigned to either the saline (SAL) or to the nicotine (NIC: (−) - nicotine hydrogen tartrate salt, 0.5 mg/Kg, calculated as free base in a volume of 0.01 mL/g; Sigma, St Louis, MO, USA) subgroups. Before each test, animals were acclimated to the testing room for 10 min.
Open field test
This test is widely used to evaluate locomotor activity. Reference Seibenhener and Wooten47–Reference Prut and Belzung49 Here, we aimed to study interactions between lifetime caffeine and acute nicotine, particularly in the context that the latter is known to induce locomotor hyperactivity, Reference Brielmaier, McDonald and Smith50 which reflects nicotine effects on the mesocorticolimbic pathway. Reference Pistillo, Clementi, Zoli and Gotti51–Reference Ferrari, Le Novère, Picciotto, Changeux and Zoli53 Since the OF is also used to assess anxiety-like behavior Reference McGranahan, Patzlaff, Grady, Heinemann and Booker36,Reference Morissette, Tull, Gulliver, Kamholz and Zimering54,Reference Picciotto, Brunzell and Caldarone55 and since increased anxiety is known to contribute to drug use, this behavioral trait was also assessed with the OF.
At PN30, immediately before the OF, animals received an intraperitoneal (i.p.) injection of NIC or SAL. Next, animals were placed in the center of the OF arena (45.5 cm length, 45.5 cm width, 34 cm height, black acrylic box) to freely explore it for 10 min. All sessions were performed during the lights-on cycle, between 11:00 a.m. and 1:00 p.m., and the arena was cleaned with paper towels soaked in 70% ethanol and dried before each test. Sessions were video recorded for subsequent analysis. The arena was divided into 16 squares of equal dimensions (12 peripheral squares and 4 center squares). The number of crossed squares was used as a locomotor activity indicator. As for the anxiety-like behavior, two variables were used: the time spent in the center of the arena and the time spent in the center of the arena relative to the number of crossed squares (time in center/locomotor activity ratio). Reference Prut and Belzung49
Conditioned place preference test
The CPP test is extensively used to investigate rewarding properties of drugs of abuse, such as nicotine, in rodents. In this test, the animals are conditioned to associate the effects of the drug with the environment in which it is administered. This association is considered a measure of drug-seeking behavior. Reference Liu, Le Foll, Liu, Wang and Lu56–Reference Tzschentke58 Here, CPP was used to assess whether lifetime caffeine affects nicotine-CPP.
The CPP apparatus (Insight, São Paulo, Brazil) consists of a box with three adjoining chambers: the “start box” is located between the other chambers, is painted gray, and is intended to be a neutral compartment. From there, two doors can be used by the animals to access the laterally located chambers, which have different floors and walls. One of the chambers is painted with alternating white and black vertical stripes on the walls and has parallel steel bars as the floor. The other chamber has walls painted with alternating white and black horizontal stripes and a checkered steel grid floor. The test protocol Reference Dutra-Tavares, Silva and Nunes-Freitas59,Reference Nunes-Freitas, Manhães and Dutra-Tavares60 consisted of four phases: habituation, pretest session, conditioning sessions, and test session. In the first 2 days (PN28–29), the animals received a once-daily i.p. injection of saline. The pretest session, which took place on PN30, was video recorded and the time spent in each lateral chamber was used to determine the preferred and nonpreferred sides. For this, each animal received an i.p. injection of saline and was immediately placed in the start box to freely explore the apparatus for 15 min. A preference of 75% or more for one of the lateral chambers was used as exclusion criterion (animals were excluded from subsequent testing). In the next 8 days (PN31–PN38), the conditioning sessions were performed. Mice from each group (CTRL, CAF0.1 or CAF0.3) were randomly assigned into either SAL or NIC subgroups. A biased design was used, Reference Tzschentke58 accordingly, nicotine was paired with the nonpreferred chamber identified in the pretest session. Mice were submitted to two sessions of 15 min per day (the first between 8:00 a.m. and 12:00 p.m. and the second between 2:00 p.m. and 6:00 p.m.). NIC-paired animals received nicotine (0.5 mg/kg) once a day: in one session, animals received nicotine paired with the nonpreferred side and, in the other session, they received saline paired with the preferred side. The session sequence was alternated throughout the conditioning period. The SAL groups received saline in both sessions/chambers. At PN39, the animals’ preference for the NIC-paired side was evaluated in a test session. Each animal was placed in the start box to freely explore the apparatus for 15 min. No injections were administered. This test was video recorded, and the time spent in each lateral chamber was quantified. CPP was accepted when the time spent in the non-preferred chamber after the conditioning sessions was higher than before these sessions.
Neurochemical analysis
Brain dissection
Immediately after each test, mice were killed by cervical dislocation and brains were dissected. Reference Abreu-Villaça, Correa-Santos and Dutra-Tavares61 Briefly, blunt cuts were made through the cerebellar peduncles and the cerebellum (including flocculi) was lifted from the underlying tissue. The midbrain + brain stem was separated from the remaining tissue by a cut made rostral to the thalamus. The cerebral cortex was separated from the midbrain + brain stem by a cut made caudal to the thalamus and the frontal 1/3 was collected. The hippocampus was removed from both left and right cerebral cortices. The brain regions were weighted, frozen in liquid nitrogen, and stored at −80°C until assayed. The frontal cerebral cortex and hippocampus were used for neurochemical analysis. These regions were chosen due to their relevance in the modulation of rewarding properties of drugs and expression of anxiety-like behaviors. Reference Gross, Zhuang and Stark44,Reference Seth, Cheeta, Tucci and File62,Reference Pistillo, Fasoli and Moretti63
High performance liquid chromatography (HPLC)
The frontal cerebral cortex was used to assess dopamine content, DOPAC, dopamine turnover, and norepinephrine levels. Tissues removed after the OF were used to evaluate the effects of chronic caffeine exposure, while the ones dissected after the CPP were also used to evaluate the effects of subchronic exposure to nicotine and the interactions between these drugs. The HPLC’s methodology was optimized for our analytical conditions. Reference Yoshitake, Kehr, Todoroki, Nohta and Yamaguchi64–Reference Ohkura, Kai and Nohta66 A Shimadzu Prominence LC-20AT HPLC with fluorescence detector (RF- 20A), adjusted to excitation λ of 345 nm and emission λ of 480 nm, was used. The mobile phase was a gradient system of acetonitrile/acetate buffer (20 mM; pH 4.5) containing EDTA (0.5 mM). The flow rate was 0.1 mL/min and the run lasted 70 min. A reverse-phase column (150 mm × 2.6 mm i.d.; packed with C 18 silica, 3 μm) was used for separation. Briefly, each frontal cortex was thawed and homogenized with 200 μL of HClO4 (0.1 M) plus sodium ascorbate (20 μM) (Precellys, BERTIN Technologies, Montigny-le-Bretonneux, France) and centrifuged at 5200×g for 30 min (4°C). The supernatant was filtrated in a 0.22 μm PVDF filter (Millipore) and diluted in 4 volumes of milli-Q water. DA and DOPAC derivatization reactions were accomplished using 10 μL standard solution or tissue sample + 20 μL of glycine (2.5 M) + 10 μL NaIO4 (5 mM) + 20 μL of acetonitrile + 50 μL of derivatization solution (0.1 M 1,2-Diphenyl-ethylenediamine / 0.1 M glycine / 62 mM sodium methoxide / 3.75 mM potassium hexacyanoferrate III). After 3 min at ambient temperature, a 20-μL portion of the final reaction mixture was injected onto the chromatograph.
Western Blotting
After the OF, the hippocampus was used to assess the effects of chronic caffeine exposure on 5-HT1A receptor levels. Tissues were homogenized in cell lysis buffer (pH 7.4), containing a protease inhibitor cocktail (Complete ULTRA Tablets, Mini, EDTA-free, EASYpack, Roche Applied Science, Mannheim, Germany) and sodium orthovanadate (Sigma Aldrich, Mannheim, Germany) and then supernadant was collected after centrifugation (13,000 rpm, 4°C, 25 min). The total protein content of each homogenate was determined by the bicinchoninic acid (BCA) protein assay kit (QuantiPro, BCA Assay, Sigma Aldrich, St Louis, MO, USA). Homogenates were denatured in the sample buffer (2-mercaptoethanol and bromophenol blue (50 mM Tris–HCl, pH 6.8, 1% SDS, 5% 2-mercaptoethanol, 10% glycerol, 0.001% bromophenol blue) and heated at 90°C for 5 min. Samples were stored at −45°C until the SDS-PAGE assay. 5HT1A receptors in hippocampus samples were assessed with a 12% polyacrylamide gel and transferred to PVDF membranes (Hybond P ECL membrane; Amersham Pharmacia Biotech, NJ, USA). Blocking of nonspecific binding sites in membranes was made with TBS-T containing 5% nonfat dry milk for 1 h. After that, membranes were incubated overnight with specific primary antibody anti-5HT1A (1:1,000) (Abcam; cat#: ab85615). In the next day, membranes were washed and then incubated with secondary anti-rabbit antibody (1:10,000) (Sigma-Aldrich; cat#: B8895) for 1 h. This was followed by washing and incubation with streptavidin HRP-conjugate (1:10,000) (Sigma-Aldrich; cat#: RPN1231V) for 1 h. The protein bands were visualized by chemiluminescence by Amersham ECL Prime Western Blotting Detection Reagent (GE Healthcare Bio-Sciences Uppsala, Sweden). The area and density of the bands were quantified by Image J software (Wayne Rasband National Institute of Health, MA, USA). The results were normalized by β-actin content (Sigma-Aldrich; cat#: A2228).
Statistical analyses
All data were analyzed using the IBM SPSS Statistics, Version 21.0 (IBM Corp, Armonk, NY, USA). Data were compiled as means and standard errors. Significant differences were defined as p < 0.05 (two-tailed).
Mixed-model analyses of variance (mxANOVA) were carried out for body mass, fluid and food intake. Day was considered the within-subjects factor. Caffeine (CTRL, CAF0.1, CAF0.3) and Sex were used as between-subjects factors. To minimize the influence of litter effects, the average of values from mice of the same sex and litter was used as the datum in the analysis. Significant main effects and interactions were followed by lower order ANOVAs and by Fisher’s Protected Least Significant Difference (FPLSD) tests.
Regarding the OF and neurochemical analysis, univariate ANOVAs (uANOVAs) were conducted. The between-subjects factors were Caffeine (CTRL, CAF0.1, CAF0.3), Nicotine (NIC or SAL), and Sex. For the CPP, mxANOVA were performed. Session (pre-test and test sessions) was considered the within-subjects factor. For both uANOVAs and mxANOVAs, significant main effects and interactions were followed by lower order ANOVAs and by FPLSD tests. There were no significant interactions between Sex and the other factors; therefore, for statistical purposes, male and female data were pooled together and figures show consolidated data of males and females. Figures segmented by sex as well as figures describing results of secondary variables are shown as supplementary material (Figures S1−S8 for the OF and S9−S16 for the CPP).
Results
Body mass and ingestion assessments
Body mass (Day: F 4.8,167.2 = 5.2, p < 0.001) (Table S1), food (Day: F 3.8,110.6 = 21.3, p < 0.001) (Table S2), and fluid intake (Day: F 3.4,72.9 = 12.6, p < 0.001) (Table S3) of dams increased during lactation. Chronic exposure to caffeine did not affect these variables. Regarding the offspring, as expected, body mass increased throughout lactation and post-weaning periods (Day: F 1.5,44.6 = 673, p < 0.001) (Table S5). No significant effect of caffeine was observed. In addition, no significant differences were identified for food and fluid intake (Tables S6 and S7, respectively).
Open field test
Mice exposed to nicotine were hyperactive (Nicotine: F 1,114 = 14.1, p < 0.001; Fig. 2A). Lifetime caffeine did not affect locomotor activity. Mice exposed to the higher caffeine dose (0.3 g/L) spent less time in the center of the OF, suggesting increased anxiety-like behavior (Caffeine: F 2,58 = 6.0, p < 0.01). Interestingly, while nicotine exposure per se did not affect the time spent in the center, it mitigated caffeine-mediated increase in anxiety-like behavior (Fig. 2B). There were no significant effect or interactions for the time in center/locomotor activity ratio (supplementary material, Figures S7 and S8). Additional OF metrics are presented as supplementary material (Figures S1–S6). Despite previous evidence that hippocampus 5HT1A receptors modulate anxiety levels, neither caffeine nor nicotine exposure affected the expression of this receptor in the frontal cerebral cortex (Fig 2C). Similarly, no significant differences between groups were detected for dopamine, DOPAC, dopamine turnover, and norepinephrine levels in the frontal cerebral cortex of animals tested in the OF (Table 1).
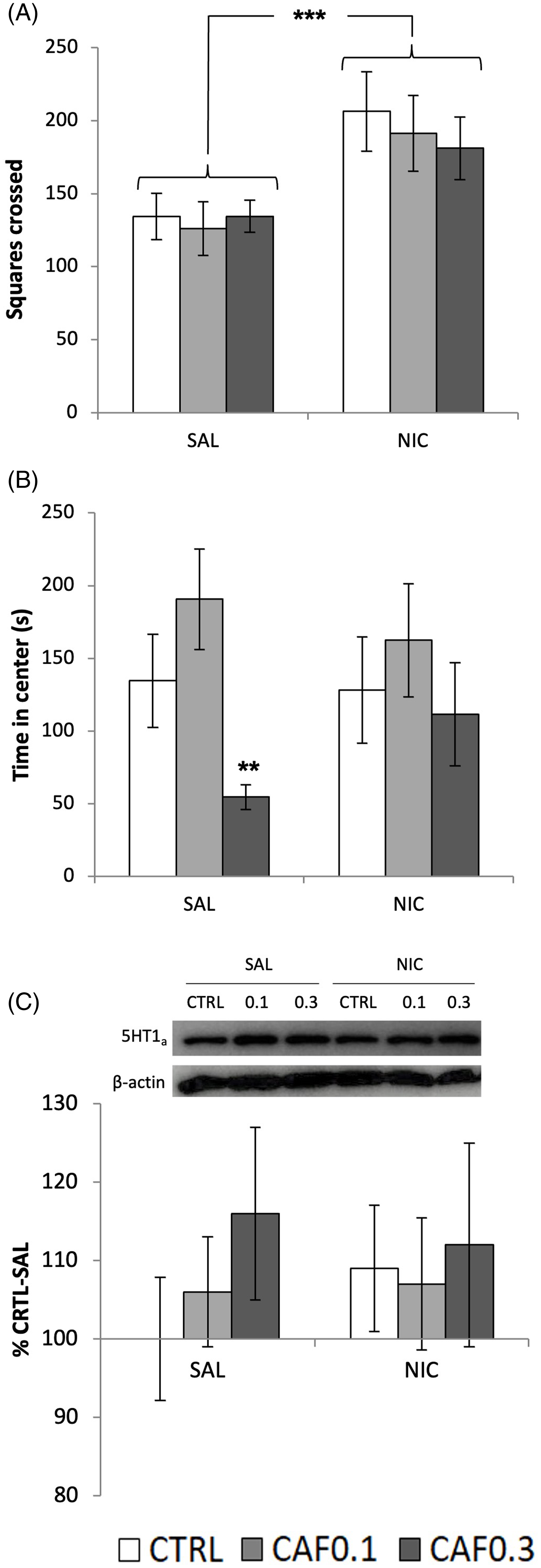
Fig. 2. (A) Locomotor activity in the open field test (OF) expressed as the total number of squares crossed. (B) Anxiety-like behavior expressed as the time spent in the center of the open field arena. (C) 5HT1A expression in the hippocampus of mice tested in the OF. Mice were chronically exposed to caffeine (CAF) in the drinking water at one of two concentrations (0.1 or 0.3 g/L), or to filtered water (CTRL) during development and, immediately before the behavioral test, received an i.p. injection of nicotine (NIC: 0.5 mg/Kg) or saline (SAL). For (A) and (B): CTRL-SAL, ♀ = 12 + ♂ = 10; CTRL-NIC, ♀ = 9 + ♂ = 12; CAF0.1-SAL, ♀ = 9 + ♂ = 11; CAF0.1-NIC, ♀ = 8 + ♂ = 11; CAF0.3-SAL, ♀ = 10 + ♂ = 12; CAF0.3-NIC, ♀ = 10 + ♂ = 12. For (C): CTRL-SAL, ♀ = 6 + ♂ = 6; CTRL-NIC, ♀ = 6 + ♂ = 6; CAF0.1-SAL, ♀ = 3 + ♂ = 6; CAF0.1-NIC, ♀ = 6 + ♂ = 5; CAF0.3-SAL, ♀ = 5 + ♂ = 4; CAF0.3-NIC, ♀ = 6 + ♂ = 3). Values are means ± S.E.M. ** p < 0.01; *** p < 0.001, FPLSD.
Table 1. Neurochemical analysis - frontal cortex
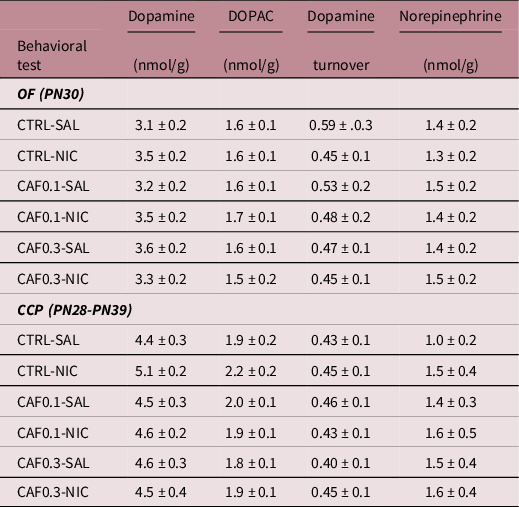
Dopamine, DOPAC, dopamine turnover and norepinephrine levels in the frontal cerebral cortex. Mice were chronically exposed to caffeine (CAF) in the drinking water at one of two concentrations (0.1 or 0.3 g/L), or to filtered water (CTRL) during development. Mice were exposed to nicotine (NIC: 0.5 mg/Kg) or saline (SAL) either before the open field test (OF) (CTRL-SAL, ♀ = 6 + ♂ =8; CTRL-NIC, ♀ = 5 + ♂ = 8; CAF0.1-SAL, ♀ = 5 + ♂ = 8; CAF0.1-NIC, ♀ = 9 + ♂ = 5; CAF0.3-SAL, ♀ = 6 + ♂ = 6; CAF0.3-NIC, ♀ = 7 + ♂ = 5) or during the conditioned place preference test (CPP) (CTRL-SAL, ♀ = 6 + ♂ = 6; CTRL-NIC, ♀ = 8 + ♂ = 6; CAF0.1-SAL, ♀ = 7 + ♂ = 5; CAF0.1-NIC, ♀ = 7 + ♂ = 7; CAF0.3-SAL, ♀ = 6 + ♂ = 6; CAF0.3-NIC, ♀ = 8 + ♂ = 6). Values are means ± S.E.M.
Conditioned place preference test
As expected, for saline-exposed mice, there was no difference in the time spent in the non-preferred side between the pretest and test sessions (Figure S11). Nicotine exposure during conditioning days increased the time spent in the nonpreferred chamber in the test session when compared to the pretest one (Session: F 1,54 = 9.6, p < 0.01), indicating induction of CPP (Fig. 3). Lifetime caffeine did not affect nicotine-induced conditioning (Fig. 3). Additional metrics for the CPP can be found as supplementary material (Figures S9–16). No significant differences between groups were detected for dopamine, DOPAC, dopamine turnover, and norepinephrine levels between groups in the frontal cerebral cortex (Table 1).
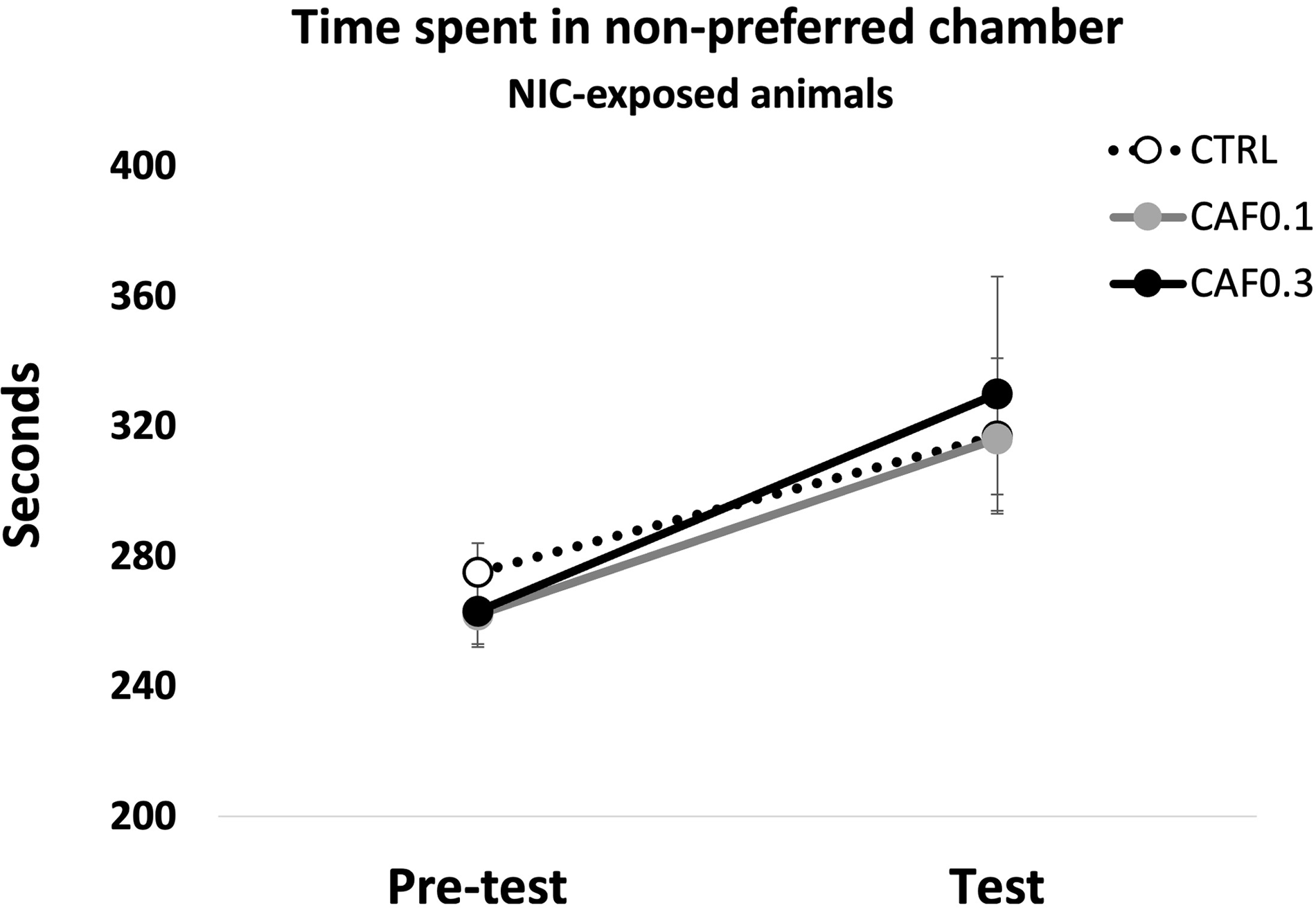
Fig. 3. Time spent in the non-preferred chamber in the pre-test and test sessions of the Conditioned Place Preference test (CPP). Mice were chronically exposed to caffeine (CAF) in the drinking water at one of two concentrations (0.1 or 0.3 g/L), or to filtered water (CTRL) during development and, at the beginning of each conditioning session, were i.p. injected with nicotine (NIC: 0.5 mg/Kg) in the non-preferred chamber. CTRL, ♀ = 11 + ♂ = 10; CAF0.1, ♀ = 8 + ♂ = 9; CAF0.3, ♀ = 9 + ♂ = 13. Values are means ± S.E.M.
Discussion
Caffeine consumption occurs throughout life and can be particularly high during adolescence, in great part due to energy drink consumption. Reference Pennington, Johnson, Delaney and Blankenship6 Adolescence is also when nicotine use typically begins and, despite the gradual decrease in consumption of combustible tobacco products, ENDS use among adolescents had a fast increase in the last few years. Reference Cullen, Gentzke and Sawdey30–Reference Miech, Leventhal, Johnston, O’Malley, Patrick and Barrington-Trimis32 Despite the well-established epidemiological association between caffeine and nicotine, Reference Kendler, Myers and Prescott67,Reference Martin, Cook and Woodring68 the underlying causes and neurobiological mechanisms are still poorly understood. Besides, most studies in animal models focus on a particular period of caffeine exposure (e.g. adolescence Reference Hinton, Andres-Beck and Nett13,Reference Ibrahim, Kamal, Tikamdas, Nouh, Tian and Sayed69–Reference Krahe, Filgueiras and Quaresma72 or prenatal life Reference Temple73,Reference James74 ), which is usually not paralleled by the pattern of exposure that occurs in humans. Here, we tested the hypothesis that lifetime caffeine exposure alters nicotine effects and increases its reward properties during adolescence. In our exposure model, caffeine did not affect nicotine-evoked psychostimulant effects or CPP. However, the higher dose of caffeine significantly decreased the time spent in the center of the OF and this effect was reversed by nicotine exposure.
The doses of caffeine used here, previously shown to antagonize adenosine receptors, Reference Silva, Métin and Fazeli10,Reference Ardais, Borges, Rocha, Sallaberry, Cunha and Porciúncula12,Reference Björklund, Kahlström, Salmi and Fredholm75–Reference El, Ledent and Ois77 failed to affect food and fluid intake as well as body mass both in dams and offspring, which corroborates previous findings. Reference Ardais, Borges, Rocha, Sallaberry, Cunha and Porciúncula12,Reference da Silva Gonçalves, Leal-Rocha, Manhães, Filgueiras, Abreu-Villaça and Ribeiro-Carvalho14 In contrast, the reduction in the time spent in the center of the OF indicates that lifetime exposure to the higher caffeine dose (0.3 g/L) increased anxiety-like behavior during adolescence. This result is in accordance with studies that evaluated the effects of caffeine on anxiety-like behaviors both during early development (prenatal/lactation) Reference Laureano-Melo, da Silveira and de Azevedo Cruz Seara9 and adolescence, Reference Neill, Newsom and Stafford78,Reference Bressan and Pilowsky79 and with previous evidence that human caffeine consumption is linked to negative consequences on mood, mostly anxiety disorders. However, when the time in center/locomotor activity ratio was used, the anxiogenic effect of caffeine failed to reach significance. This suggests that the use of other behavioral tests that assess anxiety-like behavior, such as the elevated plus maze, could be useful to corroborate our findings. Interestingly, nicotine reversed caffeine-induced effects on the time spent in the center of the OF. Even though nicotine exposure per se increased total activity, no significant effect was detected for the activity in the center of arena. OF may be used to model anxiety responses when someone is confronted with stressful or threatening situations. Reference Prut and Belzung49 In this sense, considering the strong association between anxiety and tobacco consumption, Reference McGranahan, Patzlaff, Grady, Heinemann and Booker36,Reference Morissette, Tull, Gulliver, Kamholz and Zimering54,Reference Picciotto, Brunzell and Caldarone55 our data raise the possibility that the increase in basal anxiety levels induced by the higher dose of caffeine could facilitate the use of nicotine by adolescents to mitigate this negative effect on mood. Besides, it could facilitate relapse to tobacco use. In accordance with this possibility, chronic caffeine was shown to increase anxiogenic-like effects associated with nicotine and alcohol withdrawal in rodents. Reference da Silva Gonçalves, Leal-Rocha, Manhães, Filgueiras, Abreu-Villaça and Ribeiro-Carvalho14,Reference Matovu and Alele80 Here, we failed to identify alterations in the expression of hippocampal 5-HT1A, which suggests that this receptor, in the hippocampus, does not play a critical role in the anxiety-like behavior evoked by lifetime caffeine. Arnold and colleagues Reference Arnold, Williams and McArthur71 showed that caffeine exposure during adolescence does not affect mRNA expression of htr1a, the serotonin 1a receptor gene, however, it downregulates mRNA expression of other proteins involved in serotonergic transmission in the dorsal raphe nucleus of rats. These results suggest that other receptors and brain regions relevant to serotonergic-mediated modulation of anxiety should be investigated.
Nicotine-induced hyperactivity and CPP are useful phenomena to study nicotine susceptibility, both behaviors being associated with nicotine effects on the mesocorticolimbic pathway. Reference Wilar, Shinoda, Sasaoka and Fukunaga81,Reference Cohen, Mackey and Grady82 Here, 0.5 mg/Kg of nicotine increased locomotor activity and time spent in the non-preferred chamber after conditioning during adolescence, which corroborates previous studies from our group. Reference Dutra-Tavares, Silva and Nunes-Freitas59,Reference Nunes-Freitas, Manhães and Dutra-Tavares60,Reference Dutra-Tavares, Manhães and Silva83
Regarding the CPP, the increased time spent in the non-preferred chamber indicates nicotine-evoked conditioning. However, the use of a biased CPP design makes it difficult to identify the participation of nicotine anxiolytic-like effects on its place preference conditioning, since a decrease in anxiety levels could increase the time spent in the nonpreferred chamber by reducing the aversion for this chamber. In this sense, albeit nicotine mitigated anxiety-like effects induced by the higher dose of caffeine, nicotine failed to reduce anxiety levels in the OF per se. Besides, nicotine-induced hyperactivity was evident in this test. Altogether these data support the idea that our result in the CPP is not mainly caused by a reduced aversion associated with the anxiolytic-like effects of this drug instead of the CPP produced by its rewarding effects.
Caffeine is a nonselective A1 and A2A receptors antagonist, which can form heteromeric complexes with dopaminergic receptors. Reference Taura, Sahlholm, Craenenbroeck and Ciruela84,Reference Franco, Lluis and Canela85 Both A2A and D2 receptors in the striatum are involved in the psychostimulant effects of caffeine, modulating dopaminergic transmission and influencing the reward properties of other drugs. Reference Ferré86,Reference Filip, Zaniewska, Frankowska, Wydra and Fuxe87 Besides the fact that caffeine and nicotine act as dopaminergic modulators, there is evidence of interactions between adenosine and cholinergic systems in the control of dopamine release in the rat striatum. Reference Metaxas, Al-hasani and Farshim88,Reference Garção, Szabó, Wopereis, Castro and Tomé89 Despite that, here, lifetime caffeine did not affect nicotine locomotor and reward responses in adolescent mice. In addition, no differences in dopamine, DOPAC and dopamine turnover were found between experimental groups in the frontal cerebral cortex.
It is important to mention that even though the choice of the nicotine dose was based on its conditioning and hyperactivity effects, by using a single dose we cannot rule out the possibility that the absence of lifetime caffeine exposure interaction with nicotine during adolescence was due to a ceiling effect of the latter. Future studies with multiple nicotine doses, particularly lower doses, will be useful to better understand caffeine interactions with nicotine reward properties. Additionally, the development of caffeine tolerance could be a key factor for the lack of caffeine-nicotine interactions in the present study. Accordingly, while low to moderate doses of caffeine are known to produce hyperactivity, Reference Porciúncula, Sallaberry, Mioranzza, Henrique, Botton and Rosemberg90 here, lifetime caffeine per se did not affect locomotor activity. Chronic exposure to caffeine seems to cause tolerance to its motor-activating effects, possibly by alterations on A1 receptor function. Reference Karcz-kubicha, Antoniou and Terasmaa91 Quarta and colleagues Reference Quarta, Ferre, Solinas, bing and Goldberg92 showed that chronic caffeine exposure completely blocks the effects of caffeine on dopamine release in the rat’s nucleus accumbens, which could be one of the factors that explain the absence of summation effects with nicotine in the present study.
Extrapolation of these data suggests that moderate caffeine consumption throughout life, including gestation and lactation, may not predispose nicotine use in adolescence by increasing its reward response. On the other hand, there is evidence pointing to an increase in the psychoactive effects of drugs, such as nicotine and ethanol, due to coexposure to high acute doses of caffeine. Reference Liu and Jernigan23,Reference Krahe, Filgueiras and Quaresma72,Reference Rezvani, Sexton, Johnson, Wells, Gordon and Levin93,Reference Porru, López-Cruz, Carratalá-Ros, Salamone, Acquas and Correa94 This suggests that caffeine consumption at high doses by in susceptible individuals could pose a risk for problematic use of other substances.
Albeit lifetime caffeine failed to affect the reward response to nicotine during adolescence, the higher dose used here induced anxiety-like behavior; an effect reversed by acute nicotine administration. The neurobiological mechanisms that underlie such phenomenon remain unclear, so, future studies are needed to systematically investigate this issue. Nevertheless, the caffeine-induced anxiety-like effect advises limiting consumption of this substance during development, including adolescence. In addition, considering that adolescence is a period of increased susceptibility to drugs of abuse, mood alterations elicited by caffeine could be a risk factor to nicotine consumption. Despite being considered a safe substance to consume, the association between caffeine, anxiety, and other drugs is a matter of great concern.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/S2040174423000077.
Acknowledgements
The authors are thankful to Ulisses Risso for animal care.
Financial support
This work was supported by fellowships from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES-BRAZIL) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq-BRAZIL) and grants from Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ-BRAZIL) (A. R-C., E-26/202.764/2015) (Y. A-V., E-26/103.191/2011 and E-26/111.160/2011) and CNPq-BRAZIL (Y. A-V., 306173/2012-3).
Conflicts of interest
None.
Ethical standards
The authors assert that all procedures contributing to this work comply with the ethical standards of the Guide for the Care and Use of Laboratory Animals as adopted and promulgated by the National Institutes of Health and of the relevant national guides on the care and use of laboratory animals (Brazilian Law #11.794/2008) and has been approved by the institutional committee (protocol #: CEUA/038/2014).






